
DNA-Vaccine-Induced Immune Response Correlates with Lower Viral SARS-CoV-2 Titers in a Ferret Model

 , , , , , , , , , , , and
, , , , , , , , , , , and
Abstract
:1. Introduction
2. Material and Methods
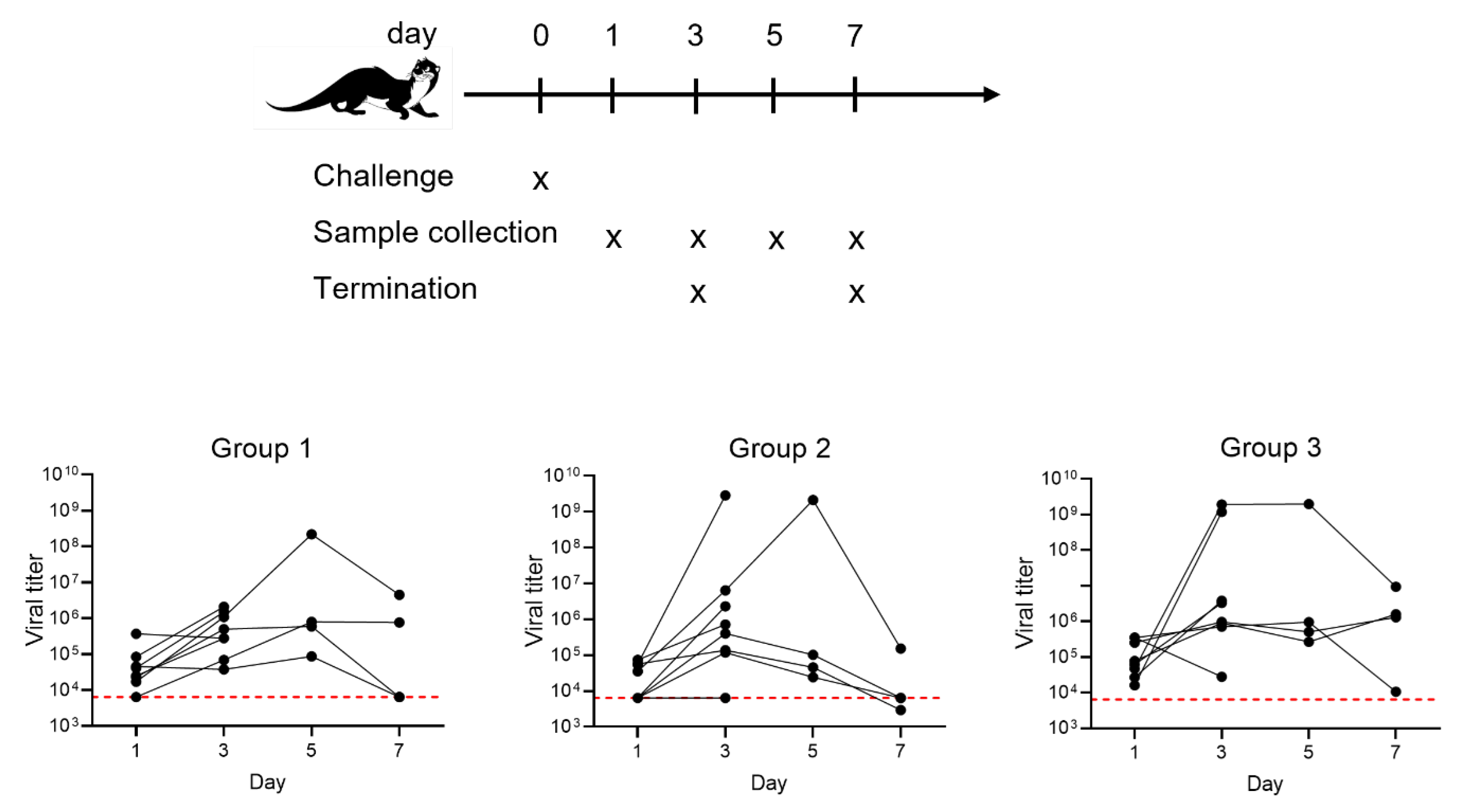
2.1. Vaccination and Viral Challenge
2.2. Clinical Data
2.3. Molecular Tests
2.4. ELISA Assay
2.5. ELISpot Assay
2.6. Neutralization Assay
2.7. Necropsy Procedure
2.8. Pathological Studies
2.9. Statistical Analysis
3. Results
3.1. COVID-eVax-Induced Immune Responses in Ferrets
3.2. Protection against SARS-CoV-2 and Neutralization of VOCs
3.3. Clinical Signs
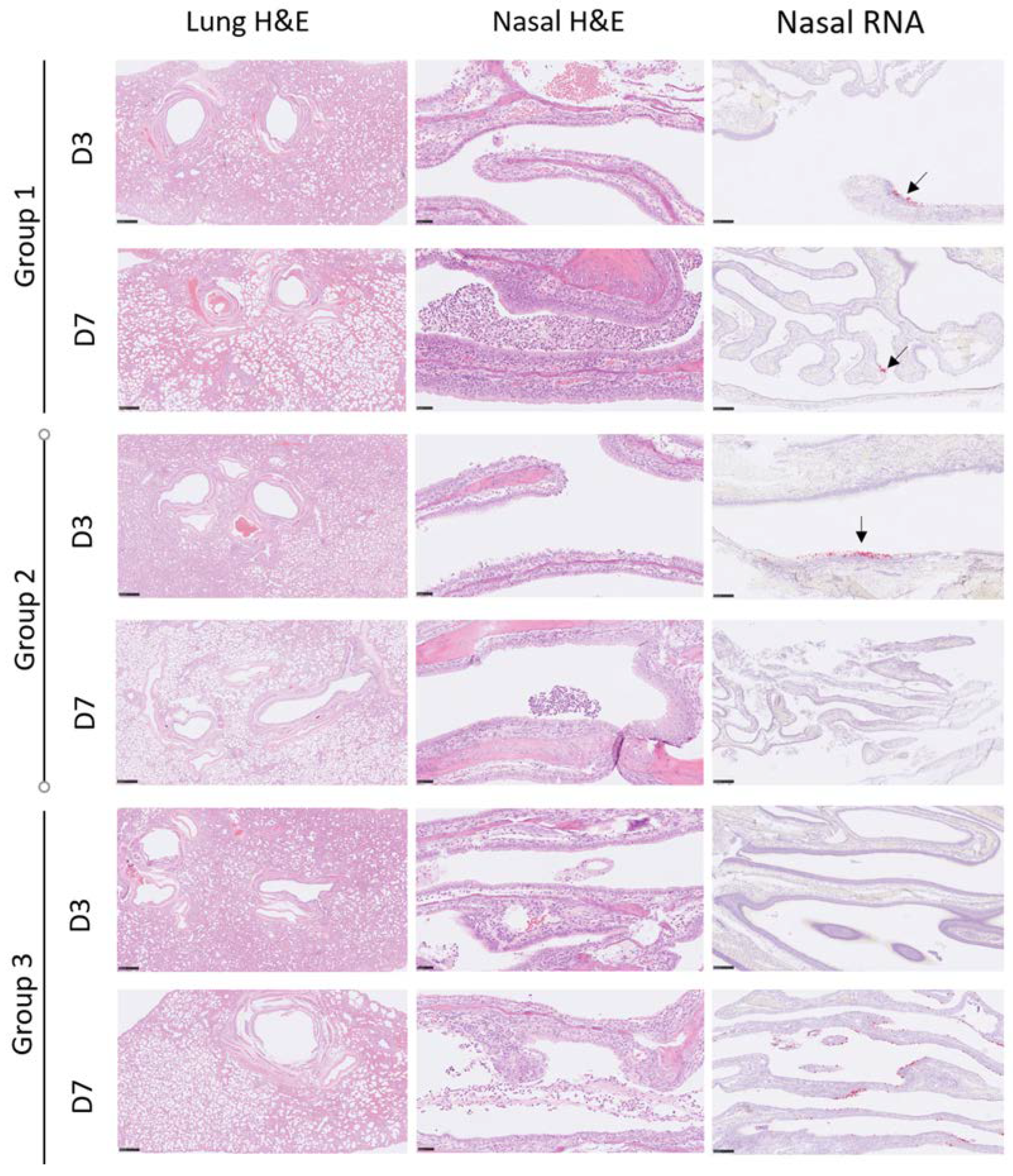
3.4. Histopathology
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhou, P.; Yang, X.-L.; Wang, X.-G.; Hu, B.; Zhang, L.; Zhang, W.; Si, H.-R.; Zhu, Y.; Li, B.; Huang, C.-L.; et al. A pneumonia outbreak associated with a new coronavirus of probable bat origin. Nature 2020, 579, 270–273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, W.; Moore, M.J.; Vasilieva, N.; Sui, J.; Wong, S.K.; Berne, M.A.; Somasundaran, M.; Sullivan, J.L.; Luzuriaga, K.; Greenough, T.C.; et al. Angiotensin-converting enzyme 2 is a functional receptor for the SARS coronavirus. Nature 2003, 426, 450–454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cleary, S.J.; Pitchford, S.C.; Amison, R.T.; Carrington, R.; Robaina Cabrera, C.L.; Magnen, M.; Looney, M.R.; Gray, E.; Page, C.P. Animal models of mechanisms of SARS-CoV-2 infection and COVID-19 pathology. Br. J. Pharmacol. 2020, 177, 4851–4865. [Google Scholar] [CrossRef] [PubMed]
- Muñoz-Fontela, C.; Dowling, W.E.; Funnell, S.G.P.; Gsell, P.-S.; Riveros-Balta, A.X.; Albrecht, R.A.; Andersen, H.; Baric, R.S.; Carroll, M.W.; Cavaleri, M.; et al. Animal models for COVID-19. Nature 2020, 586, 509–515. [Google Scholar] [CrossRef]
- Richard, M.; Kok, A.; de Meulder, D.; Bestebroer, T.M.; Lamers, M.M.; Okba, N.M.A.; Fentener van Vlissingen, M.; Rockx, B.; Haagmans, B.L.; Koopmans, M.P.G.; et al. SARS-CoV-2 is transmitted via contact and via the air between ferrets. Nat. Commun. 2020, 11, 3496. [Google Scholar] [CrossRef]
- Au, G.G.; Marsh, G.A.; McAuley, A.J.; Lowther, S.; Trinidad, L.; Edwards, S.; Todd, S.; Barr, J.; Bruce, M.P.; Poole, T.B.; et al. Characterisation and natural progression of SARS-CoV-2 infection in ferrets. Sci. Rep. 2022, 12, 5680. [Google Scholar] [CrossRef] [PubMed]
- Conforti, A.; Marra, E.; Palombo, F.; Roscilli, G.; Ravà, M.; Fumagalli, V.; Muzi, A.; Maffei, M.; Luberto, L.; Lione, L.; et al. COVID-eVax, an electroporated DNA vaccine candidate encoding the SARS-CoV-2 RBD, elicits protective responses in animal models. Mol. Ther. 2022, 30, 311–326. [Google Scholar] [CrossRef] [PubMed]
- Elia, L.; Mennuni, C.; Storto, M.; Podda, S.; Calvaruso, F.; Salucci, V.; Aurisicchio, L.; Scarito, A.; Ciliberto, G.; La Monica, N.; et al. Genetic vaccines against Ep-CAM break tolerance to self in a limited subset of subjects: Initial identification of predictive biomarkers. Eur. J. Immunol. 2006, 36, 1337–1349. [Google Scholar] [CrossRef] [PubMed]
- Aurisicchio, L.; Salvatori, E.; Lione, L.; Bandini, S.; Pallocca, M.; Maggio, R.; Fanciulli, M.; De Nicola, F.; Goeman, F.; Ciliberto, G.; et al. Poly-specific neoantigen-targeted cancer vaccines delay patient derived tumor growth. J. Exp. Clin. Cancer Res. 2019, 4, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Lione, L.; Salvatori, E.; Petrazzuolo, A.; Massacci, A.; Maggio, R.; Confroti, A.; Compagnone, M.; Aurisicchio, L.; Ciliberto, G.; Palombo, F. Antitumor efficacy of a neoantigen cancer vaccine delivered by electroporation is influenced by microbiota composition. Oncoimmunology 2021, 10, 1898832. [Google Scholar] [CrossRef] [PubMed]
- Salvatori, E.; Lione, L.; Compagnone, M.; Pinto, E.; Conforti, A.; Ciliberto, G.; Aurisicchio, L.; Palombo, F. Neoantigen cancer vaccine augments anti-CTLA-4 efficacy. NPJ Vaccines 2022, 7, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Khobragade, A.; Bhate, S.; Ramaiah, V.; Deshpande, S.; Giri, K.; Phophle, H.; Supe, P.; Godara, I.; Revanna, R.; Nagarkar, R.; et al. Efficacy, safety, and immunogenicity of the DNA SARS-CoV-2 vaccine (ZyCoV-D): The interim efficacy results of a phase 3, randomised, double-blind, placebo-controlled study in India. Lancet 2022, 399, 1313–1321. [Google Scholar] [CrossRef]
- Shi, J.; Wen, Z.; Zhong, G.; Yang, H.; Wang, C.; Huang, B.; Liu, R.; He, X.; Shuai, L.; Sun, Z.; et al. Susceptibility of ferrets, cats, dogs, and other domesticated animals to SARS-coronavirus 2. Science 2020, 368, 1016–1020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, E.J.; Poole, K.C.; Sollini, J.; Town, S.M.; Bizley, J.K. Seasonal weight changes in laboratory ferrets. PLoS ONE 2020, 15, e0232733. [Google Scholar] [CrossRef] [PubMed]
- Chenchula, S.; Karunakaran, P.; Sharma, S.; Chavan, M. Current evidence on efficacy of COVID-19 booster dose vaccination against the Omicron variant: A systematic review. J. Med. Virol. 2022, 94, 2969–2976. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Day Zero | Day Seven | ||||||
|---|---|---|---|---|---|---|---|
| Fer. | Wt | Delta | Omi | Wt | Delta | Omi | |
| 1 dose | 1 | 51 | NN | NN | - | - | - |
| 2 | 35 | NN | NN | 109 | NN | NN | |
| 3 | NN | NN | NN | NN | NN | NN | |
| 4 | NN | NN | NN | 36 | NN | NN | |
| 5 | NN | NN | NN | 139 | 166 | 77 | |
| 6 | NN | NN | NN | - | - | - | |
| 7 | 35 | NN | NN | - | - | - | |
| 8 | NN | NN | NN | - | - | - | |
| 2 doses | 9 | NN | NN | NN | - | - | - |
| 10 | NN | NN | NN | - | - | - | |
| 11 | 93 | NN | NN | 100 | 45 | 40 | |
| 12 | 3062 | 507 | 153 | 154 | 64 | NN | |
| 13 | 224 | 44 | NN | 211 | 101 | NN | |
| 14 | 64 | 46 | NN | 407 | 60 | NN | |
| 15 | 1974 | 607 | 46 | - | - | - | |
| 16 | NN | NN | NN | - | - | - | |
| not vac. | 17 | NN | NN | NN | - | - | - |
| 18 | NN | NN | NN | - | - | - | |
| 19 | NN | NN | NN | - | - | - | |
| 20 | NN | NN | NN | - | - | - | |
| 21 | - | - | - | NN | NN | NN | |
| 22 | - | - | - | NN | NN | NN | |
| 23 | - | - | - | NN | NN | NN | |
| 24 | - | - | - | NN | NN | NN |
| Group | Days Pc | Lungs | Nasal Cavity | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Bronchial Inflammation (Bronchitis) | Bronchiolar Inflammation (Bronchiolitis) | PVC | Inflammation of Alveoli | Average Lung Scores | Viral Staining Intensity | Epithelial Inflammation/Necrosis | Lumenal Exudate | Viral Staining Intensity (SARS-CoV-2) | ||
| (SARS-CoV-2) | ||||||||||
| Group 1 | 1 | 2 | 0.5 | 1 | 1.1 | 0 | 2 | 1 | 1 | |
| (one dose) | 0.5 | 1 | 1 | 1 | 0.9 | 0 | 2 | 0 | 0 | |
| 0.5 | 1.5 | 1.5 | 1 | 1.1 | 0 | 1 | 0 | 0 | ||
| 3 | 1.5 | 2 | 1.5 | 1.5 | 1.6 | 0 | 0 | 0 | 0 | |
| 1.5 | 1.5 | 1.5 | 1.5 | 1.5 | 0 | 1 | 0 | 0 | ||
| 0.5 | 2 | 1.5 | 1 | 1.3 | 0 | 2 | 2 | 0 | ||
| 1.5 | 1.5 | 2 | 1 | 1.5 | 0 | 2 | 2 | 1 | ||
| 7 | 1.5 | 0.5 | 0.5 | 0.5 | 0.8 | 0 | 0 | 0 | 1 | |
| Group 2 | 1.5 | 2.5 | 1.5 | 2 | 1.9 | 0 | 0 | 0 | 2 | |
| (two doses) | 1.5 | 2 | 0.5 | 1 | 1.3 | 0 | 2 | 1 | 3 | |
| 1 | 1.5 | 1 | 1.5 | 1.3 | 0 | 1 | 1 | 1 | ||
| 3 | 1.5 | 2 | 1.5 | 2 | 1.8 | 0 | 0 | 0 | 0 | |
| 1.5 | 1 | 0.5 | 1 | 1.0 | 0 | 1 | 1 | 0 | ||
| 1.5 | 1.5 | 0.5 | 1 | 1.1 | 0 | 1 | 1 | 0 | ||
| 1 | 1 | 0.5 | 1 | 0.9 | 0 | 1 | 1 | 0 | ||
| 7 | 0.5 | 1.5 | 0.5 | 1 | 0.9 | 0 | 1 | 1 | 0 | |
| Group 3 | 1.5 | 1.5 | 0.5 | 1 | 1.1 | 0 | 0 | 1 | 0 | |
| (unvaccinated) | 1.5 | 2 | 1 | 2 | 1.6 | 0 | 1 | 1 | 0 | |
| 1.5 | 2.5 | 2 | 1.5 | 1.9 | 0 | 0 | 0 | 0 | ||
| 3 | 1.5 | 1.5 | 1.5 | 1.5 | 1.5 | 0 | 1 | 0 | 0 | |
| 1 | 2 | 0.5 | 1 | 1.1 | 0 | 2 | 2 | 0 | ||
| 1 | 1.5 | 1 | 1.5 | 1.3 | 0 | 1 | 1 | 0 | ||
| 1 | 1 | 0.5 | 1 | 0.9 | 0 | 1 | 1 | 2 | ||
| 7 | 1 | 1.5 | 1 | 1 | 1.1 | 0 | 2 | 1 | 3 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Compagnone, M.; Pinto, E.; Salvatori, E.; Lione, L.; Conforti, A.; Marchese, S.; Ravà, M.; Ryan, K.; Hall, Y.; Rayner, E.; et al. DNA-Vaccine-Induced Immune Response Correlates with Lower Viral SARS-CoV-2 Titers in a Ferret Model. Vaccines 2022, 10, 1178. https://doi.org/10.3390/vaccines10081178
Compagnone M, Pinto E, Salvatori E, Lione L, Conforti A, Marchese S, Ravà M, Ryan K, Hall Y, Rayner E, et al. DNA-Vaccine-Induced Immune Response Correlates with Lower Viral SARS-CoV-2 Titers in a Ferret Model. Vaccines. 2022; 10(8):1178. https://doi.org/10.3390/vaccines10081178
Chicago/Turabian StyleCompagnone, Mirco, Eleonora Pinto, Erika Salvatori, Lucia Lione, Antonella Conforti, Silvia Marchese, Micol Ravà, Kathryn Ryan, Yper Hall, Emma Rayner, and et al. 2022. "DNA-Vaccine-Induced Immune Response Correlates with Lower Viral SARS-CoV-2 Titers in a Ferret Model" Vaccines 10, no. 8: 1178. https://doi.org/10.3390/vaccines10081178
APA StyleCompagnone, M., Pinto, E., Salvatori, E., Lione, L., Conforti, A., Marchese, S., Ravà, M., Ryan, K., Hall, Y., Rayner, E., Salguero, F. J., Paterson, J., Iannacone, M., De Francesco, R., Aurisicchio, L., & Palombo, F. (2022). DNA-Vaccine-Induced Immune Response Correlates with Lower Viral SARS-CoV-2 Titers in a Ferret Model. Vaccines, 10(8), 1178. https://doi.org/10.3390/vaccines10081178