
Protection against Bovine Respiratory Syncytial Virus Afforded by Maternal Antibodies from Cows Immunized with an Inactivated Vaccine
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Virus and Inoculum
2.2. Experimental Design
2.2.1. Vaccination of Cows and Colostrum Collection
2.2.2. Infection of Calves
2.3. Clinical Evaluation
2.4. Virological Examination
2.5. Evaluation of the Host Response
2.5.1. Humoral Response
2.5.2. Transcriptomic Response in BAL
2.6. Statistical Analysis
3. Results
3.1. Vaccination Induces a Strong BRSV Antibody Response in Cows and a High BRSV Antibody Titers in Colostrum and Colostrum-Fed Calves
3.2. Colostrum from the BRSV-Vaccinated Cows Protect Calves against Severe Disease after the Challenge
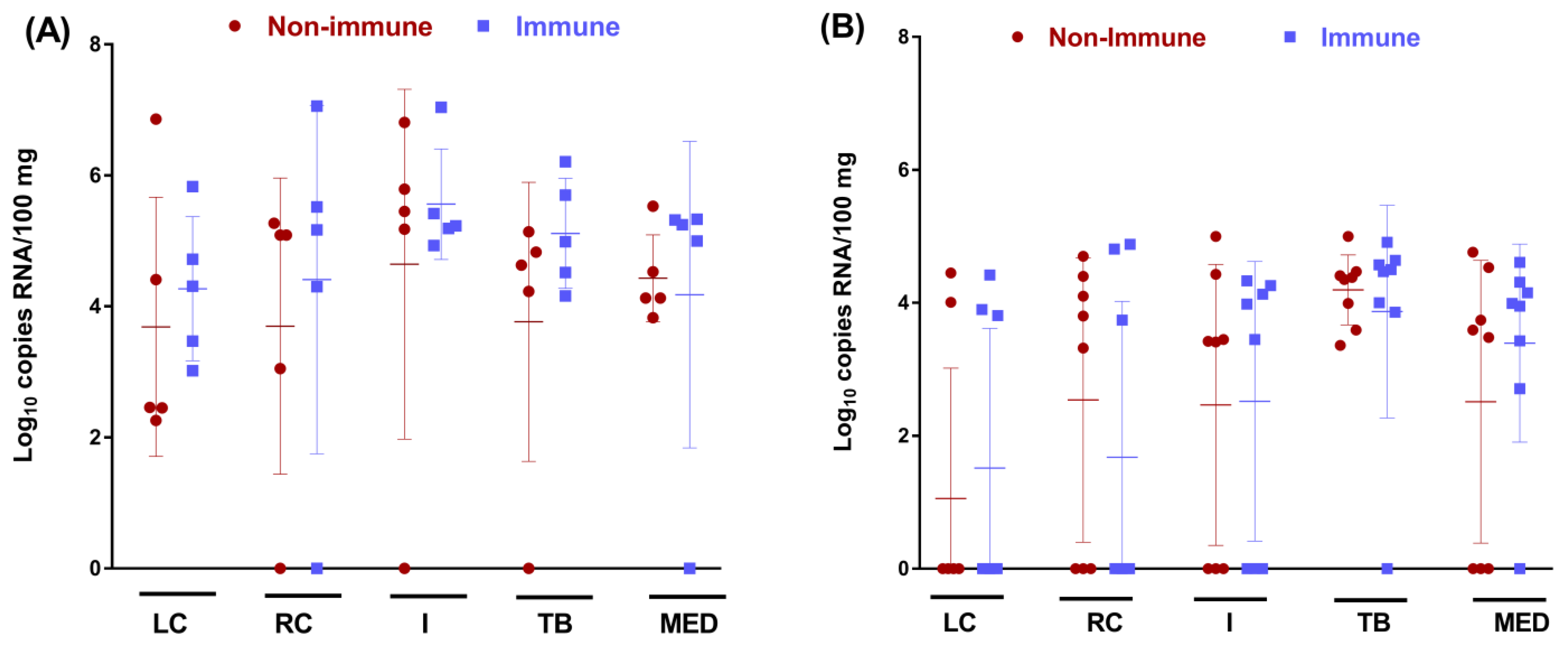
3.3. Colostrum from BRSV-Vaccinated Cows Does Not Protect Calves against BRSV Infection of the URT but Reduces the Viral Load in the Lung
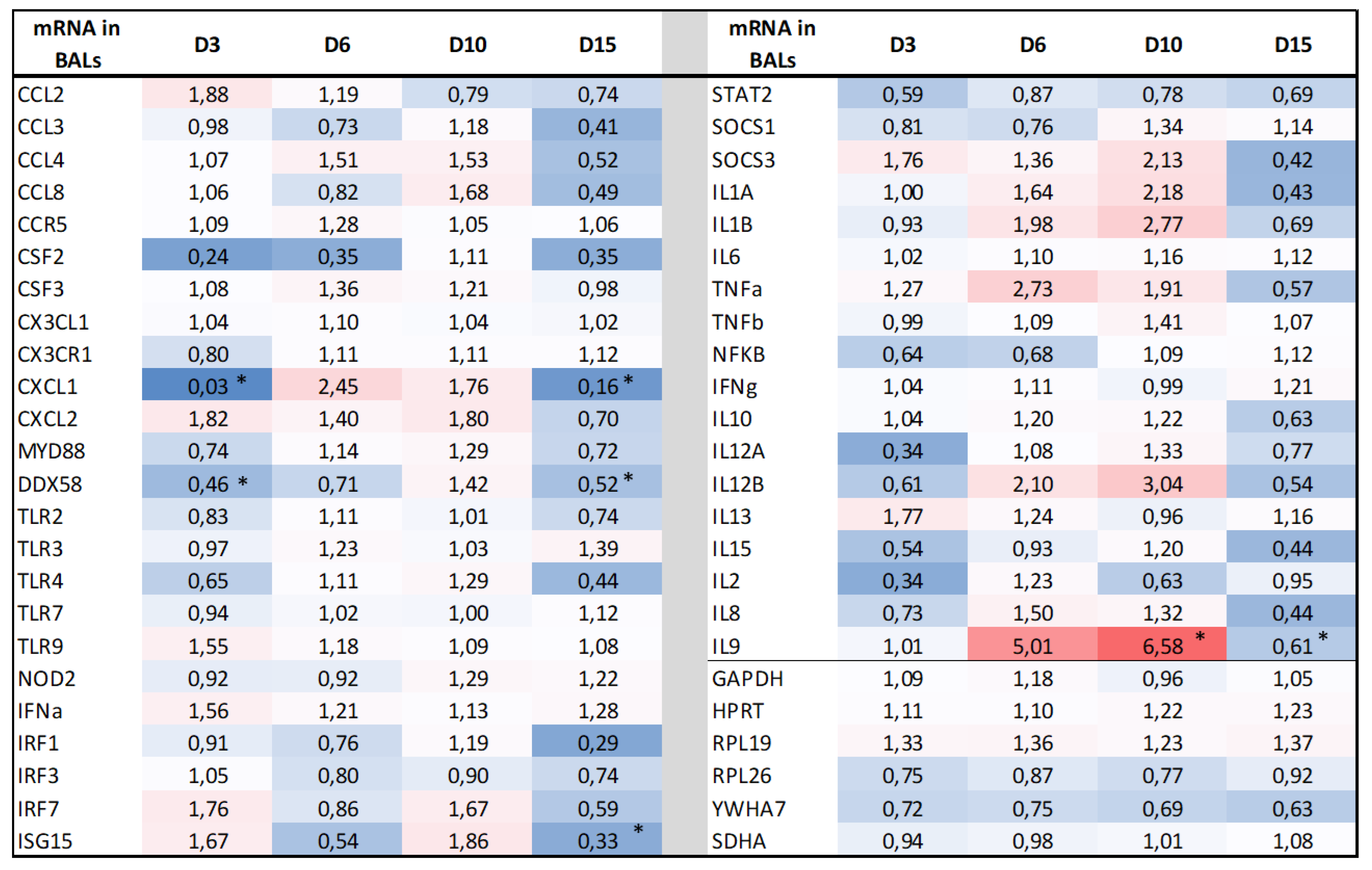
3.4. BRSV Maternal Antibodies Do Not Modify the Transcriptomic Host Response in the Lungs after the BRSV Challenge
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- International Committee on Taxonomy of Viruses (ICTV). Virus Taxonomy. 2021. Available online: https://ictv.global/taxonomy/taxondetails?taxnode_id=202101650 (accessed on 19 November 2022).
- Gaudino, M.; Nagamine, B.; Ducatez, M.F.; Meyer, G. Understanding the mechanisms of viral and bacterial coinfections in bovine respiratory disease: A comprehensive literature review of experimental evidence. Vet. Res. 2022, 53, 70. [Google Scholar] [CrossRef] [PubMed]
- Beaudeau, F.; Ohlson, A.; Emanuelson, U. Associations between bovine coronavirus and bovine respiratory syncytial virus infections and animal performance in Swedish dairy herds. J. Dairy Sci. 2010, 93, 1523–1533. [Google Scholar] [CrossRef] [Green Version]
- Johnson, K.K.; Pendell, D.L. Market impacts of reducing the prevalence of bovine respiratory disease in United States Beef Cattle Feedlots. Front. Vet. Sci. 2017, 4, 189. [Google Scholar] [CrossRef] [Green Version]
- Woolums, A.R.; Berghaus, R.D.; Smith, D.R.; White, B.J.; Engelken, T.J.; Irsik, M.B.; Matlick, D.K.; Jones, A.L.; Ellis, R.W.; Smith, I.J.; et al. Producer survey of herd-level risk factors for nursing beef calf respiratory disease. J. Am. Vet. Med. Assoc. 2013, 243, 538–547. [Google Scholar] [CrossRef] [PubMed]
- Chamorro, M.F.; Woolums, A.; Walz, P.H. Vaccination of calves against common respiratory viruses in the face of maternally derived antibodies (IFOMA). Anim. Health Res. Rev. 2016, 17, 79–84. [Google Scholar] [CrossRef]
- Makoschey, B.; Berge, A.C. Review on bovine respiratory syncytial virus and bovine parainfluenza—Usual suspects in bovine respiratory disease—A narrative review. BMC Vet. Res. 2021, 17, 261. [Google Scholar] [CrossRef]
- Ellis, J.A. How efficacious are vaccines against bovine respiratory syncytial virus in cattle? Vet. Microbiol. 2017, 206, 59–68. [Google Scholar] [CrossRef] [PubMed]
- Blodorn, K.; Hagglund, S.; Fix, J.; Dubuquoy, C.; Makabi-Panzu, B.; Thom, M.; Karlsson, P.; Roque, J.L.; Karlstam, E.; Pringle, J.; et al. Vaccine safety and efficacy evaluation of a recombinant bovine respiratory syncytial virus (BRSV) with deletion of the SH gene and subunit vaccines based on recombinant human RSV proteins: N-nanorings, P and M2-1, in calves with maternal antibodies. PLoS ONE 2014, 9, e100392. [Google Scholar] [CrossRef]
- Ellis, J.A.; Hassard, L.E.; Cortese, V.S.; Morley, P.S. Effects of perinatal vaccination on humoral and cellular immune responses in cows and young calves. J. Am. Vet. Med. Assoc. 1996, 208, 393–400. [Google Scholar]
- Ellis, J.A.; Gow, S.P.; Goji, N. Response to experimentally induced infection with bovine respiratory syncytial virus following intranasal vaccination of seropositive and seronegative calves. J. Am. Vet. Med. Assoc. 2010, 236, 991–999. [Google Scholar] [CrossRef]
- Ellis, J.; Gow, S.; Bolton, M.; Burdett, W.; Nordstrom, S. Inhibition of priming for bovine respiratory syncytial virus-specific protective immune responses following parenteral vaccination of passively immune calves. Can. Vet. J. 2014, 55, 1180–1185. [Google Scholar]
- Hagglund, S.; Hu, K.F.; Larsen, L.E.; Hakhverdyan, M.; Valarcher, J.F.; Taylor, G.; Morein, B.; Belak, S.; Alenius, S. Bovine respiratory syncytial virus ISCOMs—Protection in the presence of maternal antibodies. Vaccine 2004, 23, 646–655. [Google Scholar] [CrossRef] [PubMed]
- Harmeyer, S.S.; Murray, J.; Imrie, C.; Wiseman, A.; Salt, J.S. Efficacy of a live bovine respiratory syncytial virus vaccine in seropositive calves. Vet. Rec. 2006, 159, 456–457. [Google Scholar] [CrossRef]
- Kolb, E.A.; Buterbaugh, R.E.; Rinehart, C.L.; Ensley, D.; Perry, G.A.; Abdelsalam, K.W.; Chase, C.C.L. Protection against bovine respiratory syncytial virus in calves vaccinated with adjuvanted modified live vaccine administered in the face of maternal antibody. Vaccine 2020, 38, 298–308. [Google Scholar] [CrossRef]
- Patel, J.R.; Didlick, S.A. Evaluation of efficacy of an inactivated vaccine against bovine respiratory syncytial virus in calves with maternal antibodies. Am. J. Vet. Res. 2004, 65, 417–421. [Google Scholar] [CrossRef] [PubMed]
- Riffault, S.; Hagglund, S.; Guzman, E.; Naslund, K.; Jouneau, L.; Dubuquoy, C.; Pietralunga, V.; Laubreton, D.; Boulesteix, O.; Gauthier, D.; et al. A Single Shot Pre-fusion-Stabilized Bovine RSV F Vaccine is Safe and Effective in Newborn Calves withMaternally Derived Antibodies. Vaccines 2020, 8, 231. [Google Scholar] [CrossRef]
- Valarcher, J.F.; Hägglund, S.; Näslund, K.; Jouneau, L.; Malmström, E.; Boulesteix, O.; Pinard, A.; Leguéré, D.; Deslis, A.; Gauthier, D.; et al. Single-Shot Vaccines against Bovine Respiratory Syncytial Virus (BRSV): Comparative Evaluation of Long-Term Protection after Immunization in the Presence of BRSV-Specific Maternal Antibodies. Vaccines 2021, 9, 236. [Google Scholar] [CrossRef] [PubMed]
- Vangeel, I.; Antonis, A.F.G.; Fluess, M.; Riegler, L.; Peters, A.R.; Harmeyer, S.S. Efficacy of a modified live intranasal bovine respiratory syncytial virus vaccine in 3-week-old calves experimentally challenged with BRSV. Vet. J. 2007, 174, 627–635. [Google Scholar] [CrossRef]
- Kimman, T.G.; Westenbrink, F.; Straver, P.J. Priming for local and systemic antibody memory responses to bovine respiratory syncytial virus: Effect of amount of virus, virus replication, route of administration and maternal antibodies. Vet. Immunol. Immunopathol. 1989, 22, 145–160. [Google Scholar] [CrossRef]
- Baker, J.C.; Ames, T.R.; Markhamv, R.J. Seroepizootiologic study of bovine respiratory syncytial virus in a dairy herd. Am. J. Vet. Res. 1986, 47, 240–245. [Google Scholar]
- Kimman, T.G.; Westenbrink, F.; Schreuder, B.E.; Straver, P.J. Local and systemic antibody response to bovine respiratory syncytial virus infection and reinfection in calves with and without maternal antibodies. J. Clin. Microbiol. 1987, 25, 1097–1106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kimman, T.G.; Zimmer, G.M.; Westenbrink, F.; Mars, J.; van Leeuwen, E. Epidemiological study of bovine respiratory syncytial virus infections in calves: Influence of maternal antibodies on the outcome of disease. Vet. Rec. 1988, 123, 104–109. [Google Scholar] [CrossRef] [PubMed]
- Belknap, E.B.; Baker, J.C.; Patterson, J.S.; Walker, R.D.; Haines, D.M.; Clark, E.G. The role of passive immunity in bovine respiratory syncytial virus-infected calves. J. Infect. Dis. 1991, 163, 470–476. [Google Scholar] [CrossRef]
- Baker, J.C.; Ellis, J.A.; Clark, E.G. Bovine respiratory syncytial virus. Vet. Clin. N. Am. Food Anim. Pract. 1997, 13, 425–454. [Google Scholar] [CrossRef]
- Riffault, S.; Meyer, G.; Deplanche, M.; Dubuquoy, C.; Durand, G.; Soulestin, M.; Castagné, N.; Bernard, J.; Bernardet, P.; Dubosclard, V.; et al. A new subunit vaccine based on nucleoprotein nanoparticles confers partial clinical and virological protection in calves against bovine respiratory syncytial virus. Vaccine 2010, 28, 3722–3734. [Google Scholar] [CrossRef] [Green Version]
- Lhermie, G.; Ferran, A.A.; Assié, S.; Cassard, H.; El Garch, F.; Schneider, M.; Woerhlé, F.; Pacalin, D.; Delverdier, M.; Bousquet-Mélou, A.; et al. Impact of timing and dosage of a fluoroquinolone treatment on the microbiological, pathological, and clinical outcomes of calves challenged with Mannheimia haemolytica. Front. Microbiol. 2016, 7, 237. [Google Scholar] [CrossRef] [Green Version]
- Salem, E.; Hägglund, S.; Cassard, H.; Corre, T.; Näslund, K.; Foret, C.; Gauthier, D.; Pinard, A.; Delverdier, M.; Zohari, S.; et al. Pathogenesis, host innate immune response, and aerosol transmission of Influenza D virus in cattle. J. Virol. 2019, 93, e01853-18. [Google Scholar] [CrossRef] [Green Version]
- Makoschey, B.; Ramage, C.; Reddick, D.; Fraser, S.; Donachie, W. Colostrum from cattle immunized with a vaccine based on iron regulated proteins of Mannheimia haemolytica confers partial protection. Vaccine 2012, 30, 969–973. [Google Scholar] [CrossRef]
- Tian, D.; Battles, M.B.; Moin, S.M.; Chen, M.; Modjarrad, K.; Kumar, A.; Kanekiyo, M.; Graepel, K.W.; Taher, N.M.; Hotard, A.L.; et al. Structural basis of respiratory syncytial virus subtype-dependent neutralization by an antibody targeting the fusion glycoprotein. Nat. Commun. 2017, 8, 1877. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McLellan, J.S. Neutralizing epitopes on the respiratory syncytial virus fusion glycoprotein. Curr. Opin. Virol. 2015, 11, 70–75. [Google Scholar] [CrossRef] [Green Version]
- Goodwin, E.; Gilman, M.S.A.; Wrapp, D.; Chen, M.; Ngwuta, J.O.; Moin, S.M.; Bai, P.; Sivasubramanian, A.; Connor, R.I.; Wright, P.F.; et al. Infants infected with respiratory syncytial virus generate potent neutralizing antibodies that lack somatic hypermutation. Immunity 2018, 48, P339–P349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koivisto, K.; Nieminen, T.; Mejias, A.; Gonzalez, C.C.; Ye, F.; Mertz, S.; Peeples, M.; Ramilo, O.; Saxén, H. Respiratory Syncytial Virus (RSV)–Specific Antibodies in Pregnant Women and Subsequent Risk of RSV Hospitalization in Young Infants. J. Infect. Dis. 2022, 225, 1189–1196. [Google Scholar] [CrossRef]
- Stensballe, L.G.; Ravn, H.; Kristensen, K.; Meakins, T.; Aaby, P.; Simoes, E.A.F. Seasonal Variation of Maternally Derived Respiratory Syncytial Virus Antibodies and Association with Infant Hospitalizations for Respiratory Syncytial Virus. J. Pediatr. 2009, 154, P296–P298. [Google Scholar] [CrossRef]
- Marchant, A.; Sadarangani, M.; Garand, M.; Dauby, N.; Verhasselt, V.; Pereira, L.; Bjornson, G.; Joners, C.E.; Halperin, S.A.; Edwards, K.L.; et al. Maternal immunisation: Collaborating with mother nature. Lancet Infect. Dis. 2017, 17, E197–E208. [Google Scholar] [CrossRef]
- Heath, P.T.; Culley, F.J.; Jones, C.E.; Kampmann, B.; Le Doare, K.; Nunes, M.C.; Sadarangani, M.; Chaudhry, Z.; Baker, C.J.; Openshaw, P.J.M. Group B streptococcus and respiratory syncytial virus immunisation during pregnancy: A landscape analysis. Lancet Infect. Dis. 2017, 17, E223–E234. [Google Scholar] [CrossRef]
- Dodd, J.S.; Lum, E.; Goulding, J.; Muir, R.; Van Snick, J.; Openshaw, P.J.M. IL-9 regulates pathology during primary and memory responses to respiratory syncytial virus infection. J. Immunol. 2009, 183, 7006–7013. [Google Scholar] [CrossRef] [Green Version]
- McNamara, P.S.; Flanagan, B.J.; Baldwin, L.M.; Newland, P.; Hart, C.A.; Smyth, R.L. Interleukin 9 production in the lungs of infants with severe respiratory syncytial virus bronchiolitis. Lancet 2004, 363, 1031–1037. [Google Scholar] [CrossRef] [PubMed]
- Semple, M.G.; Dankert, H.M.; Ebrahimi, B.; Correia, J.B.; Booth, J.A.; Stewart, J.P.; Smyth, R.L.; Hart, C.A. Severe respiratory syncytial virus bronchiolitis in infants is associated with reduced airway interferon gamma and substance P. PLoS ONE 2007, 2, e1038. [Google Scholar] [CrossRef] [PubMed]
- Hoebee, B.; Bont, L.; Rietveld, E.; van Oosten, M.; Hodemaekers, H.M.; Nagelkerke, N.J.D.; Neijens, H.J.; Kimpen, J.L.L.; Kimman, T.G. Influence of promoter variants of interleukin-10, interleukin-9, and tumor necrosis factor-alpha genes on respiratory syncytial virus bronchiolitis. J. Infect. Dis. 2004, 189, 239–247. [Google Scholar] [CrossRef] [PubMed]
- Sebina, I.; Phipps, S. The Contribution of Neutrophils to the Pathogenesis of RSV Bronchiolitis. Viruses 2020, 12, 808. [Google Scholar] [CrossRef]
- Tahamtan, A.; Besteman, S.; Samadizadeh, S.; Rastegar, M.; Bont, L.; Salimi, V. Neutrophils in respiratory syncytial virus infection: From harmful effects to therapeutic opportunities. Br. J. Pharmacol. 2021, 178, 515–530. [Google Scholar] [CrossRef]
- Fulton, R.W.; Briggs, R.E.; Payton, M.E.; Confer, A.W.; Saliki, J.T.; Ridpath, J.F.; Burge, L.J.; Duff, G.C. Maternally derived humoral immunity to bovine viral diarrhea virus (BVDV) 1a, BVDV1b, BVDV2, bovine herpesvirus-1, parainfluenza-3 virus bovine respiratory syncytial virus, Mannheimia haemolytica and Pasteurella multocida in beef calves, antibody decline by half-life studies and effect on response to vaccination. Vaccine 2004, 22, 643–649. [Google Scholar] [PubMed]
- Martin, S.W.; Nagy, E.; Armstrong, D.; Rosendal, S. The associations of viral and Mycoplasmal antibody titers with respiratory disease and weight gain in feedlot calves. Can. Vet. J. 1999, 40, 560–567. [Google Scholar]
- Moerman, A.; Straver, P.J.; de Jong, M.C.; Quak, J.; Baanvinger, T.; van Oirschot, J.T. Clinical consequences of a bovine virus diarrhea virus infection in a dairy herd: A longitudinal study. Vet. Q. 1999, 16, 115–119. [Google Scholar] [CrossRef]
- Chamorro, M.F.; Walz, P.H.; Haines, D.M.; Passler, T.; Earleywine, T.; Palomares, R.A.; Riddell, K.P.; Galik, P.; Zhang, Y.; Givens, M.D. Comparison of levels and duration of detection of antibodies to bovineviral diarrhea virus 1, bovine viral diarrhea virus 2, bovine respiratory syncytial virus, bovine herpesvirus 1, and bovine parainfluenza virus 3 in calves fed maternal colostrum or a colostrum-replacement product. Can. J. Vet. Res. 2014, 78, 81–88. [Google Scholar] [PubMed]
- Ferellaa, A.; Aguirreburualdeb, M.S.P.; Sammarrucoa, A.; Parreñov, V.; Santos, M.J.D.; Mozgovoj, M. Dynamics of neutralizing antibodies against Bovine respiratory syncytial virus in a dairy herd from Santa Fe Province, Argentina. Rev. Argent. Microbiol. 2020, 52, 293–297. [Google Scholar] [CrossRef] [PubMed]
- Kirkpatrick, J.; Fulton, R.W.; Burge, L.J.; Dubois, W.R.; Payton, M. Passively transferred immunity in newborn calves, rate of antibody decay, and effect on subsequent vaccination with modified live virus vaccine. Bov. Pract. 2001, 35, 47–55. [Google Scholar]
- Munoz-Zanzi, C.; Thurmond, M.; Johnson, W.O.; Hietala, S.K. Predicted ages of dairy calves when colostrum derived bovine viral diarrhea virus antibodies would no longer offer protection against disease or interfere with vaccination. J. Am. Vet. Med. Assoc. 2002, 221, 678–685. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Date | ELISA (Number of Animals) | Neutralization (Mean Titers with Range in Square Brackets; Log2 EID50/mL) | |||
|---|---|---|---|---|---|
| Non-Vaccinated | Vaccinated | Non-Vaccinated | Vaccinated | ||
| Cows (n = 16/group) | Before vaccination | Neg (6) + (6) ++ (4) | Neg (6) + (6) ++ (4) | 2.1 [0–3.4] | 2.4 [0–3.9] |
| Two weeks after vaccination | Neg (6) + (6) ++ (4) | ++ (1) +++ (2) ++++ (13) | 2.3 [0–3.4] | 8.6 [7–9.2] | |
| Colostrum (n = 16/group) | Neg (12) + (4) | ++ (4) +++ (4) ++++ (8) | 1.8 [0–7.6] | 9.4 [6.1–11.7] | |
| Calves (ELISA, n = 14/group) (SN, n = 7/group) | D-19 | Neg (6) + (4) ++ (4) | ++ (4) +++ (5) ++++ (5) | 0.9 [0.2–3.8] | 8.8 [7.2–9.1] |
| D0 | Neg (6) + (4) ++ (4) | ++ (4) +++ (5) ++++ (5) | 0.6 [0–3.8] | 8.4 [7.1–9] | |
| D16 | ++ (2) +++ (6) ++++ (6) | ++ (3) +++ (6) ++++ (5) | 8.7 [7.5–10.5] | 8.8 [8.3–9.3] | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Meyer, G.; Foret-Lucas, C.; Delverdier, M.; Cuquemelle, A.; Secula, A.; Cassard, H. Protection against Bovine Respiratory Syncytial Virus Afforded by Maternal Antibodies from Cows Immunized with an Inactivated Vaccine. Vaccines 2023, 11, 141. https://doi.org/10.3390/vaccines11010141
Meyer G, Foret-Lucas C, Delverdier M, Cuquemelle A, Secula A, Cassard H. Protection against Bovine Respiratory Syncytial Virus Afforded by Maternal Antibodies from Cows Immunized with an Inactivated Vaccine. Vaccines. 2023; 11(1):141. https://doi.org/10.3390/vaccines11010141
Chicago/Turabian StyleMeyer, Gilles, Charlotte Foret-Lucas, Maxence Delverdier, Antoine Cuquemelle, Aurélie Secula, and Hervé Cassard. 2023. "Protection against Bovine Respiratory Syncytial Virus Afforded by Maternal Antibodies from Cows Immunized with an Inactivated Vaccine" Vaccines 11, no. 1: 141. https://doi.org/10.3390/vaccines11010141
APA StyleMeyer, G., Foret-Lucas, C., Delverdier, M., Cuquemelle, A., Secula, A., & Cassard, H. (2023). Protection against Bovine Respiratory Syncytial Virus Afforded by Maternal Antibodies from Cows Immunized with an Inactivated Vaccine. Vaccines, 11(1), 141. https://doi.org/10.3390/vaccines11010141