
Longitudinal Dynamics of SARS-CoV-2 IgG Antibody Responses after the Two-Dose Regimen of BNT162b2 Vaccination and the Effect of a Third Dose on Healthcare Workers in Japan

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
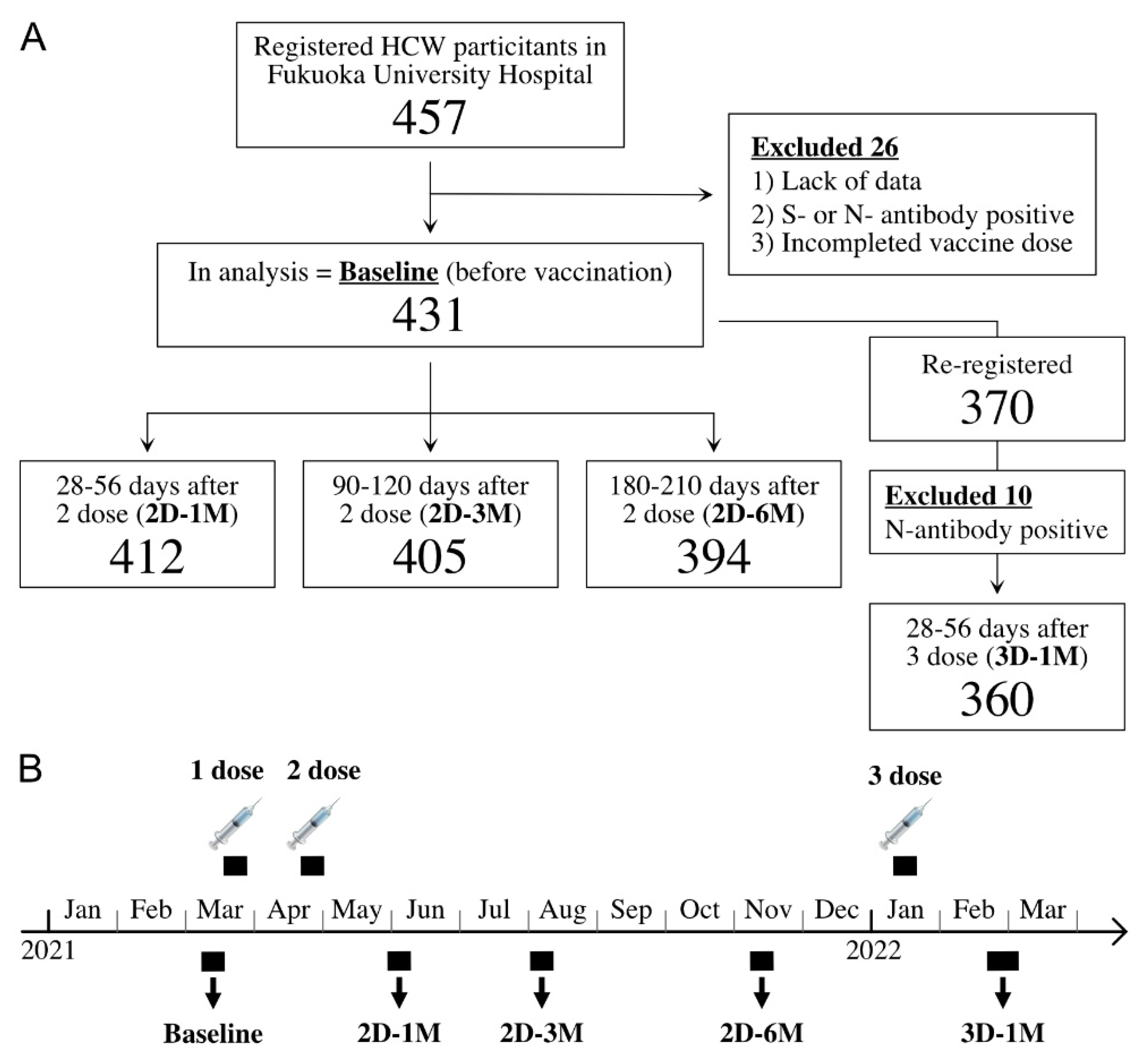
2.1. Study Population and Survey Period
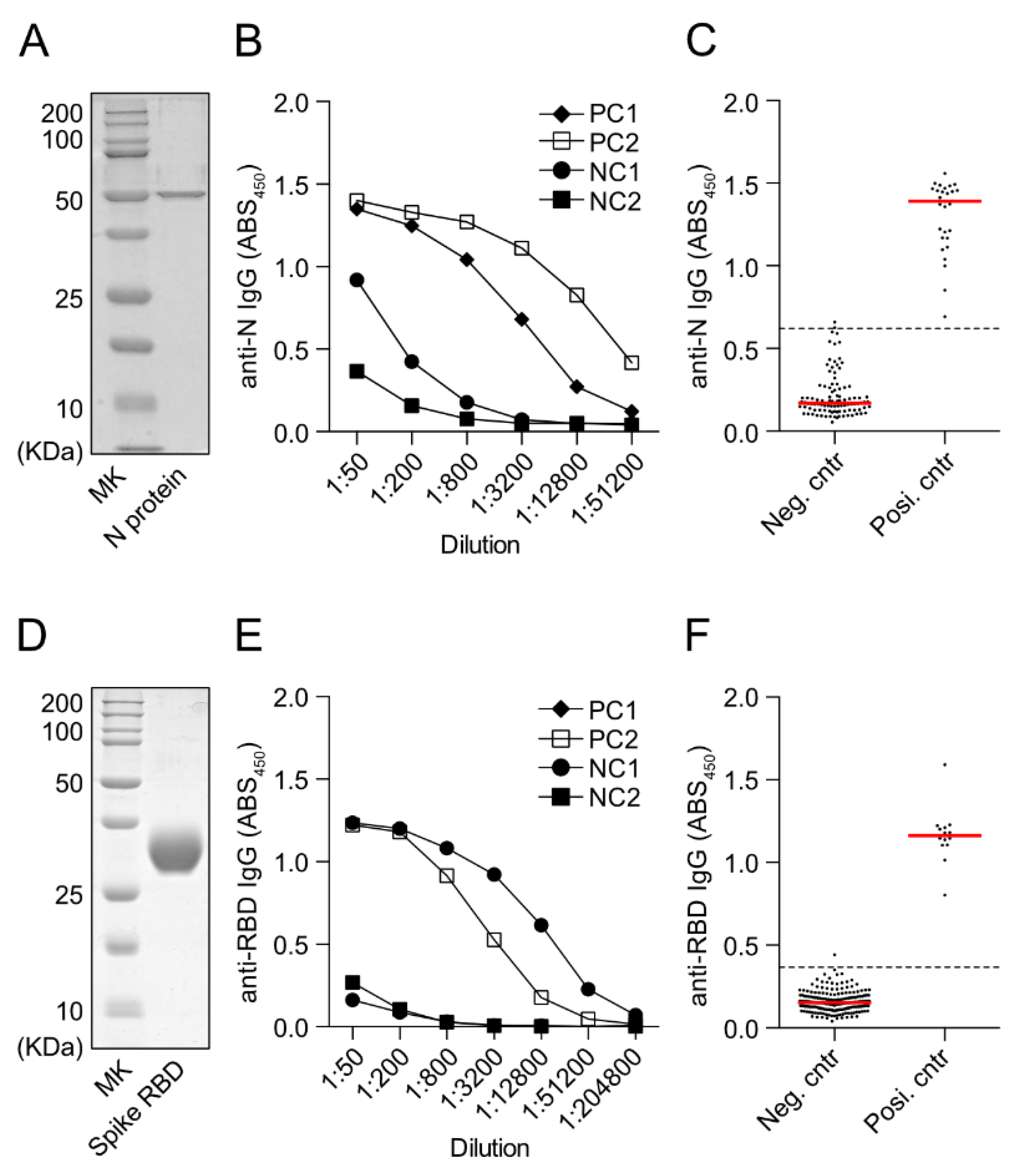
2.2. ELISA for Anti-SARS-CoV-2 RBD IgG or Anti-N-Protein IgG Detection
2.3. Statistical Analyses
3. Results
3.1. Demographic Data
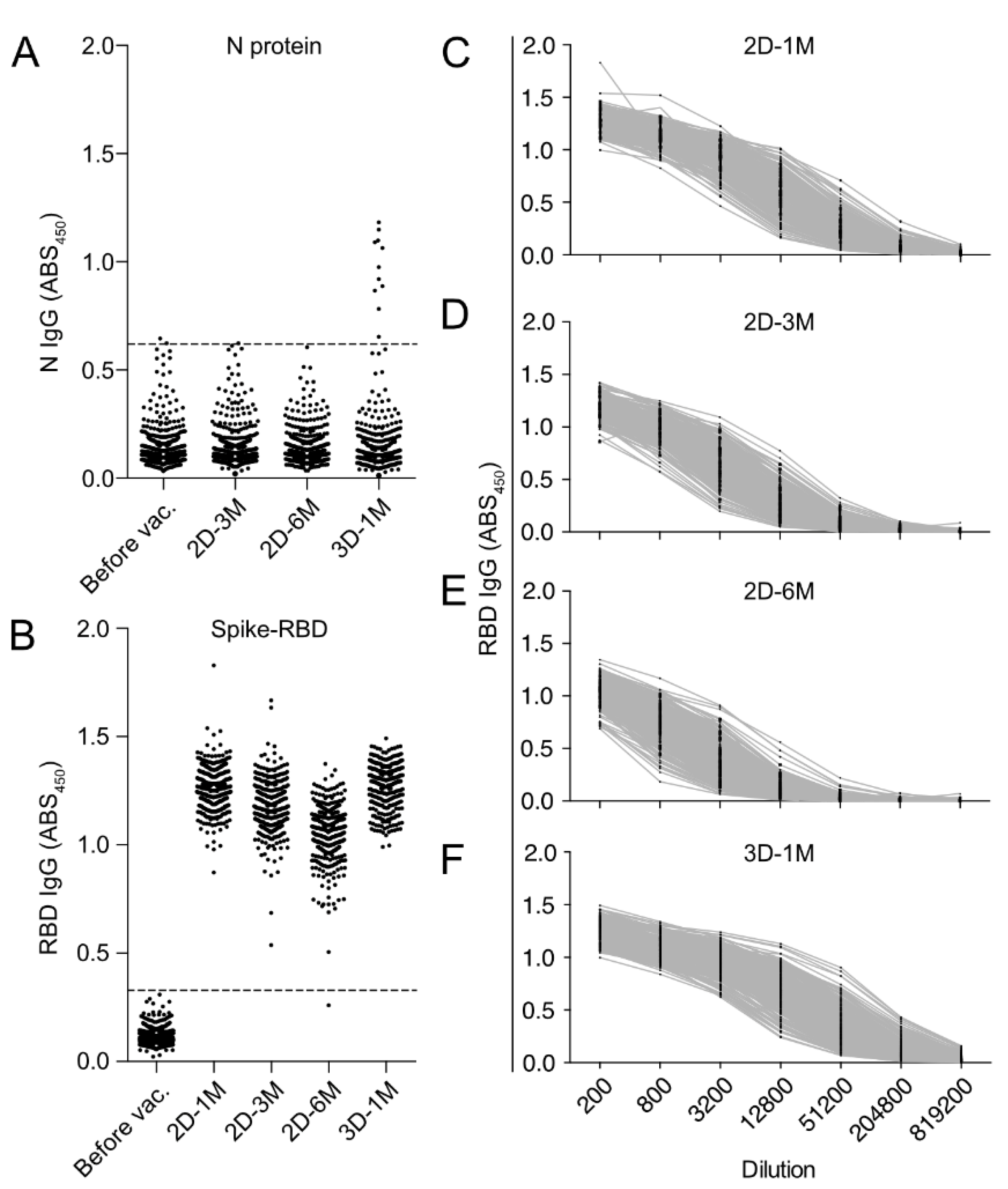
3.2. Measurement of Anti-SARS-CoV-2 N-Protein IgG and Anti-SARS-CoV-2 S-Protein RBD IgG by In-House ELISA
3.3. Kinetics of Anti-RBD IgG Titers after the Second Dose and the Third Dose of BNT162b2 Vaccination in HCWs in Japan
3.4. Longitudinal Kinetics of Anti-SARS-CoV-2 RBD IgG Antibodies after the Second and Third Doses of BNT162b2 Vaccine
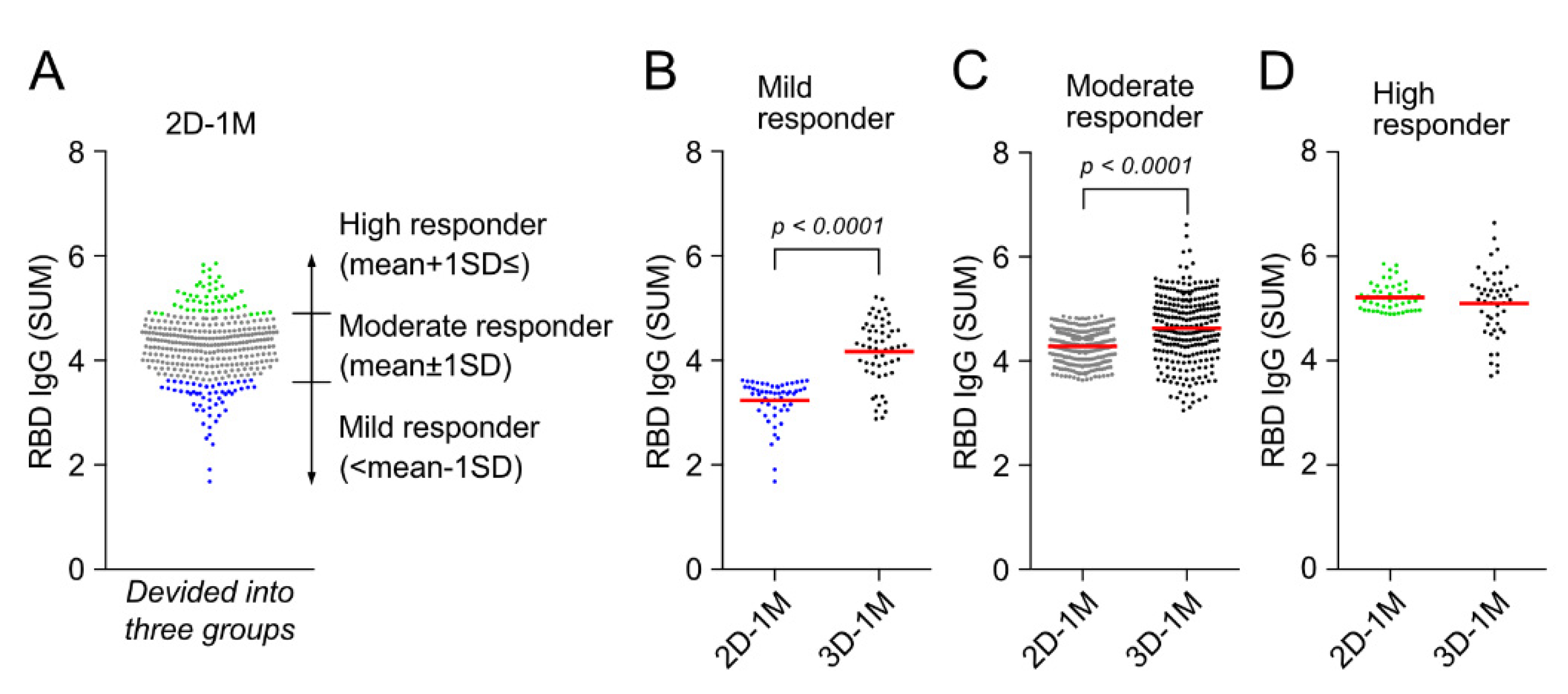
3.5. Third Dose of BNT162b2 Vaccination Augments Anti-RBD IgG Responses of Mild Responder at 1 M after the Second Dose of BNT162b2 Vaccination
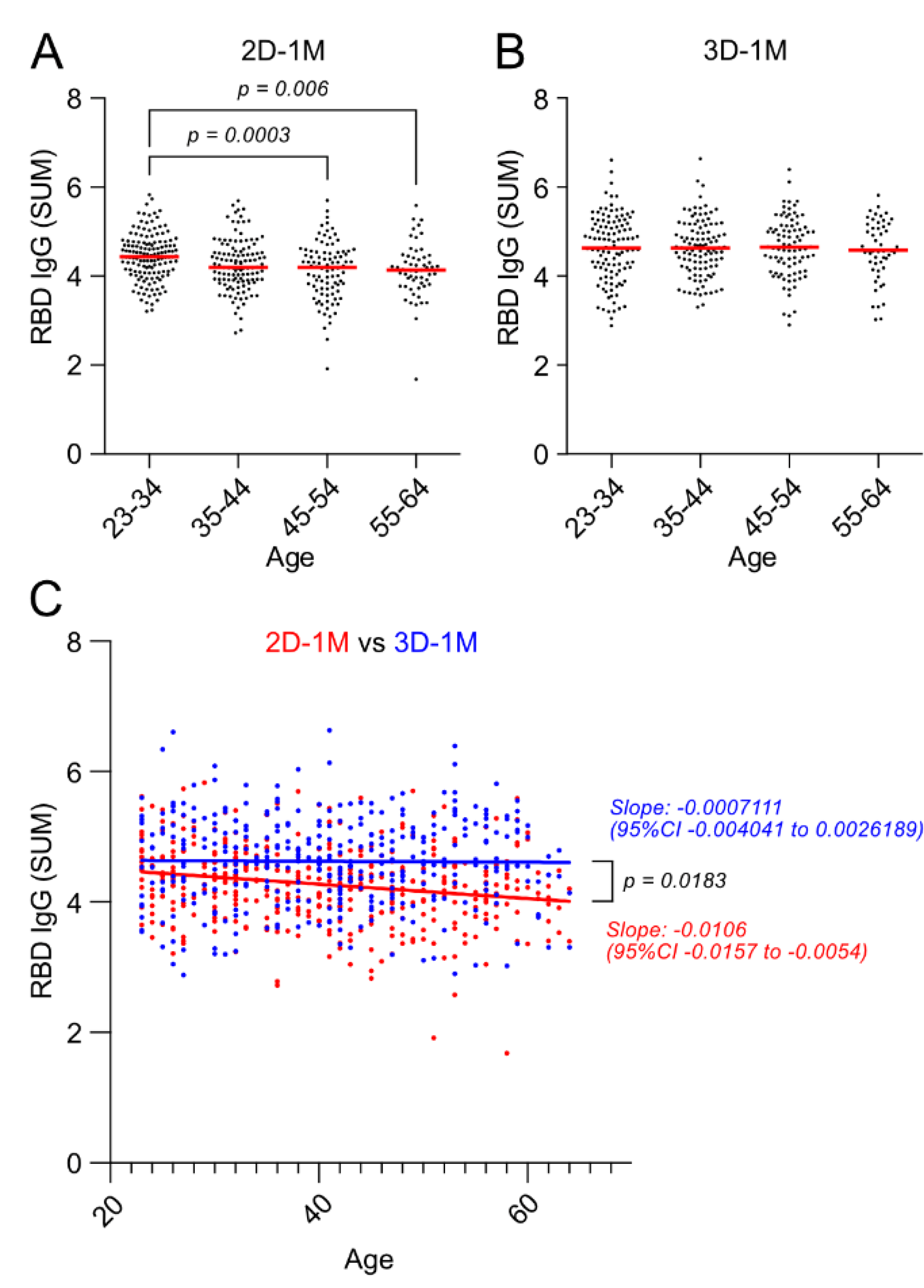
3.6. Negative Effect of Age on Anti-RBD IgG Responses after the Second Dose of BNT162b2 Vaccination Was Lost after the Third Dose of BNT162b2 Vaccination
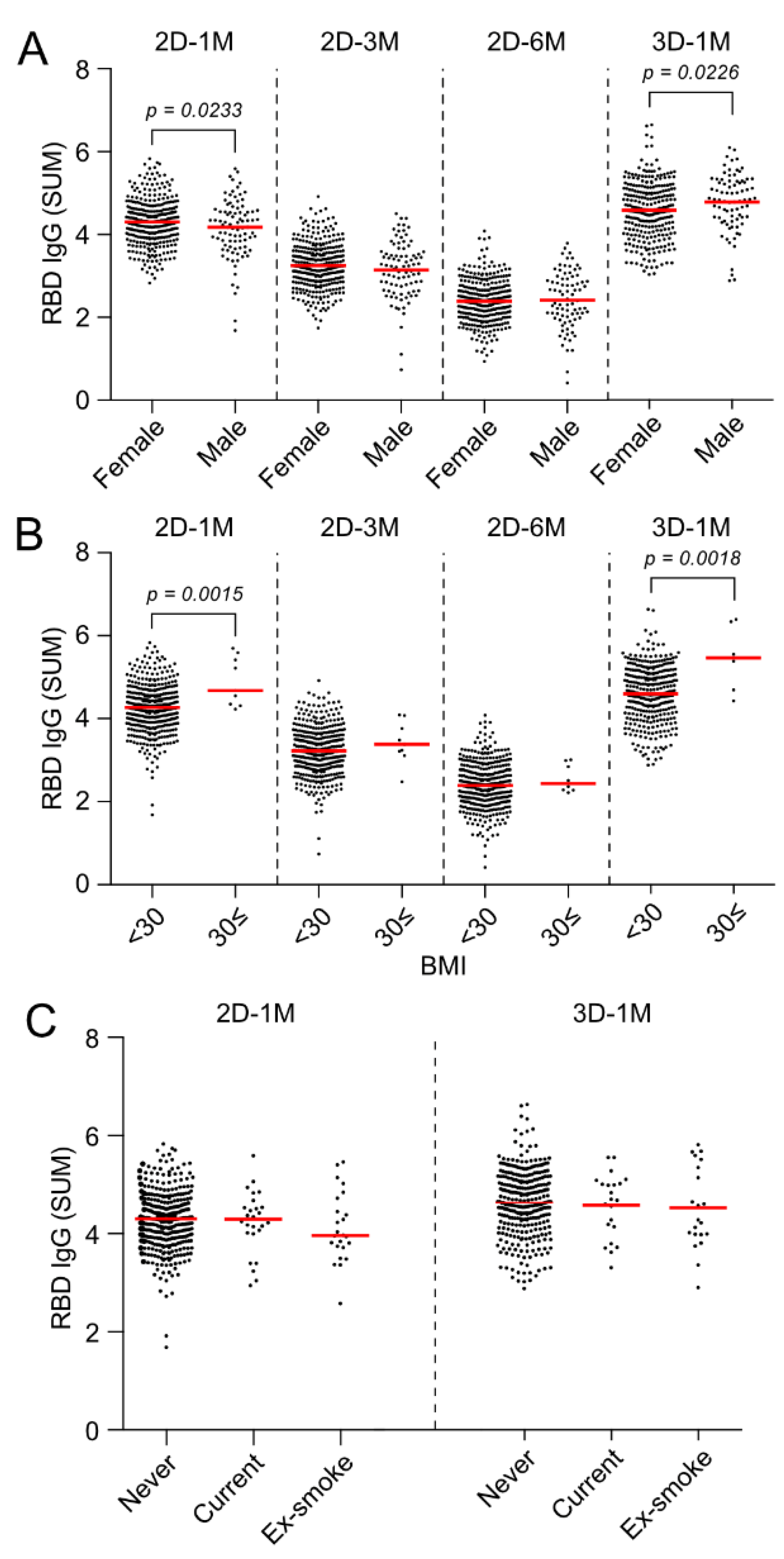
3.7. Effect of Gender and BMI on Anti-RBD IgG Responses at 3 M and 6 M after the Second Dose and 1 M after the Third Dose of the BNT162b2 Vaccination
3.8. Multivariate Analysis on the Effects of Sex, Age and BMI on Anti-RBD IgG Responses at 1 M after the Second Dose and 1 M after the Third Dose of BNT162b2 Vaccinations
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Polack, F.P.; Thomas, S.J.; Kitchin, N.; Absalon, J.; Gurtman, A.; Lockhart, S.; Perez, J.L.; Pérez Marc, G.; Moreira, E.D.; Zerbini, C.; et al. Safety and Efficacy of the BNT162b2 mRNA Covid-19 Vaccine. N. Engl. J. Med. 2020, 383, 2603–2615. [Google Scholar] [CrossRef] [PubMed]
- Dagan, N.; Barda, N.; Kepten, E.; Miron, O.; Perchik, S.; Katz, M.A.; Hernán, M.A.; Lipsitch, M.; Reis, B.; Balicer, R.D. BNT162b2 mRNA Covid-19 Vaccine in a Nationwide Mass Vaccination Setting. N. Engl. J. Med. 2021, 384, 1412–1423. [Google Scholar] [CrossRef] [PubMed]
- Hall, V.J.; Foulkes, S.; Saei, A.; Andrews, N.; Oguti, B.; Charlett, A.; Wellington, E.; Stowe, J.; Gillson, N.; Atti, A.; et al. COVID-19 vaccine coverage in health-care workers in England and effectiveness of BNT162b2 mRNA vaccine against infection (SIREN): A prospective, multicentre, cohort study. Lancet 2021, 397, 1725–1735. [Google Scholar] [CrossRef]
- El Sahly, H.M.; Baden, L.R.; Essink, B.; Doblecki-Lewis, S.; Martin, J.M.; Anderson, E.J.; Campbell, T.B.; Clark, J.; Jackson, L.A.; Fichtenbaum, C.J.; et al. Efficacy of the mRNA-1273 SARS-CoV-2 Vaccine at Completion of Blinded Phase. N. Engl. J. Med. 2021, 385, 1774–1785. [Google Scholar] [CrossRef] [PubMed]
- Baden, L.R.; El Sahly, H.M.; Essink, B.; Kotloff, K.; Frey, S.; Novak, R.; Diemert, D.; Spector, S.A.; Rouphael, N.; Creech, C.B.; et al. Efficacy and Safety of the mRNA-1273 SARS-CoV-2 Vaccine. N. Engl. J. Med. 2021, 384, 403–416. [Google Scholar] [CrossRef]
- Planas, D.; Veyer, D.; Baidaliuk, A.; Staropoli, I.; Guivel-Benhassine, F.; Rajah, M.M.; Planchais, C.; Porrot, F.; Robillard, N.; Puech, J.; et al. Reduced sensitivity of SARS-CoV-2 variant Delta to antibody neutralization. Nature 2021, 596, 276–280. [Google Scholar] [CrossRef]
- Elliott, P.; Haw, D.; Wang, H.; Eales, O.; Walters, C.E.; Ainslie, K.E.C.; Atchison, C.; Fronterre, C.; Diggle, P.J.; Page, A.J.; et al. Exponential growth, high prevalence of SARS-CoV-2, and vaccine effectiveness associated with the Delta variant. Science 2021, 374, eabl9551. [Google Scholar] [CrossRef]
- Hatzakis, A.; Karabinis, A.; Roussos, S.; Pantazis, N.; Degiannis, D.; Chaidaroglou, A.; Petsios, K.; Pavlopoulou, I.; Tsiodras, S.; Paraskevis, D.; et al. Modelling SARS-CoV-2 Binding Antibody Waning 8 Months after BNT162b2 Vaccination. Vaccines 2022, 10, 285. [Google Scholar] [CrossRef]
- Levin, E.G.; Lustig, Y.; Cohen, C.; Fluss, R.; Indenbaum, V.; Amit, S.; Doolman, R.; Asraf, K.; Mendelson, E.; Ziv, A.; et al. Waning Immune Humoral Response to BNT162b2 Covid-19 Vaccine over 6 Months. N. Engl. J. Med. 2021, 385, e84. [Google Scholar] [CrossRef]
- Israel, A.; Shenhar, Y.; Green, I.; Merzon, E.; Golan-Cohen, A.; Schäffer, A.A.; Ruppin, E.; Vinker, S.; Magen, E. Large-Scale Study of Antibody Titer Decay following BNT162b2 mRNA Vaccine or SARS-CoV-2 Infection. Vaccines 2021, 10, 64. [Google Scholar] [CrossRef]
- Doria-Rose, N.; Suthar, M.S.; Makowski, M.; O’Connell, S.; McDermott, A.B.; Flach, B.; Ledgerwood, J.E.; Mascola, J.R.; Graham, B.S.; Lin, B.C.; et al. Antibody Persistence through 6 Months after the Second Dose of mRNA-1273 Vaccine for Covid-19. N. Engl. J. Med. 2021, 384, 2259–2261. [Google Scholar] [CrossRef] [PubMed]
- Campo, F.; Venuti, A.; Pimpinelli, F.; Abril, E.; Blandino, G.; Conti, L.; De Virgilio, A.; De Marco, F.; Di Noia, V.; Di Domenico, E.G.; et al. Antibody Persistence 6 Months Post-Vaccination with BNT162b2 among Health Care Workers. Vaccines 2021, 9, 1125. [Google Scholar] [CrossRef] [PubMed]
- Ferrari, D.; Clementi, N.; Criscuolo, E.; Ambrosi, A.; Corea, F.; Di Resta, C.; Tomaiuolo, R.; Mancini, N.; Locatelli, M.; Plebani, M.; et al. Antibody Titer Kinetics and SARS-CoV-2 Infections Six Months after Administration with the BNT162b2 Vaccine. Vaccines 2021, 9, 1357. [Google Scholar] [CrossRef] [PubMed]
- Stadlbauer, D.; Amanat, F.; Chromikova, V.; Jiang, K.; Strohmeier, S.; Arunkumar, G.A.; Tan, J.; Bhavsar, D.; Capuano, C.; Kirkpatrick, E.; et al. SARS-CoV-2 Seroconversion in Humans: A Detailed Protocol for a Serological Assay, Antigen Production, and Test Setup. Curr. Protoc. Microbiol. 2020, 57, e100. [Google Scholar] [CrossRef] [Green Version]
- Amanat, F.; Stadlbauer, D.; Strohmeier, S.; Nguyen, T.H.O.; Chromikova, V.; McMahon, M.; Jiang, K.; Arunkumar, G.A.; Jurczyszak, D.; Polanco, J.; et al. A serological assay to detect SARS-CoV-2 seroconversion in humans. Nat. Med. 2020, 26, 1033–1036. [Google Scholar] [CrossRef]
- Weisberg, S.P.; Connors, T.J.; Zhu, Y.; Baldwin, M.R.; Lin, W.H.; Wontakal, S.; Szabo, P.A.; Wells, S.B.; Dogra, P.; Gray, J.; et al. Distinct antibody responses to SARS-CoV-2 in children and adults across the COVID-19 clinical spectrum. Nat. Immunol. 2021, 22, 25–31. [Google Scholar] [CrossRef]
- Lustig, Y.; Sapir, E.; Regev-Yochay, G.; Cohen, C.; Fluss, R.; Olmer, L.; Indenbaum, V.; Mandelboim, M.; Doolman, R.; Amit, S.; et al. BNT162b2 COVID-19 vaccine and correlates of humoral immune responses and dynamics: A prospective, single-centre, longitudinal cohort study in health-care workers. Lancet Respir. Med. 2021, 9, 999–1009. [Google Scholar] [CrossRef]
- Terpos, E.; Trougakos, I.P.; Apostolakou, F.; Charitaki, I.; Sklirou, A.D.; Mavrianou, N.; Papanagnou, E.D.; Liacos, C.I.; Gumeni, S.; Rentziou, G.; et al. Age-dependent and gender-dependent antibody responses against SARS-CoV-2 in health workers and octogenarians after vaccination with the BNT162b2 mRNA vaccine. Am. J. Hematol. 2021, 96, E257–E259. [Google Scholar] [CrossRef]
- Kageyama, T.; Ikeda, K.; Tanaka, S.; Taniguchi, T.; Igari, H.; Onouchi, Y.; Kaneda, A.; Matsushita, K.; Hanaoka, H.; Nakada, T.A.; et al. Antibody responses to BNT162b2 mRNA COVID-19 vaccine and their predictors among healthcare workers in a tertiary referral hospital in Japan. Clin. Microbiol. Infect. 2021, 27, 1861.e1–1861.e5. [Google Scholar] [CrossRef]
- Pellini, R.; Venuti, A.; Pimpinelli, F.; Abril, E.; Blandino, G.; Campo, F.; Conti, L.; De Virgilio, A.; De Marco, F.; Di Domenico, E.G.; et al. Initial observations on age, gender, BMI and hypertension in antibody responses to SARS-CoV-2 BNT162b2 vaccine. EClinicalMedicine 2021, 36, 100928. [Google Scholar] [CrossRef]
- Eliakim-Raz, N.; Leibovici-Weisman, Y.; Stemmer, A.; Ness, A.; Awwad, M.; Ghantous, N.; Stemmer, S.M. Antibody Titers Before and After a Third Dose of the SARS-CoV-2 BNT162b2 Vaccine in Adults Aged ≥60 Years. JAMA 2021, 326, 2203–2204. [Google Scholar] [CrossRef] [PubMed]
- Ward, H.; Whitaker, M.; Flower, B.; Tang, S.N.; Atchison, C.; Darzi, A.; Donnelly, C.A.; Cann, A.; Diggle, P.J.; Ashby, D.; et al. Population antibody responses following COVID-19 vaccination in 212,102 individuals. Nat. Commun. 2022, 13, 907. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.H.; Kim, Y.R.; Heo, S.T.; Oh, H.; Kim, M.; Lee, H.R.; Yoo, J.R. Healthcare Workers in South Korea Maintain a SARS-CoV-2 Antibody Response Six Months After Receiving a Second Dose of the BNT162b2 mRNA Vaccine. Front. Immunol. 2022, 13, 827306. [Google Scholar] [CrossRef] [PubMed]
- McIntyre, P.B.; Aggarwal, R.; Jani, I.; Jawad, J.; Kochhar, S.; MacDonald, N.; Madhi, S.A.; Mohsni, E.; Mulholland, K.; Neuzil, K.M.; et al. COVID-19 vaccine strategies must focus on severe disease and global equity. Lancet 2022, 399, 406–410. [Google Scholar] [CrossRef]
- Cho, A.; Muecksch, F.; Schaefer-Babajew, D.; Wang, Z.; Finkin, S.; Gaebler, C.; Ramos, V.; Cipolla, M.; Mendoza, P.; Agudelo, M.; et al. Anti-SARS-CoV-2 receptor-binding domain antibody evolution after mRNA vaccination. Nature 2021, 600, 517–522. [Google Scholar] [CrossRef]
- Sahin, U.; Muik, A.; Derhovanessian, E.; Vogler, I.; Kranz, L.M.; Vormehr, M.; Baum, A.; Pascal, K.; Quandt, J.; Maurus, D.; et al. COVID-19 vaccine BNT162b1 elicits human antibody and TH1 T cell responses. Nature 2020, 586, 594–599. [Google Scholar] [CrossRef]
- Guerrera, G.; Picozza, M.; D’Orso, S.; Placido, R.; Pirronello, M.; Verdiani, A.; Termine, A.; Fabrizio, C.; Giannessi, F.; Sambucci, M.; et al. BNT162b2 vaccination induces durable SARS-CoV-2-specific T cells with a stem cell memory phenotype. Sci. Immunol. 2021, 6, eabl5344. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sex | BMI | ||||
|---|---|---|---|---|---|
| Female | 337 | (78.2%) | <18.5 | 62 | (14.4%) |
| Male | 94 | (21.8%) | 18.5≤, <25 | 313 | (72.6%) |
| Age | 25≤, <30 | 43 | (10.0%) | ||
| 30≤ | 9 | (2.1%) | |||
| 23–34 | 150 | (34.8%) | NA | 4 | (0.9%) |
| 35–44 | 126 | (29.2%) | Smoking status | ||
| 45–54 | 95 | (22.0%) | Never smoker | 378 | (87.7%) |
| 55–64 | 58 | (13.5%) | Current smoker | 28 | (6.5%) |
| NA | 2 | (0.5%) | Ex-smoker | 25 | (5.8%) |
| Mild Responder | Moderate Responder | High Responder | ||||
|---|---|---|---|---|---|---|
| n | 53 | 249 | 50 | |||
| Sex | ||||||
| Female | 33 | (62.3%) | 203 | (81.5%) | 39 | (78.0%) |
| Male | 20 | (37.7%) | 46 | (18.5%) | 11 | (22.0%) |
| Age | ||||||
| median | 45.5 | 41 | 40 | |||
| IQR | 36.0–52.0 | 32.0–41.0 | 28.3–40.0 | |||
| BMI | ||||||
| median | 21.5 | 20.7 | 21.8 | |||
| IQR | 20.0–22.7 | 19.1–23.0 | 19.9–24.1 | |||
| Smoking | ||||||
| never | 43 | (81.1%) | 224 | (90.0%) | 45 | (90.0%) |
| current | 5 | (9.4%) | 13 | (5.2%) | 2 | (4.0%) |
| ex- | 5 | (9.4%) | 12 | (4.8%) | 3 | (6.0%) |
| 2D-1M | 3D-1M | ||
|---|---|---|---|
| All | All | Selected 1 | |
| (Intercept) | 4.380 *** | 4.037 *** | 3.958 *** |
| p < 0.001 | p < 0.001 | p < 0.001 | |
| Age | −0.013 (−0.230) *** | −0.003 (−0.044) | |
| p < 0.001 | p = 0.412 | ||
| Sex | −0.230 (−0.157) ** | 0.157 (0.096) + | 0.158 (0.096) + |
| p = 0.004 | p = 0.083 | p = 0.080 | |
| BMI | 0.022 (0.106) + | 0.031 (0.134) * | 0.029 (0.127) * |
| p = 0.054 | p = 0.017 | p = 0.022 | |
| Num. Obs. | 346 | 346 | 346 |
| R2 | 0.074 | 0.034 | 0.032 |
| R2 Adj. | 0.066 | 0.025 | 0.026 |
| AIC | 621.1 | 716.0 | 714.7 |
| RMSE | 0.59 | 0.68 | 0.67 |
| F | 9.166 | 3.967 | 5.619 |
| p | <0.001 | 0.008 | 0.004 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sakamoto, A.; Yoshimura, M.; Itoh, R.; Ozuru, R.; Ishii, K.; Sechi, Y.; Nabeshima, S.; Hiromatsu, K. Longitudinal Dynamics of SARS-CoV-2 IgG Antibody Responses after the Two-Dose Regimen of BNT162b2 Vaccination and the Effect of a Third Dose on Healthcare Workers in Japan. Vaccines 2022, 10, 830. https://doi.org/10.3390/vaccines10060830
Sakamoto A, Yoshimura M, Itoh R, Ozuru R, Ishii K, Sechi Y, Nabeshima S, Hiromatsu K. Longitudinal Dynamics of SARS-CoV-2 IgG Antibody Responses after the Two-Dose Regimen of BNT162b2 Vaccination and the Effect of a Third Dose on Healthcare Workers in Japan. Vaccines. 2022; 10(6):830. https://doi.org/10.3390/vaccines10060830
Chicago/Turabian StyleSakamoto, Atsuhiko, Michinobu Yoshimura, Ryota Itoh, Ryo Ozuru, Kazunari Ishii, Yusuke Sechi, Shigeki Nabeshima, and Kenji Hiromatsu. 2022. "Longitudinal Dynamics of SARS-CoV-2 IgG Antibody Responses after the Two-Dose Regimen of BNT162b2 Vaccination and the Effect of a Third Dose on Healthcare Workers in Japan" Vaccines 10, no. 6: 830. https://doi.org/10.3390/vaccines10060830
APA StyleSakamoto, A., Yoshimura, M., Itoh, R., Ozuru, R., Ishii, K., Sechi, Y., Nabeshima, S., & Hiromatsu, K. (2022). Longitudinal Dynamics of SARS-CoV-2 IgG Antibody Responses after the Two-Dose Regimen of BNT162b2 Vaccination and the Effect of a Third Dose on Healthcare Workers in Japan. Vaccines, 10(6), 830. https://doi.org/10.3390/vaccines10060830