
Recombinant Porcine Interferon-α Decreases Pseudorabies Virus Infection


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. DNA Construct of Porcine Interferon-α
2.3. Preparation of Recombinant Lentivirus HIV-pCDH-PoIFN-α
2.4. Generation of Stable Expression CHO-K1
2.4.1. Identification of Monoclonal Cells Using Western Blot, Immunofluorescence (IF), and Quantitative Reverse-Transcription Polymerase Chain Reaction (qRT-PCR)
2.4.2. ELISA Quantification of Recombinant Protein (HIV-pCDH-PoIFN-α)
2.5. Identification of Toxicity of Recombinant Protein
2.6. In Vivo Experiment
2.6.1. Detection of Viral Load of PVR by qPCR
2.6.2. Hematoxylin and Eosin Stain of Paraffin Sections
3. Results
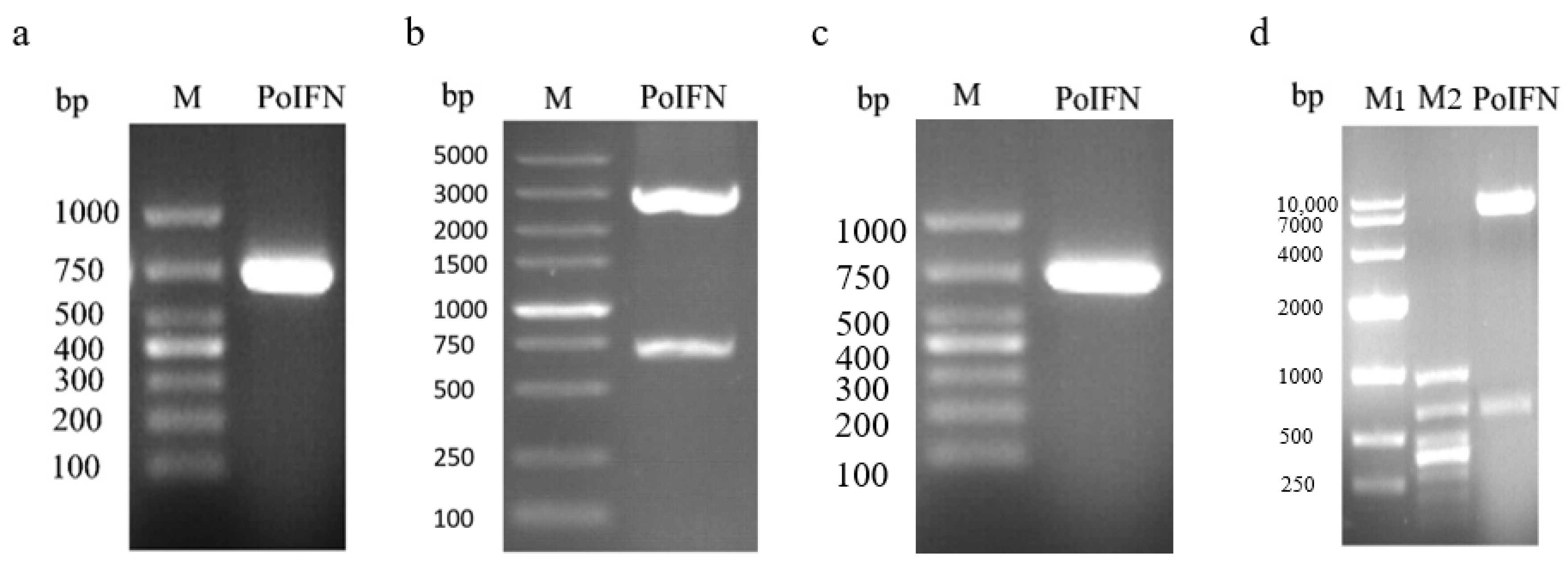
3.1. Plasmid Identification
3.2. Expression and Characterization of Recombinant Proteins
3.3. The Concentration of Recombinant Protein PoIFNα in Positive Cells
3.4. Identification of Toxicity of Recombinant Protein
3.5. Calculation of TCID50 and TD50
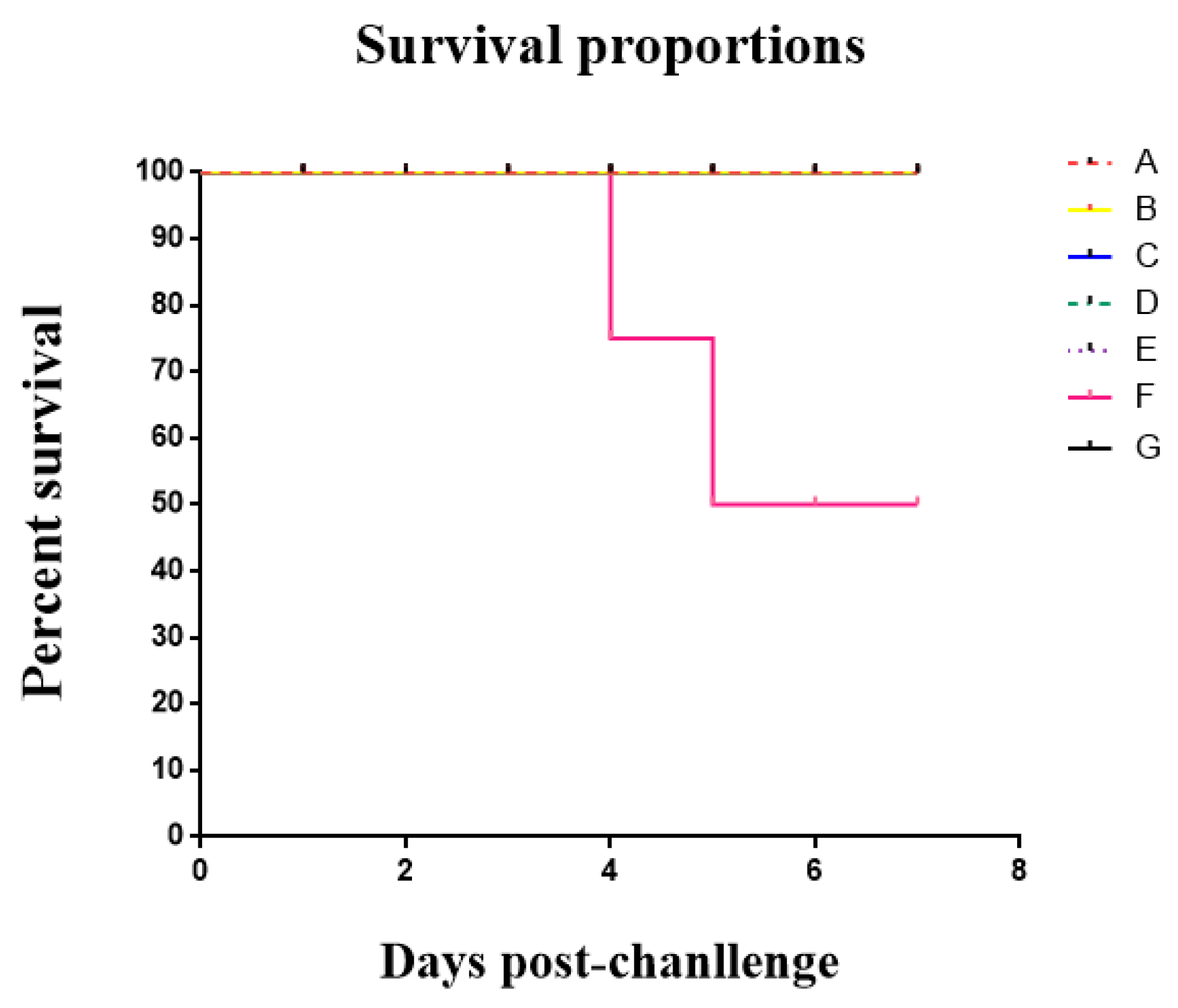
3.6. The Result of the Clinical Trial
3.6.1. Viral Load in Different Tissues of Mice
3.6.2. Histopathological Sections of Mouse Tissues
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nagano, Y.; Kojima, Y. Inhibition of vaccinia infection by a liquid factor in tissues infected by homologous virus. C. R. Séances Société Biol. Ses Fil. 1958, 152, 1627–1629. [Google Scholar]
- Belardelli, F. Role of interferons and other cytokines in the regulation of the immune response. APMIS Acta Pathol. Microbiol. Immunol. Scand. 1995, 103, 161–179. [Google Scholar] [CrossRef]
- Samuel, C.E. Antiviral actions of interferon. Interferon-regulated cellular proteins and their surprisingly selective antiviral activities. Virology 1991, 183, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Finter, N.B. The naming of cats—And alpha-interferons. Lancet 1996, 348, 348–349. [Google Scholar] [CrossRef] [PubMed]
- Holding, M.L.; Frazier, J.A.; Dorr, S.W.; Pollock, N.B.; Muelleman, P.J.; Branske, A.; Henningsen, S.N.; Eikenaar, C.; Escallón, C.; Montgomery, C.E.; et al. Wet- and dry-season steroid hormone profiles and stress reactivity of an insular dwarf snake, the Hog Island boa (Boa constrictor imperator). Physiol. Biochem. Zool. 2014, 87, 363–373. [Google Scholar] [CrossRef]
- Zhou, A.; Paranjape, J.M.; Der, S.D.; Williams, B.R.; Silverman, R.H. Interferon action in triply deficient mice reveals the existence of alternative antiviral pathways. Virology 1999, 258, 435–440. [Google Scholar] [CrossRef] [PubMed]
- Finke, D.; Eloranta, M.L.; Ronnblom, L. Endogenous type I interferon inducers in autoimmune diseases. Autoimmunity 2009, 42, 349–352. [Google Scholar] [CrossRef] [PubMed]
- Fu, X.Y.; Kessler, D.S.; Veals, S.A.; Levy, D.E.; Darnell, J.E., Jr. ISGF3, the transcriptional activator induced by interferon alpha, consists of multiple interacting polypeptide chains. Proc. Natl. Acad. Sci. USA 1990, 87, 8555–8559. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Yang, W.; Wang, X.; Zhang, X.; Tian, H.; Deng, H.; Zhang, L.; Gao, G. Identification of new type I interferon-stimulated genes and investigation of their involvement in IFN-beta activation. Protein Cell 2018, 9, 799–807. [Google Scholar] [CrossRef]
- Levy, D.E.; Kessler, D.S.; Pine, R.; Reich, N.; Darnell, J.E. Interferon-induced nuclear factors that bind a shared promoter element correlate with positive and negative transcriptional control. Genes Dev. 1988, 2, 383–393. [Google Scholar] [CrossRef] [PubMed]
- Fink, K.; Grandvaux, N. STAT2 and IRF9: Beyond ISGF3. Jak-Stat 2013, 2, e27521. [Google Scholar] [CrossRef]
- Arshanapalli, A.; Shah, M.; Veerula, V.; Somani, A.K. The role of type I interferons and other cytokines in dermatomyositis. Cytokine 2015, 73, 319–325. [Google Scholar] [CrossRef] [PubMed]
- Miermont, A.; Uhlendorf, J.; McClean, M.; Hersen, P. The Dynamical systems properties of the HOG signaling cascade. J. Recept. Signal Transduct. 2011, 2011, 930940. [Google Scholar] [CrossRef]
- Brown, F. The classification and nomenclature of viruses: Summary of results of meetings of the International Committee on Taxonomy of Viruses in Edmonton, Canada 1987. Intervirology 1989, 30, 181–186. [Google Scholar] [CrossRef] [PubMed]
- Roizman, B.B.N.; Knipe, D.M.; Howley, P.M. Herpesviridae. In Fields Virology, 3rd ed.; Fields, Ed.; Lippencott-Raven: Philadelphia, PA, USA, 1996; pp. 2221–2230. [Google Scholar]
- Roizmann, B.; Desrosiers, R.C.; Fleckenstein, B.; Lopez, C.; Minson, A.C.; Studdert, M.J. The family Herpesviridae: An update. The Herpesvirus Study Group of the International Committee on Taxonomy of Viruses. Arch. Virol. 1992, 123, 425–449. [Google Scholar] [CrossRef] [PubMed]
- Zhao, P.; Wang, Y.; Zhang, P.; Du, F.; Li, J.; Wang, C.; Fang, R.; Zhao, J. Epidemiological Investigation, Risk Factors, Spatial-Temporal Cluster, and Epidemic Trend Analysis of Pseudorabies Virus Seroprevalence in China (2017 to 2021). Microbiol. Spectr. 2023, 11, e0529722. [Google Scholar] [CrossRef] [PubMed]
- Verin, R.; Varuzza, P.; Mazzei, M.; Poli, A. Serologic, molecular, and pathologic survey of pseudorabies virus infection in hunted wild boars (Sus scrofa) in Italy. J. Wildl. Dis. 2014, 50, 559–565. [Google Scholar] [CrossRef]
- Pensaert, M.; Kluge, J. Pseudorabies virus (Aujeszky’s disease). Virus Infect. Porc. 1989, 2, 39–65. [Google Scholar]
- Glass, C.M.; McLean, R.G.; Katz, J.B.; Maehr, D.S.; Cropp, C.B.; Kirk, L.J.; McKeirnan, A.J.; Evermann, J.F. Isolation of pseudorabies (Aujeszky’s disease) virus from a Florida panther. J. Wildl. Dis. 1994, 30, 180–184. [Google Scholar] [CrossRef] [PubMed]
- Baskerville, A.; McFerran, J.B.; Dow, C. Aujeszky’s disease in pigs. Vet. Bull. 1973, 43, 465–480. [Google Scholar]
- Mulder, W.A.; Pol, J.M.; Gruys, E.; Jacobs, L.; De Jong, M.C.M.; Peeters, B.P.H.; Kimman, T.G. Pseudorabies virus infections in pigs. Role of viral proteins in virulence, pathogenesis and transmission. Vet. Res. 1997, 28, 1–17. [Google Scholar]
- Iglesias, G.J.; Trujano, M.; Lokensgard, J.; Molitor, T. Study of the potential involvement of pseudorabies virus in swine respiratory disease. Can. J. Vet. Res. 1992, 56, 74–77. [Google Scholar]
- Peeters, B.; de Wind, N.; Broer, R.; Gielkens, A.; Moormann, R. Glycoprotein H of pseudorabies virus is essential for entry and cell-to-cell spread of the virus. J. Virol. 1992, 66, 3888–3892. [Google Scholar] [CrossRef]
- Engel, M.; Wierup, M. Eradication of Aujeszky’s disease virus from a swedish pig herd using gI-/TK-vaccine. Vet. Rec. 1997, 140, 493–495. [Google Scholar] [CrossRef] [PubMed]
- Zheng, H.H.; Fu, P.F.; Chen, H.Y.; Wang, Z.Y. Pseudorabies Virus: From Pathogenesis to Prevention Strategies. Viruses 2022, 14, 1638. [Google Scholar] [CrossRef] [PubMed]
- Ye, N.; Feng, W.; Fu, T.; Tang, D.; Zeng, Z.; Wang, B. Membrane fusion, potential threats, and natural antiviral drugs of pseudorabies virus. Vet. Res. 2023, 54, 39. [Google Scholar] [CrossRef]
- Teijaro, J.R. Type I interferons in viral control and immune regulation. Curr. Opin. Virol. 2016, 16, 31–40. [Google Scholar] [CrossRef] [PubMed]
- Szulc, B.; Piasecki, E. Effects of interferons, interferon inducers and growth factors on phagocytosis measured by quantitative determination of synthetic compound ingested by mouse bone marrow-derived macrophages. Arch. Immunol. Ther. Exp. 1988, 36, 537–545. [Google Scholar]
- Hance, K.W.; Rogers, C.J.; Zaharoff, D.A.; Canter, D.; Schlom, J.; Greiner, J.W. The antitumor and immunoadjuvant effects of IFN-alpha in combination with recombinant poxvirus vaccines. Clin. Cancer Res. 2009, 15, 2387–2396. [Google Scholar] [CrossRef]
- Li, L.; Sherry, B. IFN-alpha expression and antiviral effects are subtype and cell type specific in the cardiac response to viral infection. Virology 2010, 396, 59–68. [Google Scholar] [CrossRef]
- Sun, M.; Hao, F.; Li, W.; Cheng, Z.; Zhang, W.; Yang, L.; Mao, L.; Liu, M. Temporal Dynamics of the Ruminant Type I IFN-Induced Antiviral State against Homologous Parainfluenza Virus 3 Challenge In Vitro. Viruses 2022, 14, 1025. [Google Scholar] [CrossRef] [PubMed]
- Matos, A.D.R.; Wunderlich, K.; Schloer, S.; Schughart, K.; Geffers, R.; Seders, M.; Witt, M.; Christersson, A.; Wiewrodt, R.; Wiebe, K.; et al. Antiviral potential of human IFN-α subtypes against influenza A H3N2 infection in human lung explants reveals subtype-specific activities. Emerg. Microbes Infect. 2019, 8, 1763–1776. [Google Scholar] [CrossRef] [PubMed]
- Quintana, M.E.; Barone, L.J.; Trotta, M.V.; Turco, C.; Mansilla, F.C.; Capozzo, A.V.; Cardoso, N.P. In-vivo Activity of IFN-λ and IFN-α Against Bovine-Viral-Diarrhea Virus in a Mouse Model. Front. Vet. Sci. 2020, 7, 45. [Google Scholar] [CrossRef] [PubMed]
- Porowińska, D.; Wujak, M.; Roszek, K.; Komoszyński, M. Prokariotyczne systemy ekspresyjne [Prokaryotic expression systems]. Adv. Hyg. Exp. Med. 2013, 67, 119–129. (In Polish) [Google Scholar]
- Baghban, R.; Farajnia, S.; Rajabibazl, M.; Ghasemi, Y.; Mafi, A.; Hoseinpoor, R.; Rahbarnia, L.; Aria, M. Yeast Expression Systems: Overview and Recent Advances. Mol. Biotechnol. 2019, 61, 365–384. [Google Scholar] [CrossRef]
- Murphy, C.I.; Piwnica-Worms, H. Overview of the baculovirus expression system. Curr. Protoc. Neurosci. 2001, 10, 4–18. [Google Scholar]
- Zhu, J.; Hatton, D. New Mammalian Expression Systems. In New Bioprocessing Strategies: Development and Manufacturing of Recombinant Antibodies and Proteins; Advances in Biochemical Engineering/Biotechnology; Kiss, B., Gottschalk, U., Pohlscheidt, M., Eds.; Springer: Cham, Switzerland, 2017; Volume 165. [Google Scholar]
- Mark, J.K.K.; Lim, C.S.Y.; Nordin, F.; Tye, G.J. Expression of mammalian proteins for diagnostics and therapeutics: A review. Mol. Biol. Rep. 2022, 49, 10593–10608. [Google Scholar] [CrossRef] [PubMed]
- Fischer, S.; Handrick, R.; Otte, K. The art of CHO cell engineering: A comprehensive retrospect and future perspectives. Biotechnol. Adv. 2015, 33, 1878–1896. [Google Scholar] [CrossRef] [PubMed]
- Segura, M.M.; Garnier, A.; Kamen, A. Purification and characterization of retrovirus vector particles by rate zonal ultracentrifugation. J. Virol. Methods 2006, 133, 82–91. [Google Scholar] [CrossRef]
- Gonçalves, M.A.; Janssen, J.M.; Holkers, M.; de Vries, A.A. Rapid and sensitive lentivirus vector-based conditional gene expression assay to monitor and quantify cell fusion activity. PLoS ONE 2010, 5, e10954. [Google Scholar] [CrossRef] [PubMed]
- Seidlits, S.K.; Gower, R.M.; Shepard, J.A.; Shea, L.D. Hydrogels for lentiviral gene delivery. Expert Opin. Drug Deliv. 2013, 10, 499–509. [Google Scholar] [CrossRef]
- Naldini, L.; Blömer, U.; Gallay, P.; Ory, D.; Mulligan, R.; Gage, F.H.; Verma, I.M.; Trono, D. In vivo gene delivery and stable transduction of nondividing cells by a lentiviral vector. Science 1996, 272, 263–267. [Google Scholar] [CrossRef]
- Mochizuki, H.; Schwartz, J.P.; Tanaka, K.; Brady, R.O.; Reiser, J. High-titer human immunodeficiency virus type 1-based vector systems for gene delivery into nondividing cells. J. Virol. 1998, 72, 8873–8883. [Google Scholar] [CrossRef] [PubMed]
- Pauwels, K.; Gijsbers, R.; Toelen, J.; Schambach, A.; Willard-Gallo, K.; Verheust, C.; Debyser, Z.; Herman, P. State-of-the-art lentiviral vectors for research use: Risk assessment and biosafety recommendations. Curr. Gene Ther. 2009, 9, 459–474. [Google Scholar] [CrossRef]
- Sena-Esteves, M.; Gao, G. Titration of Lentivirus Vectors. Cold Spring Harb. Protoc. 2018, 286. [Google Scholar] [CrossRef]
- Wang, G.S.; Du, Y.; Wu, J.Q.; Tian, F.L.; Yu, X.J.; Wang, J.B. Vaccine resistant pseudorabies virus causes mink infection in China. BMC Vet. Res. 2018, 14, 20. [Google Scholar] [CrossRef] [PubMed]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Song, B.; Wei, W.; Liu, X.; Huang, Y.; Zhu, S.; Yi, L.; Eerdunfu; Ding, H.; Zhao, M.; Chen, J. Recombinant Porcine Interferon-α Decreases Pseudorabies Virus Infection. Vaccines 2023, 11, 1587. https://doi.org/10.3390/vaccines11101587
Song B, Wei W, Liu X, Huang Y, Zhu S, Yi L, Eerdunfu, Ding H, Zhao M, Chen J. Recombinant Porcine Interferon-α Decreases Pseudorabies Virus Infection. Vaccines. 2023; 11(10):1587. https://doi.org/10.3390/vaccines11101587
Chicago/Turabian StyleSong, Bowen, Wenkang Wei, Xueyi Liu, Yaoyao Huang, Shuaiqi Zhu, Lin Yi, Eerdunfu, Hongxing Ding, Mingqiu Zhao, and Jinding Chen. 2023. "Recombinant Porcine Interferon-α Decreases Pseudorabies Virus Infection" Vaccines 11, no. 10: 1587. https://doi.org/10.3390/vaccines11101587
APA StyleSong, B., Wei, W., Liu, X., Huang, Y., Zhu, S., Yi, L., Eerdunfu, Ding, H., Zhao, M., & Chen, J. (2023). Recombinant Porcine Interferon-α Decreases Pseudorabies Virus Infection. Vaccines, 11(10), 1587. https://doi.org/10.3390/vaccines11101587