
CAR-T Cells with Phytohemagglutinin (PHA) Provide Anti-Cancer Capacity with Better Proliferation, Rejuvenated Effector Memory, and Reduced Exhausted T Cell Frequencies
 , , , ,
, , , ,  ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Synthesis of CAR Construct and Lentivirus Production
2.2. Lentivirus Titration
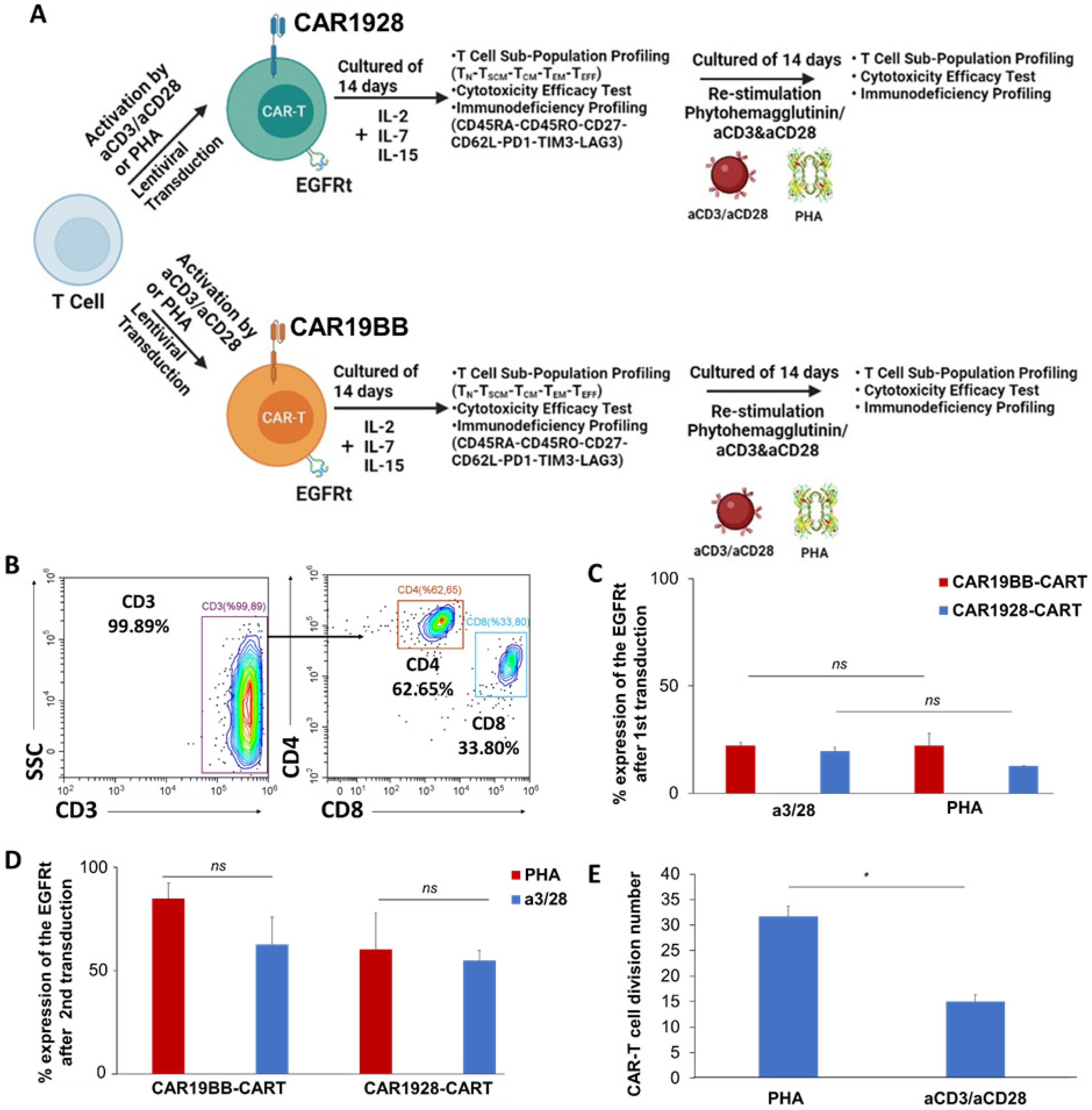
2.3. T Cell Transduction and CAR-T Culture Conditions
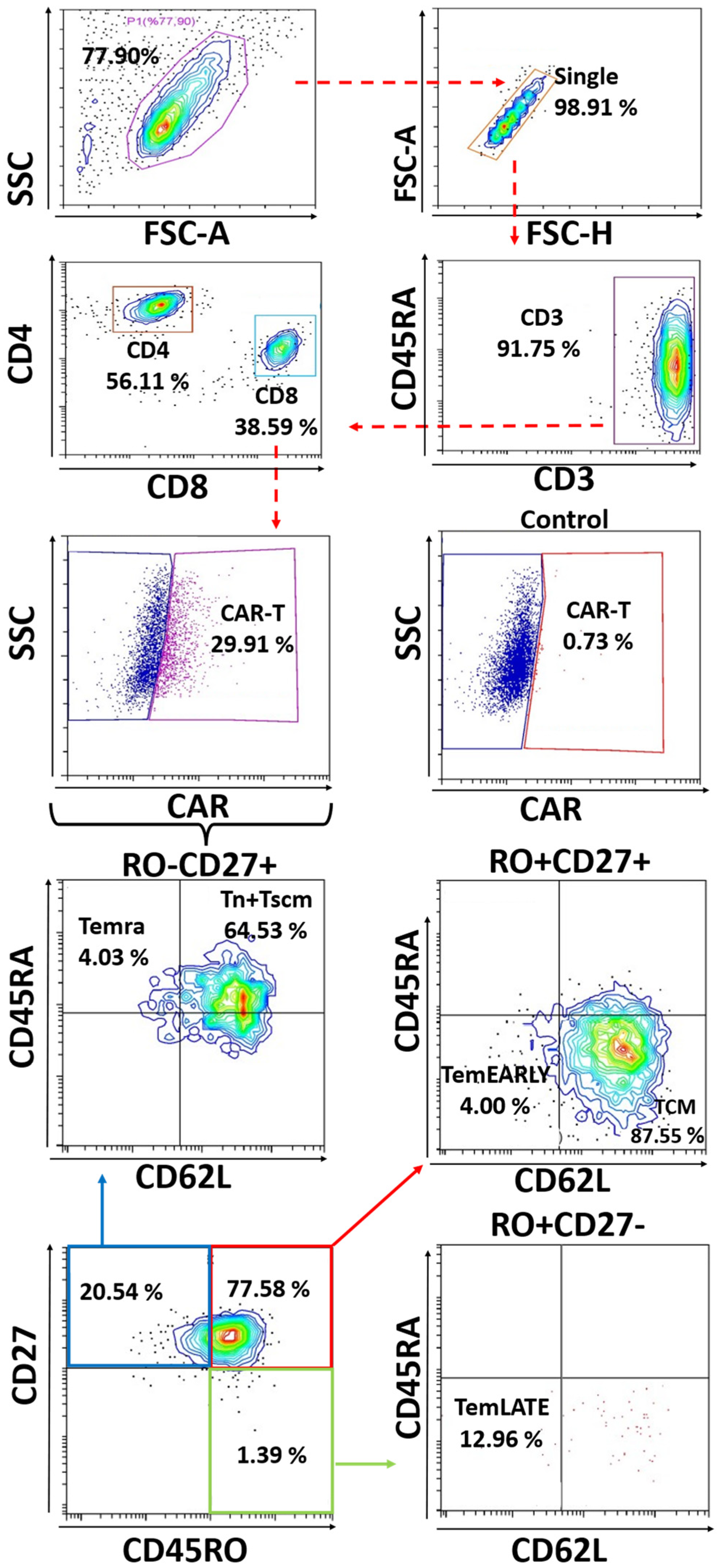
2.4. Analysis of T Cell Sub-Populations (Tn-Tcm-Tem-Tef) and Exhaustion Profile
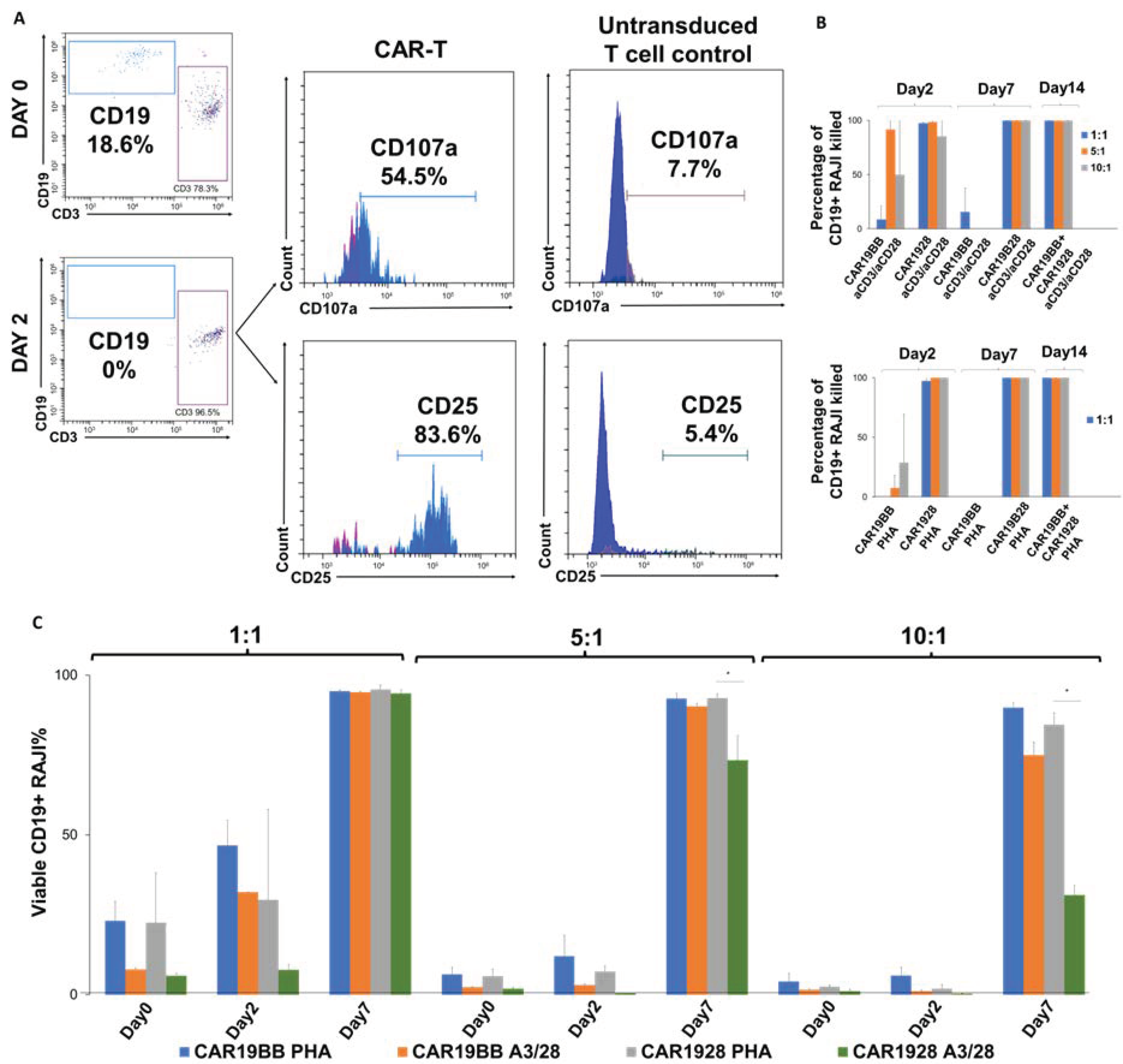
2.5. In Vitro Anti-Tumor Cytotoxicity and Efficacy Assay
2.6. Statistics Analysis
3. Results
3.1. In Vitro Assessment of CAR-T Cell Proliferation, Differentiation, and Anti-Cancer Capacity
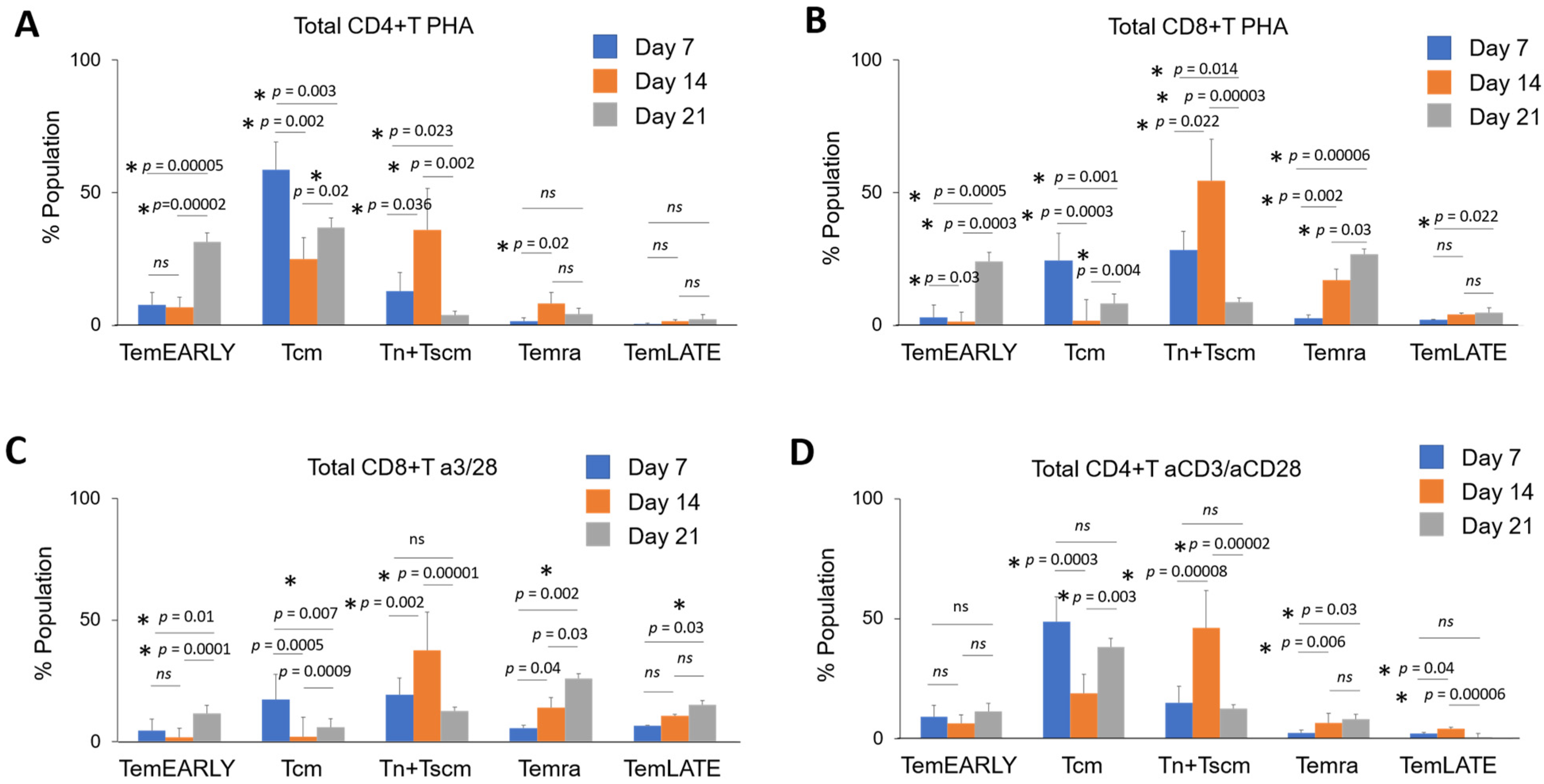
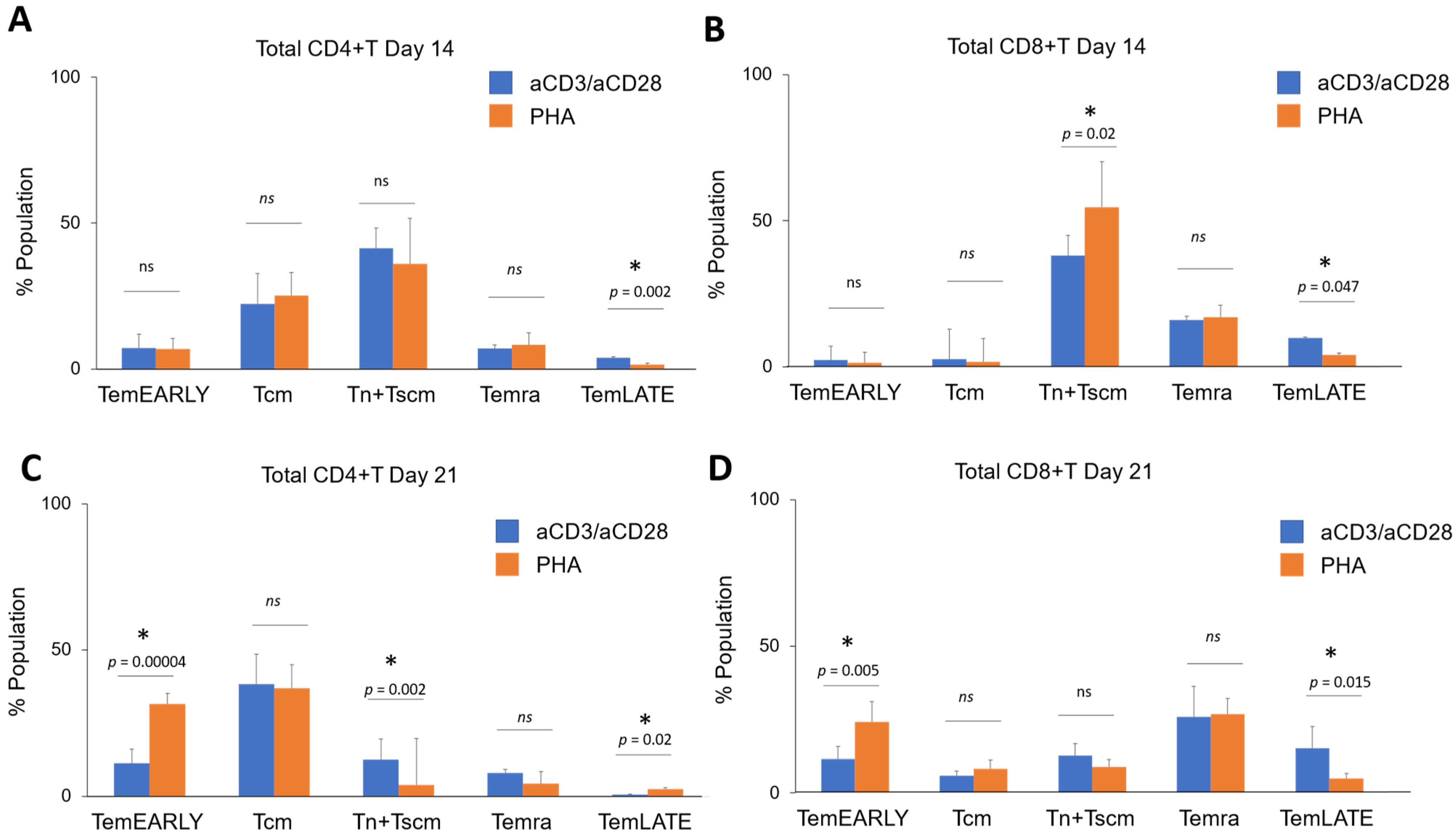
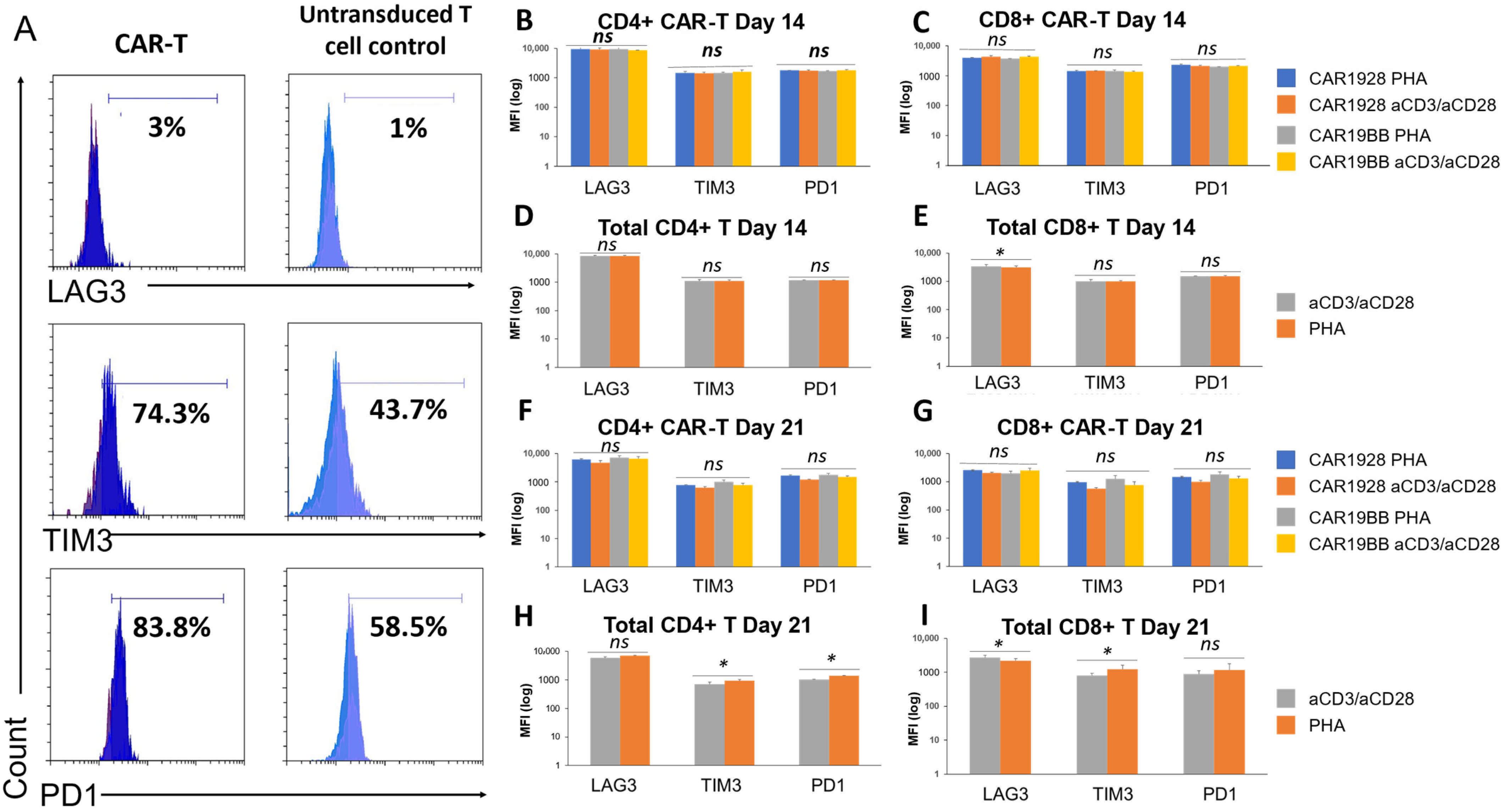
3.2. Immunoprofiling Sub-Populations of The Activated T Cell (Tn-Tcm-Tem-Tef)
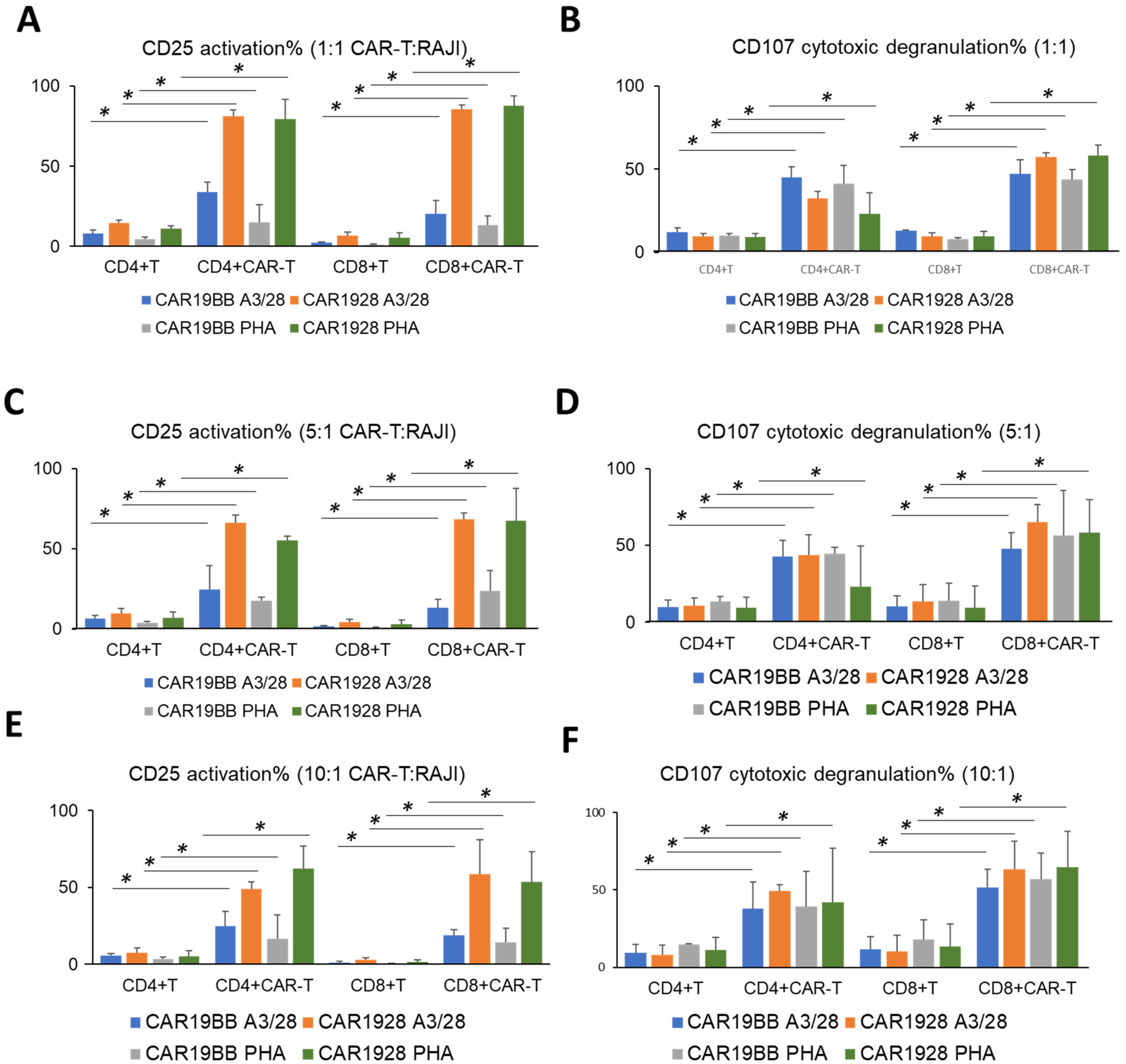
3.3. Cytotoxic Activity of PHA- or aCD3/aCD28-Activated CAR-T Cells
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Frey, A.B. Suppression of T cell responses in the tumor microenvironment. Vaccine 2015, 33, 7393–7400. [Google Scholar] [CrossRef]
- Finney, H.M.; Lawson, A.D.; Bebbington, C.R.; ve Weir, A.N. Chimeric receptors provide both primary and costimulatory signaling in T cells from a single gene product. J. Immunol. 1950, 161, 2791–2797. [Google Scholar] [CrossRef]
- Sadelain, M.; Brentjens, R.; Riviere, I. The basic principles of chimeric antigen receptor design. Cancer Discov. 2013, 3, 388–398. [Google Scholar] [CrossRef] [Green Version]
- Sadelain, M. CAR therapy: The CD19 paradigm. J. Clin. Investig. 2015, 125, 3392–3400. [Google Scholar] [CrossRef] [Green Version]
- Chang, Z.L.; Chen, Y.Y. CARs: Synthetic immunoreceptors for cancer therapy and beyond. Trends Mol. Med. 2017, 23, 430–450. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.; Liu, J.; Zhong, J.F.; Zhang, X. Engineering CAR-T cells. Biomark. Res. 2017, 5, 22. [Google Scholar] [CrossRef] [Green Version]
- Lee, D.W.; Kochenderfer, J.N.; Stetler-Stevenson, M.; Cui, Y.K.; Delbrook, C.; Feldman, S.A.; Fry, T.J.; Orentas, R.; Sabatino, M.; Shah, N.N.; et al. T cells expressing CD19 chimeric antigen receptors for acute lymphoblastic leukaemia in children and young adults: A phase 1 dose-escalation trial. Lancet 2015, 385, 517–528. [Google Scholar] [CrossRef]
- Maude, S.L.; Frey, N.; Shaw, P.A.; Aplenc, R.; Barrett, D.M.; Bunin, N.J.; Chew, A.; Gonzalez, V.E.; Zheng, Z.; Lacey, S.F.; et al. Chimeric antigen receptor T cells for sustained remissions in leukemia. N. Engl. J. Med. 2014, 371, 1507–1517. [Google Scholar] [CrossRef] [Green Version]
- Porter, D.L.; Hwang, W.-T.; Frey, N.V.; Lacey, S.F.; Shaw, P.A.; Loren, A.W.; Bagg, A.; Marcucci, K.T.; Shen, A.; Gonzalez, V.; et al. Chimeric antigen receptor T cells persist and induce sustained remissions in relapsed refractory chronic lymphocytic leukemia. Sci. Transl. Med. 2015, 7, 303ra139. [Google Scholar] [CrossRef] [Green Version]
- Long, A.H.; Haso, W.M.; Shern, J.F.; Wanhainen, K.M.; Murgai, M.; Ingaramo, M.; Smith, J.P.; Walker, A.J.; Kohler, M.E.; Venkateshwara, V.R.; et al. 4-1BB costimulation ameliorates T cell exhaustion induced by tonic signaling of chimeric antigen receptors. Nat. Med. 2015, 21, 581–590. [Google Scholar] [CrossRef]
- Sabbagh, L.; Pulle, G.; Liu, Y.; Tsitsikov, E.N.; Watts, T.H. ERK-dependent Bim modulation downstream of the 4-1BB-TRAF1 signaling axis is a critical mediator of CD8 T cell survival in vivo. J. Immunol. 2008, 180, 8093–8101. [Google Scholar] [CrossRef] [Green Version]
- Blaeschke, F.; Stenger, D.; Kaeuferle, T.; Willier, S.; Lotfi, R.; Kaiser, A.D.; Assenmacher, M.; Döring, M.; Feucht, J.; Feuchtinger, T. Induction of a central memory and stem cell memory phenotype in functionally active CD4+ and CD8+ CAR T cells produced in an automated good manufacturing practice system for the treatment of CD19+ acute lymphoblastic leukemia. Cancer Immunol. Immunother. 2018, 67, 1053–1066. [Google Scholar] [CrossRef]
- Blaeschke, F.; Kaeuferle, T.; Feucht, J.; Weber, D.; Lotfi, R.; Kaiser, A.; Assenmacher, M.; Doering, M.; Feuchtinger, T. Defined Central Memory and Stem Memory T Cell Phenotype of CD4 and CD8 CAR T Cells for the Treatment of CD19+ Acute Lymphoblastic Leukemia in an Automated Closed System. Blood 2016, 128, 4558. [Google Scholar] [CrossRef]
- Martinez-Forero, I.; Azpilikueta, A.; Bolaños-Mateo, E.; Nistal-Villan, E.; Palazon, A.; Teijeira, A.; Perez-Chacon, G.; Morales-Kastresana, A.; Murillo, O.; Jure-Kunkel, M.; et al. T Cell Costimulation with Anti-CD137 Monoclonal Antibodies Is Mediated by K63–Polyubiquitin-Dependent Signals from Endosomes. J. Immunol. 2013, 190, 6694–6706. [Google Scholar] [CrossRef] [Green Version]
- Soskic, B.; Qureshi, O.S.; Hou, T.; Sansom, D.M. A transendocytosis perspective on the CD28/CTLA-4 pathway. Adv. Immunol. 2014, 124, 95–136. [Google Scholar]
- Lever, M.; Maini, P.K.; Van Der Merwe, P.A.; Dushek, O. Phenotypic models of T cell activation. Nat. Rev. Immunol. 2014, 14, 619–629. [Google Scholar] [CrossRef] [Green Version]
- Pène, J.; Rahmoun, M.; Temmerman, S.; Yssel, H. Use of anti-CD3/CD28 mAb coupled magnetic beads permitting subsequent phenotypic analysis of activated human T cells by indirect immunofluorescence. J. Immunol. Methods 2003, 283, 59–66. [Google Scholar] [CrossRef]
- Faguet, G.B. Mechanisms of lymphocyte activation. Binding kinetics of phytohemagglutinin to human lymphocytes. J. Biol. Chem. 1977, 252, 2095–2100. [Google Scholar] [CrossRef]
- Movafagh, A.; Heydary, H.; Mortazavi-Tabatabaei, S.A.; Azargashb, E. The Significance Application of Indigenous Phytohemagglutinin (PHA) Mitogen on Metaphase and Cell Culture Procedure. Iran. J. Pharm. Res. IJPR 2011, 10, 895–903. [Google Scholar]
- Duarte, R.F.; E Chen, F.; Lowdell, M.W.; Potter, M.N.; Lamana, M.L.; Prentice, H.G.; A Madrigal, J. Functional impairment of human T-lymphocytes following PHAinduced expansion and retroviral transduction: Implications for gene therapy. Gene Ther. 2002, 9, 1359–1368. [Google Scholar] [CrossRef] [Green Version]
- Isabel Da Cunha, M.; Terra, M. Interleukin-15 and Phytohaemagglutinin: A Combination to Study Human CD8+ T Cell Differentiation In Vitro; CESPU: Porto, Portugal, 2012. [Google Scholar]
- Supimon, K.; Sangsuwannukul, T.; Sujjitjoon, J.; Phanthaphol, N.; Chieochansin, T.; Poungvarin, N.; Wongkham, S.; Junking, M.; Yenchitsomanus, P.-T. Anti-mucin 1 chimeric antigen receptor T cells for adoptive T cell therapy of cholangiocarcinoma. Sci. Rep. 2021, 11, 6276. [Google Scholar] [CrossRef]
- Kabelitz, D.; Janssen, O. Antigen-induced death of T-lymphocytes. Front. Biosci. A J. Virtual Libr. 1997, 2, d61–d77. [Google Scholar] [CrossRef]
- Bulfone-Paus, S.; Ungureanu, D.; Pohl, T.; Lindner, G.; Paus, R.; Rückert, R.; Krause, H.; Kunzendorf, U. Interleukin-15 protects from lethal apoptosis in vivo. Nat. Med. 1997, 3, 1124–1128. [Google Scholar] [CrossRef]
- Taştan, C.; Kançağı, D.D.; Turan, R.D.; Yurtsever, B.; Çakırsoy, D.; Abanuz, S.; Yılancı, M.; Seyis, U.; Özer, S.; Mert, S.; et al. Preclinical Assessment of Efficacy and Safety Analysis of CAR-T Cells (ISIKOK-19) Targeting CD19-Expressing B-Cells for the First Turkish Academic Clinical Trial with Relapsed/Refractory ALL and NHL Patients. Turk. J. Hematol. 2020, 37, 234. [Google Scholar] [CrossRef]
- Ammirati, E.; Cianflone, D.; Vecchio, V.; Banfi, M.; Vermi, A.C.; De Metrio, M.; Grigore, L.; Pellegatta, F.; Pirillo, A.; Garlaschelli, K. Effector memory T cells are associated with atherosclerosis in humans and animal models. J. Am. Heart Assoc. 2012, 1, e000125. [Google Scholar] [CrossRef] [Green Version]
- Larbi, A.; Fulop, T. From “truly naïve” to “exhausted senescent” T cells: When markers predict functionality. Cytom. Part A 2014, 85, 25–35. [Google Scholar] [CrossRef]
- Alvarez-Fernandez, C.; Escribà-Garcia, L.; Vidal, S.; Sierra, J.; Briones, J. A short CD3/CD28 costimulation combined with IL-21 enhance the generation of human memory stem T cells for adoptive immunotherapy. J. Transl. Med. 2016, 14, 214. [Google Scholar] [CrossRef] [Green Version]
- Tuluc, F.; Spitsin, S.; Tustin, N.B.; Murray, J.B.; Tustin, R., III; Schankel, L.A.; Wiznia, A.; Nachman, S.; Douglas, S.D. Decreased PD-1 expression on CD8 lymphocyte subsets and increase in CD8 Tscm cells in children with HIV receiving raltegravir. AIDS Res. Hum. Retrovir. 2017, 33, 133–142. [Google Scholar] [CrossRef] [Green Version]
- Devarajan, P.; Chen, Z. Autoimmune effector memory T cells: The bad and the good. Immunol. Res. 2013, 57, 12–22. [Google Scholar] [CrossRef]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gulden, G.; Sert, B.; Teymur, T.; Ay, Y.; Tiryaki, N.N.; Mishra, A.K.; Ovali, E.; Tarhan, N.; Tastan, C. CAR-T Cells with Phytohemagglutinin (PHA) Provide Anti-Cancer Capacity with Better Proliferation, Rejuvenated Effector Memory, and Reduced Exhausted T Cell Frequencies. Vaccines 2023, 11, 313. https://doi.org/10.3390/vaccines11020313
Gulden G, Sert B, Teymur T, Ay Y, Tiryaki NN, Mishra AK, Ovali E, Tarhan N, Tastan C. CAR-T Cells with Phytohemagglutinin (PHA) Provide Anti-Cancer Capacity with Better Proliferation, Rejuvenated Effector Memory, and Reduced Exhausted T Cell Frequencies. Vaccines. 2023; 11(2):313. https://doi.org/10.3390/vaccines11020313
Chicago/Turabian StyleGulden, Gamze, Berranur Sert, Tarik Teymur, Yasin Ay, Nulifer Neslihan Tiryaki, Abhinava K. Mishra, Ercument Ovali, Nevzat Tarhan, and Cihan Tastan. 2023. "CAR-T Cells with Phytohemagglutinin (PHA) Provide Anti-Cancer Capacity with Better Proliferation, Rejuvenated Effector Memory, and Reduced Exhausted T Cell Frequencies" Vaccines 11, no. 2: 313. https://doi.org/10.3390/vaccines11020313
APA StyleGulden, G., Sert, B., Teymur, T., Ay, Y., Tiryaki, N. N., Mishra, A. K., Ovali, E., Tarhan, N., & Tastan, C. (2023). CAR-T Cells with Phytohemagglutinin (PHA) Provide Anti-Cancer Capacity with Better Proliferation, Rejuvenated Effector Memory, and Reduced Exhausted T Cell Frequencies. Vaccines, 11(2), 313. https://doi.org/10.3390/vaccines11020313