
Variant Allele of ALDH2, rs671, Associates with Attenuated Post-Vaccination Response in Anti-SARS-CoV-2 Spike Protein IgG: A Prospective Study in the Japanese General Population

 , , ,
, , ,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Design and Participants
2.2. Serological Tests
2.3. Self-Administered Questionnaire
2.3.1. Covariates
2.3.2. Sensitivity Analysis
2.4. Genotyping
2.5. Statistical Analyses
3. Results
3.1. Baseline Characteristics
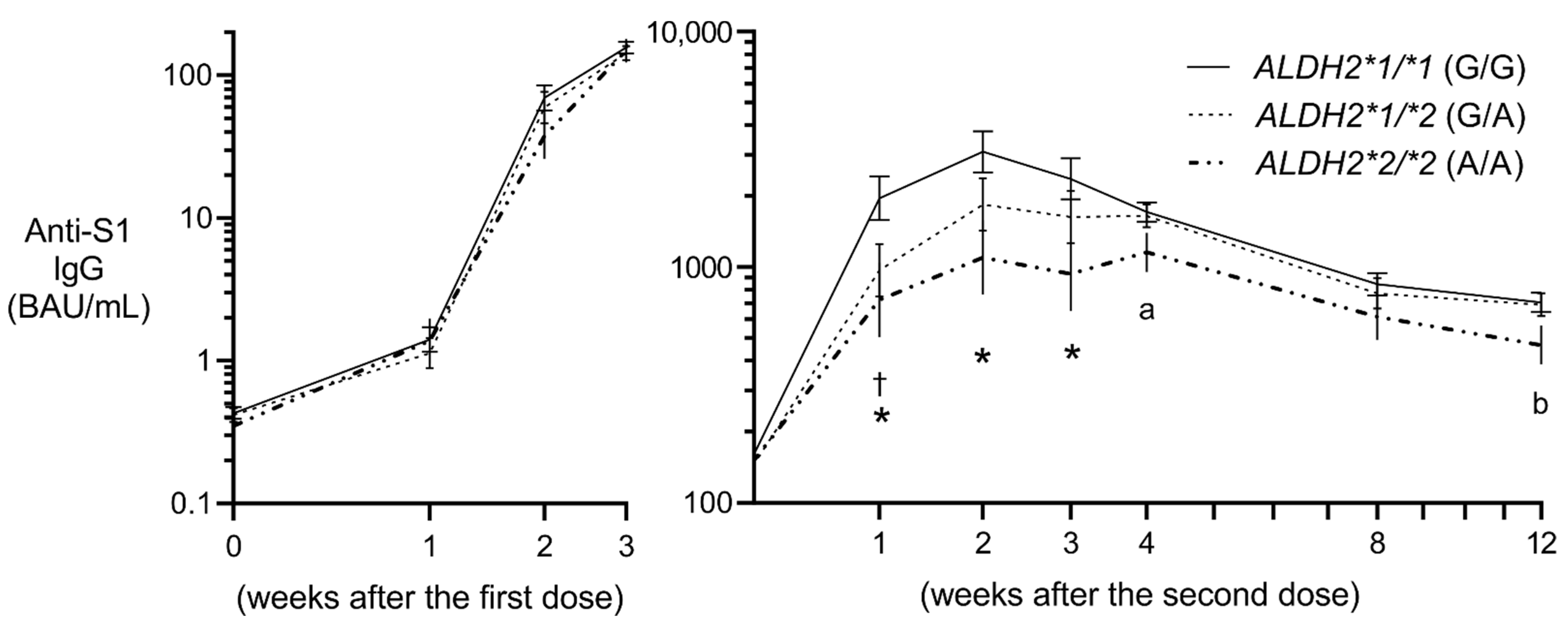
3.2. Antibody Production Post Vaccination
3.3. Effect of rs671 on Anti-S1 IgG Post Vaccination
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Marchitti, S.A.; Brocker, C.; Stagos, D.; Vasiliou, V. Non-P450 aldehyde oxidizing enzymes: The aldehyde dehydrogenase superfamily. Expert Opin. Drug Metab. Toxicol. 2008, 4, 697–720. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsumoto, A. The Bidirectional Effect of Defective ALDH2 Polymorphism and Disease Prevention. Adv. Exp. Med. Biol. 2019, 1193, 69–87. [Google Scholar] [PubMed]
- Sakaue, S.; Akiyama, M.; Hirata, M.; Matsuda, K.; Murakami, Y.; Kubo, M.; Kamatani, Y.; Okada, Y. Functional variants in ADH1B and ALDH2 are non-additively associated with all-cause mortality in Japanese population. Eur. J. Hum. Genet. 2020, 28, 378–382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shang, D.; Wang, P.; Tang, W.; Mo, R.; Lai, R.; Lu, J.; Li, Z.; Wang, X.; Cai, W.; Wang, H.; et al. Genetic Variations of ALDH (rs671) Are Associated with the Persistence of HBV Infection among the Chinese Han Population. Front. Med. 2022, 9, 811639. [Google Scholar] [CrossRef] [PubMed]
- Liao, W.L.; Chan, F.C.; Chang, K.P.; Chang, Y.W.; Chen, C.H.; Su, W.H.; Chang, H.H. Associations between ALDH Genetic Variants, Alcohol Consumption, and the Risk of Nasopharyngeal Carcinoma in an East Asian Population. Genes 2021, 12, 1547. [Google Scholar] [CrossRef]
- Matsumoto, A.; Thompson, D.C.; Chen, Y.; Kitagawa, K.; Vasiliou, V. Roles of defective ALDH2 polymorphism on liver protection and cancer development. Environ. Health Prev. Med. 2016, 21, 395–402. [Google Scholar] [CrossRef]
- Matsumoto, A. Fundamental properties of aldehyde dehydrogenase 2 (ALDH2) and the importance of the ALDH2 polymorphism. Nihon Eiseigaku Zasshi 2016, 71, 55–68. [Google Scholar] [CrossRef] [Green Version]
- Matsumoto, A. Importance of an aldehyde dehydrogenase 2 polymorphism in preventive medicine. Nihon Eiseigaku Zasshi 2018, 73, 9–20. [Google Scholar] [CrossRef] [Green Version]
- Sakaue, S.; Kanai, M.; Tanigawa, Y.; Karjalainen, J.; Kurki, M.; Koshiba, S.; Narita, A.; Konuma, T.; Yamamoto, K.; Akiyama, M.; et al. A cross-population atlas of genetic associations for 220 human phenotypes. Nat. Genet. 2021, 53, 1415–1424. [Google Scholar] [CrossRef]
- Matsumoto, A.; Ito, S.; Wakamatsu, K.; Ichiba, M.; Vasiliou, V.; Akao, C.; Song, B.J.; Fujita, M. Ethanol induces skin hyperpigmentation in mice with aldehyde dehydrogenase 2 deficiency. Chem. Biol. Interact 2019, 302, 61–66. [Google Scholar] [CrossRef]
- Matsumoto, A.; Nakashima, C.; Kimura, S.; Sueoka, E.; Aragane, N. ALDH2 polymorphism rs671 is a predictor of PD-1/PD-L1 inhibitor efficacy against thoracic malignancies. BMC Cancer 2021, 21, 584. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, A.; Terashima, Y.; Toda, E.; Akao, C.; Miyake, Y.; Matsuo, K.; Shimanoe, C.; Nishida, Y.; Hara, M.; Tanaka, K.; et al. Reduced T cell immunity in variant ALDH2 allele carriers. In Proceedings of the 90th Annual Meeting of the Japanese Society for Hygiene, Iwate, Japan, 26–28 March 2020. [Google Scholar]
- Kageyama, T.; Ikeda, K.; Tanaka, S.; Taniguchi, T.; Igari, H.; Onouchi, Y.; Kaneda, A.; Matsushita, K.; Hanaoka, H.; Nakada, T.-A.; et al. Antibody responses to BNT162b2 mRNA COVID-19 vaccine and their predictors among healthcare workers in a tertiary referral hospital in Japan. Clin. Microbiol. Infect. 2021, 27, 1861.e1–1861.e5. [Google Scholar] [CrossRef]
- Rotshild, V.; Hirsh-Raccah, B.; Miskin, I.; Muszkat, M.; Matok, I. Comparing the clinical efficacy of COVID-19 vaccines: A systematic review and network meta-analysis. Sci. Rep. 2021, 11, 22777. [Google Scholar] [CrossRef]
- Zheng, C.; Shao, W.; Chen, X.; Zhang, B.; Wang, G.; Zhang, W. Real-world effectiveness of COVID-19 vaccines: A literature review and meta-analysis. Int. J. Infect. Dis. 2022, 114, 252–260. [Google Scholar] [CrossRef] [PubMed]
- Feikin, D.R.; Higdon, M.M.; Abu-Raddad, L.J.; Andrews, N.; Araos, R.; Goldberg, Y.; Groome, M.J.; Huppert, A.; O’Brien, K.L.; Smith, P.G.; et al. Duration of effectiveness of vaccines against SARS-CoV-2 infection and COVID-19 disease: Results of a systematic review and meta-regression. Lancet 2022, 399, 924–944. [Google Scholar] [CrossRef]
- Steensels, D.; Pierlet, N.; Penders, J.; Mesotten, D.; Heylen, L. Comparison of SARS-CoV-2 antibody response following vaccination with BNT162b2 and mRNA-1273. JAMA 2021, 326, 1533–1535. [Google Scholar] [CrossRef] [PubMed]
- Prendecki, M.; Clarke, C.; Brown, J.; Cox, A.; Gleeson, S.; Guckian, M.; Randell, P.; Pria, A.D.; Lightstone, L.; Xu, X.-N.; et al. Effect of previous SARS-CoV-2 infection on humoral and T-cell responses to single-dose BNT162b2 vaccine. Lancet 2021, 397, 1178–1181. [Google Scholar] [CrossRef]
- Vogel, A.B.; Kanevsky, I.; Che, Y.; Swanson, K.A.; Muik, A.; Vormehr, M.; Kranz, L.M.; Walzer, K.C.; Hein, S.; Guler, A.; et al. BNT162b vaccines protect rhesus macaques from SARS-CoV-2. Nature 2021, 592, 283–289. [Google Scholar] [CrossRef]
- Loyal, L.; Braun, J.; Henze, L.; Kruse, B.; Dingeldey, M.; Reimer, U.; Kern, F.; Schwarz, T.; Mangold, M.; Unger, C.; et al. Cross-reactive CD4+ T cells enhance SARS-CoV-2 immune responses upon infection and vaccination. Science 2021, 374, eabh1823. [Google Scholar] [CrossRef]
- Zuo, J.; Dowell, A.C.; Pearce, H.; Verma, K.; Long, H.M.; Begum, J.; Aiano, F.; Amin-Chowdhury, Z.; Hoschler, K.; Brooks, T.; et al. Robust SARS-CoV-2-specific T cell immunity is maintained at 6 months following primary infection. Nat. Immunol. 2021, 22, 620–626. [Google Scholar] [CrossRef]
- Kroemer, M.; Boullerot, L.; Ramseyer, M.; Spehner, L.; Barisien, C.; Gravelin, E.; Renaudin, A.; Cognasse, F.; Gallian, P.; Hermine, O.; et al. The quality of anti-SARS-CoV-2 T Cell responses predicts the neutralizing antibody titer in convalescent plasma donors. Front. Public Health 2022, 10, 816848. [Google Scholar] [CrossRef] [PubMed]
- Levin, E.G.; Lustig, Y.; Cohen, C.; Fluss, R.; Indenbaum, V.; Amit, S.; Doolman, R.; Asraf, K.; Mendelson, E.; Ziv, A.; et al. Waning immune humoral response to BNT162b2 COVID-19 vaccine over 6 months. N. Engl. J. Med. 2021, 385, e84. [Google Scholar] [CrossRef] [PubMed]
- Ashenagar, M.S.; Matsumoto, A.; Sakai, H.; Tokiya, M.; Hara, M.; Hirota, Y. Comparison of CLEIA and ELISA for SARS-CoV-2 Virus Antibodies after First and Second Dose Vaccinations with the BNT162b2 mRNA Vaccine. Vaccines 2022, 10, 487. [Google Scholar] [CrossRef]
- Zheng, H.; Cao, J.J. Angiotensin-converting enzyme gene polymorphism and severe lung injury in patients with coronavirus disease 2019. Am. J. Pathol. 2020, 190, 2013–2017. [Google Scholar] [CrossRef] [PubMed]
- Deb, P.; Zannat, K.E.; Talukder, S.; Bhuiyan, A.H.; Jilani, M.S.A.; Saif-Ur-Rahman, K.M. Association of HLA gene polymorphism with susceptibility, severity, and mortality of COVID-19: A systematic review. HLA 2022, 99, 281–312. [Google Scholar] [CrossRef]
- Shimanoe, C.; Matsumoto, A.; Hara, M.; Akao, C.; Nishida, Y.; Horita, M.; Nanri, H.; Higaki, Y.; Tanaka, K. Perceived stress, depressive symptoms, and cortisol-to-cortisone ratio in spot urine in 6878 older adults. Psychoneuroendocrinology 2021, 125, 105125. [Google Scholar] [CrossRef]
- Salimans, L.; Liberman, K.; Njemini, R.; Kortekaas Krohn, I.; Gutermuth, J.; Bautmans, I. The effect of resistance exercise on the immune cell function in humans: A systematic review. Exp. Gerontol. 2022, 164, 111822. [Google Scholar] [CrossRef]
- De Spiegeleer, A.; Bronselaer, A.; Teo, J.T.; Byttebier, G.; De Tre, G.; Belmans, L.; Dobson, R.; Wynendaele, E.; Van De Wiele, C.; Vandaele, F.; et al. The effects of ARBs, ACEis, and statins on clinical outcomes of COVID-19 infection among nursing home residents. J. Am. Med. Dir. Assoc. 2020, 21, 909–914.e902. [Google Scholar] [CrossRef]
- Daniels, L.B.; Sitapati, A.M.; Zhang, J.; Zou, J.; Bui, Q.M.; Ren, J.; Longhurst, C.A.; Criqui, M.H.; Messer, K. Relation of statin use prior to admission to severity and recovery among COVID-19 inpatients. Am. J. Cardiol. 2020, 136, 149–155. [Google Scholar] [CrossRef]
- Masaoka, H.; Gallus, S.; Ito, H.; Watanabe, M.; Yokomizo, A.; Eto, M.; Matsuo, K. Aldehyde dehydrogenase 2 polymorphism is a predictor of smoking cessation. Nicotine Tob. Res. 2017, 19, 1087–1094. [Google Scholar] [CrossRef]
- Masaoka, H.; Ito, H.; Gallus, S.; Watanabe, M.; Yokomizo, A.; Eto, M.; Matsuo, K. Combination of ALDH2 and ADH1B polymorphisms is associated with smoking initiation: A large-scale cross-sectional study in a Japanese population. Drug Alcohol Depend. 2017, 173, 85–91. [Google Scholar] [CrossRef] [PubMed]
- Tokiya, M.; Hara, M.; Matsumoto, A.; Ashenagar, M.S.; Nakano, T.; Hirota, Y. Acceptance booster COVID-19 vaccine and its association with components of vaccination readiness in the general Japanese population. Vaccines, 2022; submitted—under revision. [Google Scholar]
- Brunsdon, H.; Brombin, A.; Peterson, S.; Postlethwait, J.H.; Patton, E.E. Aldh2 is a lineage-specific metabolic gatekeeper in melanocyte stem cells. Development 2022, 149, dev200277. [Google Scholar] [CrossRef] [PubMed]
- Balber, A.E. Concise review: Aldehyde dehydrogenase bright stem and progenitor cell populations from normal tissues: Characteristics, activities, and emerging uses in regenerative medicine. Stem Cells 2011, 29, 570–575. [Google Scholar] [CrossRef] [PubMed]
- Garaycoechea, J.I.; Crossan, G.P.; Langevin, F.; Daly, M.; Arends, M.J.; Patel, K.J. Genotoxic consequences of endogenous aldehydes on mouse haematopoietic stem cell function. Nature 2012, 489, 571–575. [Google Scholar] [CrossRef]


{kind=link}
| Participants | Healthcare Workers | University Students | University Employees and Students | ||||||
|---|---|---|---|---|---|---|---|---|---|
| N | 20 | 42 | 26 | ||||||
| First Dose | April 2021 | May 2021 | May 2021 | ||||||
| Type of Vaccine | BNT162b2 | BNT162b2 | mRNA-1273 | ||||||
| Second Dose | Three Weeks after the First Dose | Three Weeks after the First Dose | Four Weeks after the First Dose | ||||||
| GG | GA | AA | GG | GA | AA | GG | GA | AA | |
| Males, N | 3 | 4 | 3 | 11 | 6 | 5 | 3 | 7 | 1 |
| Females, N | 7 | 3 | 0 | 13 | 7 | 0 | 7 | 6 | 2 |
| Age, years | |||||||||
| Median | 42 | 36 | 35 | 22 | 22 | 22 | 39 | 47 | 21 |
| (IQR) | (36–49) | (25–43) | (25–40) | (22–23) | (22–23) | (22–23) | (22–56) | (21–55) | (21–42) |
| Body height, cm | |||||||||
| Median | 164 | 168 | 170 | 165.5 | 162 | 172 | 162 | 166 | 157 |
| (IQR) | (162–169) | (153.8–178) | (170–176) | (158–170.5) | (156–174) | (170–173) | (155–166) | (163–170) | (151–163) |
| Smoking status, yes | 1 | 1 | 1 | 0 | 0 | 0 | 0 | 1 | 0 |
| Ethanol intake * | |||||||||
| <1 g/d | 1 | 4 | 3 | 12 | 8 | 4 | 6 | 9 | 3 |
| ≥1, <20 g/d | 7 | 3 | 0 | 12 | 5 | 1 | 4 | 2 | 0 |
| ≥20 g/d | 2 | 0 | 0 | 0 | 0 | 0 | 0 | 2 | 0 |
| Exercise habit | |||||||||
| No habit | 8 | 2 | 1 | 10 | 4 | 2 | 3 | 4 | 1 |
| <1 d/w | 0 | 0 | 1 | 5 | 1 | 1 | 2 | 2 | 0 |
| 1 to 3 d/w | 2 | 2 | 1 | 5 | 6 | 2 | 2 | 4 | 1 |
| ≥3 d/w | 0 | 3 | 0 | 4 | 2 | 0 | 3 | 3 | 1 |
| Perceived stress | |||||||||
| 0 (no) | 2 | 3 | 1 | 12 | 5 | 1 | 1 | 5 | 1 |
| 1 | 0 | 0 | 1 | 2 | 1 | 0 | 3 | 1 | 0 |
| 2 | 3 | 1 | 1 | 4 | 4 | 3 | 2 | 2 | 0 |
| 3 | 4 | 2 | 0 | 6 | 3 | 1 | 4 | 4 | 0 |
| 4 (yes) | 1 | 1 | 0 | 0 | 0 | 0 | 0 | 1 | 2 |
| Steroid use, yes | 0 | 0 | 0 | 2 | 0 | 0 | 0 | 0 | 0 |
| Allergic disease, yes | 2 | 4 | 0 | 11 | 3 | 1 | 2 | 6 | 1 |
| Dyslipidemia | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 |
| Healthcare Workers | University Students | University Employees and Students | |||||||
|---|---|---|---|---|---|---|---|---|---|
| BNT162b2 (Second Dose at Week 3) | BNT162b2 (Second Dose at Week 3) | mRNA-1273 (Second Dose at Week 4) | |||||||
| Genotype | GG | GA | AA | GG | GA | AA | GG | GA | AA |
| Week 0 | |||||||||
| N | 10 | 7 | 3 | 24 | 13 | 5 | 10 | 13 | 3 |
| Median | 0.37 | 0.49 | 0.41 | 0.39 | 0.46 | 0.35 | 0.49 | 0.46 | 0.34 |
| (IQR) | (0.33–0.5) | (0.45–0.58) | (0.4–0.53) | (0.33–0.52) | (0.35–0.57) | (0.28–0.39) | (0.3–0.64) | (0.41–0.52) | (0.3–0.62) |
| Week 1 | 1 week after the first dose | ||||||||
| N | 10 | 7 | 3 | ||||||
| Median | 0.62 | 0.54 | 0.5 | ||||||
| (IQR) | (0.37–1.76) | (0.51–1.15) | (0.37–7.2) | ||||||
| Week 2 | 2 weeks after the first dose | ||||||||
| N | 10 | 6 | 3 | ||||||
| Median | 64 | 46 | 50 | ||||||
| (IQR) | (44–89) | (18–76) | (5.71–95) | ||||||
| Week 3 | 3 weeks after the first dose | 3 weeks after the first dose | 3 weeks after the first dose | ||||||
| N | 10 | 6 | 3 | 24 | 13 | 5 | 10 | 13 | 3 |
| Median | 187 | 92 | 122 | 132 | 134 | 122 | 372 | 214 | 560 |
| (IQR) | (101–295) | (70–113) | (56–153) | (75–210) | (91–200) | (105–135) | (290–393) | (143–320) | (410–572) |
| Week 4 | 1 week after the second dose | ||||||||
| N | 9 | 6 | 3 | ||||||
| Median | 1498 | 629 | 898 | ||||||
| (IQR) | (691–2529) | (576–1271) | (241–909) | ||||||
| Week 5 | 2 weeks after the second dose | ||||||||
| N | 10 | 6 | 3 | ||||||
| Median | 2482 | 1694 | 725 | ||||||
| (IQR) | (1901–2667) | (956–2523) | (592–1594) | ||||||
| Week 6 | 3 weeks after the second dose | ||||||||
| N | 10 | 6 | 3 | ||||||
| Median | 1958 | 1372 | 564 | ||||||
| (IQR) | (1507–2113) | (918–1975) | (558–1350) | ||||||
| Week 7 | 1 month after the second dose | 1 month after the second dose | |||||||
| N | 10 | 6 | 3 | 24 | 13 | 5 | |||
| Median | 1592 | 1109 | 510 | 1597 | 1880 | 1339 | |||
| (IQR) | (1129–1779) | (619–1658) | (433–818) | (1198–2269) | (1461–2292) | (1174–1680) | |||
| Week 8 | 1 month after the second dose | ||||||||
| N | 10 | 13 | 3 | ||||||
| Median | 3200 | 2959 | 2854 | ||||||
| (IQR) | (2756–3681) | (1661–3593) | (1362–3565) | ||||||
| Week 11 | 2 months after the second dose | 2 months after the second dose | |||||||
| N | 10 | 6 | 3 | 24 | 12 | 5 | |||
| Median | 761 | 526 | 337 | 940 | 859 | 632 | |||
| (IQR) | (493–854) | (419–819) | (228–523) | (560–1080) | (534–1133) | (572–752) | |||
| Week 15 | 3 months after the second dose | 3 months after the second dose | |||||||
| N | 10 | 6 | 3 | 24 | 12 | 5 | |||
| Median | 361 | 341 | 162 | 815 | 680 | 426 | |||
| (IQR) | (271–486) | (300–402) | (113–331) | (464–1087) | (457–1085) | (418–677) | |||
| Week 16 | 3 months after the second dose | ||||||||
| N | 10 | 13 | 3 | ||||||
| Median | 1579 | 1227 | 1268 | ||||||
| (IQR) | (1220–1740) | (742–1906) | (983–1746) | ||||||
| Model 1 | Model 2 | Model 3 | ||||
|---|---|---|---|---|---|---|
| AIC = 1007 | AIC = 993 | AIC = 997 | ||||
| 503 Observations | 503 Observations | 503 Observations | ||||
| 88 Subjects | 88 Subjects | 88 Subjects | ||||
| Fixed Effects | β | p-Value | β | p-Value | β | p-Value |
| BNT162b2 (reference) | ||||||
| mRNA-1273 | 0.53 | 0.0004 | 0.50 | <0.0001 | 0.48 | <0.0001 |
| Age (per year old) | −0.01 | 0.0087 | −0.01 | 0.0007 | −0.01 | 0.0030 |
| Female sex | 0.21 | 0.0013 | 0.07 | 0.4522 | 0.05 | 0.6345 |
| Height (per cm) | −0.01 | 0.2000 | −0.01 | 0.1725 | ||
| Smoking status, yes | 0.20 | 0.1138 | 0.20 | 0.1039 | ||
| Ethanol intake (per category) | −0.05 | 0.3908 | ||||
| Exercise habit (per category) | −0.03 | 0.2716 | −0.03 | 0.2802 | ||
| Perceived stress (per category) | 0.07 | 0.0043 | 0.07 | 0.0041 | ||
| Steroid use, yes | −0.07 | 0.7518 | −0.09 | 0.6877 | ||
| Allergic disease, yes | −0.04 | 0.5208 | −0.04 | 0.4938 | ||
| Dyslipidemia, yes | −1.02 | <0.0001 | −1.02 | <0.0001 | ||
| ALDH2 variant allele number | −0.11 | 0.0116 | −0.13 | 0.0021 | −0.15 | 0.0016 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Matsumoto, A.; Hara, M.; Ashenagar, M.S.; Tokiya, M.; Sawada, T.; Iwasaka, C.; Furukawa, T.; Kitagawa, K.; Miyake, Y.; Hirota, Y. Variant Allele of ALDH2, rs671, Associates with Attenuated Post-Vaccination Response in Anti-SARS-CoV-2 Spike Protein IgG: A Prospective Study in the Japanese General Population. Vaccines 2022, 10, 1035. https://doi.org/10.3390/vaccines10071035
Matsumoto A, Hara M, Ashenagar MS, Tokiya M, Sawada T, Iwasaka C, Furukawa T, Kitagawa K, Miyake Y, Hirota Y. Variant Allele of ALDH2, rs671, Associates with Attenuated Post-Vaccination Response in Anti-SARS-CoV-2 Spike Protein IgG: A Prospective Study in the Japanese General Population. Vaccines. 2022; 10(7):1035. https://doi.org/10.3390/vaccines10071035
Chicago/Turabian StyleMatsumoto, Akiko, Megumi Hara, Mohammad Said Ashenagar, Mikiko Tokiya, Takeshi Sawada, Chiharu Iwasaka, Takuma Furukawa, Kyoko Kitagawa, Yasunobu Miyake, and Yoshio Hirota. 2022. "Variant Allele of ALDH2, rs671, Associates with Attenuated Post-Vaccination Response in Anti-SARS-CoV-2 Spike Protein IgG: A Prospective Study in the Japanese General Population" Vaccines 10, no. 7: 1035. https://doi.org/10.3390/vaccines10071035
APA StyleMatsumoto, A., Hara, M., Ashenagar, M. S., Tokiya, M., Sawada, T., Iwasaka, C., Furukawa, T., Kitagawa, K., Miyake, Y., & Hirota, Y. (2022). Variant Allele of ALDH2, rs671, Associates with Attenuated Post-Vaccination Response in Anti-SARS-CoV-2 Spike Protein IgG: A Prospective Study in the Japanese General Population. Vaccines, 10(7), 1035. https://doi.org/10.3390/vaccines10071035