
Therapeutic Potential of Luteolin on Cancer

Abstract
:1. Introduction
2. Structure, Physiochemical Properties, and the Natural Sources of Luteolin
3. Pharmacokinetics of Luteolin
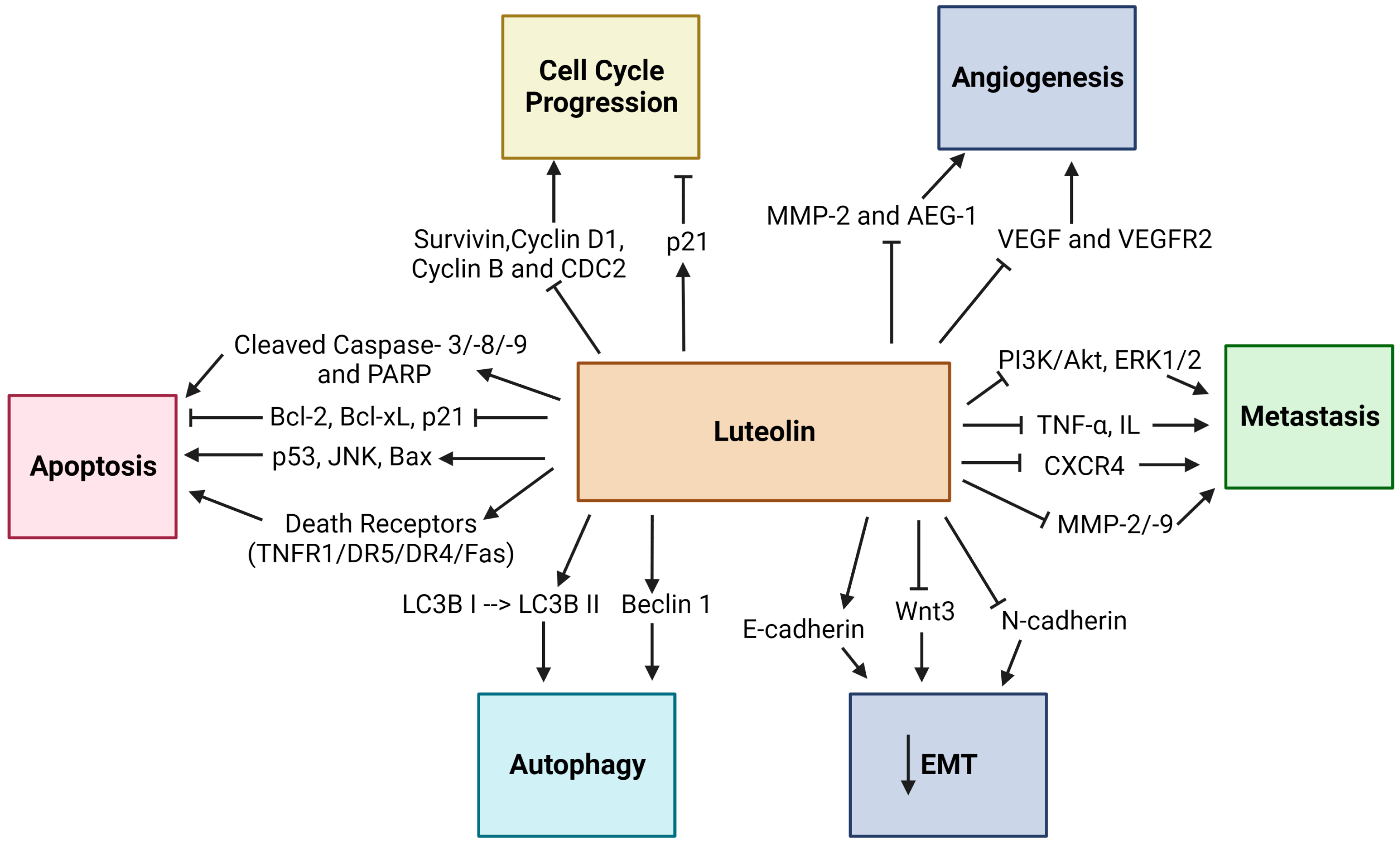
4. Anticancer Properties of Luteolin on Different Cancers
4.1. Luteolin and Colon Cancer
4.2. Luteolin and Lung Cancer
4.3. Luteolin and Prostate Cancer
4.4. Luteolin and Gastric Cancer
4.5. Luteolin and Glioblastoma
4.6. Luteolin and Liver Cancer
4.7. Luteolin and Breast Cancer

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cancer Type | Model System (Cell Lines or Animal Models) | Key Molecular Target(s) or Signaling Pathway(s) | Effects | Reference |
|---|---|---|---|---|
| Colon | Azoxymethane-induced mice | ↓ MMP-2/-9 | Inhibition of metastasis | [43] |
| HCT-15 cells | ↓ Wnt/β-Catenin/GSK-3β signaling cyclin D1 and Bcl-2 levels, ↑ Bax and Caspase-3 levels | Inhibition of proliferation, induction of apoptosis, and G2/M cell cycle arrest | [45] | |
| HT-29 and SNU-407 cells | ↑ Nrf2 levels and interaction between Nrf2 and p53 | Induction of apoptosis | [48] | |
| LoVo cells | ↓ CDC2 and Cyclin B levels ↑cytochrome c- and dATP-mediated activation of APAF-1 | G2/M cell cycle arrest Induction of apoptosis | [50] | |
| HT-29 cells | ↑ miR-384 levels | Inhibition of migration and invasion | [55] | |
| Lung | A549 cells | ↑ MEK/ERK signaling pathway | Inhibition of migration and induction of apoptosis | [57] |
| NCI-H460 cells | ↑ peIF2-α and CHOP levels Accumulation of LC3 II protein and ↑ LC3 puncta levels | Induction of ER stress-mediated apoptosis Induction of autophagy | [57] | |
| NCI-H1975 and NCI-H1650 cells Patient-derived xenograft mouse model | ↓ LIMK1 signaling pathway | Inhibition of proliferation and anchorage-independent cell growth Inhibition of tumor growth | [59] | |
| A549 cells | ↓ Src/FAK and its downstream Rac1, Cdc42, and RhoA pathways | Inhibition of invasion and metastasis | [71] | |
| Vascular endothelial cells of NSCLC | ↑ miR-133a-3p/PURB- mediated MAPK and PI3K/Akt pathways | Inhibition of migration and invasion | [74] | |
| H460 and A549 cells Mice xenograft models of lung cancer | ↓ circ_0000190 levels and ↑ miR-130a-3p (target of circ_0000190) | Inhibition of cell viability, migration, invasion, and colony formation and induction of apoptosis Inhibition of tumor growth | [75] | |
| Prostate | PC-3 cells Xenograft prostate tumor model | ↓ VEGFR-2-regulated AKT/ERK/mTOR/P70S6K/MMPs pathway | Inhibition of cell viability, migration, and invasion Inhibition of tumor growth and angiogenesis | [78] |
| PC-3 cells | Inhibition of Wnt signaling by ↑ FZD6 levels | Inhibition of prostate cancer stemness | [79] | |
| LNCaP cells | ↑ Prostate-derived Ets factor (PDEF) levels | Inhibition of proliferation and invasion | [80] | |
| PC3 and LNCaP cells | ↓ miR-301 levels | Inhibition of proliferation and induction of apoptosis | [85] | |
| Gastric | BGC-823 gastric carcinoma xenografts | ↓ VEGF-A and MMP-9 expressions | Inhibition of tumor growth | [87] |
| MKN45 and SGC7901 cells cMet-overexpressing Patient-derived human tumor xenograft models | ↓ cMet/Akt/ERK signaling | Inhibition of invasiveness and induction of apoptosis | [88] | |
| Hs-746T and MKN28 cells Mice xenograft model of gastric cancer | ↓ Notch1 signaling | Inhibition of proliferation and migration, and induction of apoptosis Inhibition of tumor growth and induction of apoptosis | [89] | |
| Hs-746T cells | ↓ Notch1-VEGF signaling | Inhibition of angiogenesis and vasculogenic mimicry formation | [90] | |
| BGC-823 and SGC-7901 cells | ↑ miR-34a levels | Induction of apoptosis | [92] | |
| Glioblastoma | U87MG and T98G cells | ↓ Cdc42 expression and PI3K/Akt activity | Inhibition of migration of glioblastoma cells | [101] |
| U251MG and U87MG cells | ↓ p-IGF-1R/PI3K/AKT/mTOR signaling pathway | Inhibition of migration of glioblastoma cells and reduction of the EMT process | [102] | |
| Liver | HepG2 cells | ↑ AMPK signaling pathway and ROS release | Induction of cell death | [106] |
| SK-Hep-1 cells | ↓ Akt/ osteopontin pathway | Induction of caspase-dependent apoptosis | [107] | |
| SMMC-7721 cells | Induction of LC3B-I conversion to LC3B-II, and ↑ Beclin 1 expression | Induction of apoptosis, partially via autophagy | [115] | |
| Breast | MCF-7 cells | ↓ EGFR signaling via mediation of PI3K/Akt, MAPK/Erk1/2 and STAT3 signaling pathways | Inhibition of cell proliferation induced by EGF | [118] |
| MDA-MB-231 cells | ↓ EGFR signaling | Induction of cell cycle arrest at the G2/M and S stages and apoptosis | [121] | |
| MCF-7 cells | Activation of Caspase-8 and Caspase-9 induced caspase-3 activity, ↑ Bax expression by ↓ Bcl-2 expression | Induction of apoptosis by activating the extrinsic and intrinsic pathways | [122] | |
| MDA-MB-231 cells | ↓ Human telomerase reverse transcriptase (hTERT) expression | Induction of cell cycle arrest at the S phase and apoptosis | [123] | |
| Tamoxifen-resistant MCF-7 cells | ↑ MLL3 expression | Induction of apoptosis through H3K4 monomethylation and suppression of the PI3K/AKT/mTOR pathway | [124] | |
| MDA-MB-231 cells | ↓ β-catenin expression | Inhibition of metastasis by reversing EMT | [128] | |
| MDA-MB-231 and 4T1 cells Mice xenograft model of breast cancer | ↓ YAP/TAZ activity | Inhibition of EMT and migration Inhibition of tumor growth | [129] | |
| BT-20 and MDA-MB-231 cells | ↓ MMP-9 expression through ↓AKT/mTOR-inducing H3K27Ac and H3K56Ac | Inhibition of proliferation and migration | [131] | |
| MCF-7 and MDA-MB-453 cells | ↑ miR-203 levels and ↓ Ras/Raf/MEK/ERK signaling pathway | Inhibition of breast cancer cell growth and EMT progress | [132] |
5. Nanodelivery Systems for Luteolin in Cancer Treatment
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ferlay, J.; Laversanne, M.; Ervik, M.; Lam, F.; Colombet, M.; Mery, L.; Piñeros, M.; Znaor, A.; Soerjomataram, I.; Bray, F. Global Cancer Observatory: Cancer Tomorrow. Available online: https://gco.iarc.fr/tomorrow/en (accessed on 3 December 2022).
- Wang, Y.; Chen, M.; Yu, H.; Yuan, G.; Luo, L.; Xu, X.; Xu, Y.; Sui, X.; Leung, E.L.-H.; Wu, Q. The Role and Mechanisms of Action of Natural Compounds in the Prevention and Treatment of Cancer and Cancer Metastasis. Front. Biosci. Landmark 2022, 27, 192. [Google Scholar] [CrossRef]
- Salehi, B.; Azzini, E.; Zucca, P.; Maria Varoni, E.; Kumar, N.V.A.; Dini, L.; Panzarini, E.; Rajkovic, J.; Valere Tsouh Fokou, P.; Peluso, I.; et al. Plant-Derived Bioactives and Oxidative Stress-Related Disorders: A Key Trend towards Healthy Aging and Longevity Promotion. Appl. Sci. 2020, 10, 947. [Google Scholar] [CrossRef] [Green Version]
- Sak, K. Anticancer Action of Plant Products: Changing Stereotyped Attitudes. Explor. Target Antitumor. Ther. 2022, 3, 423–427. [Google Scholar] [CrossRef] [PubMed]
- Slika, H.; Mansour, H.; Wehbe, N.; Nasser, S.A.; Iratni, R.; Nasrallah, G.; Shaito, A.; Ghaddar, T.; Kobeissy, F.; Eid, A.H. Therapeutic Potential of Flavonoids in Cancer: ROS-Mediated Mechanisms. Biomed. Pharmacother. 2022, 146, 112442. [Google Scholar] [CrossRef] [PubMed]
- Ullah, A.; Munir, S.; Badshah, S.L.; Khan, N.; Ghani, L.; Poulson, B.G.; Emwas, A.-H.; Jaremko, M. Important Flavonoids and Their Role as a Therapeutic Agent. Molecules 2020, 25, 5243. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Zheng, J.; Li, Y.; Xu, D.-P.; Li, S.; Chen, Y.-M.; Li, H.-B. Natural Polyphenols for Prevention and Treatment of Cancer. Nutrients 2016, 8, 515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Imran, M.; Rauf, A.; Abu-Izneid, T.; Nadeem, M.; Shariati, M.A.; Khan, I.A.; Imran, A.; Orhan, I.E.; Rizwan, M.; Atif, M.; et al. Luteolin, a Flavonoid, as an Anticancer Agent: A Review. Biomed. Pharmacother. 2019, 112, 108612. [Google Scholar] [CrossRef] [PubMed]
- Caporali, S.; de Stefano, A.; Calabrese, C.; Giovannelli, A.; Pieri, M.; Savini, I.; Tesauro, M.; Bernardini, S.; Minieri, M.; Terrinoni, A. Anti-Inflammatory and Active Biological Properties of the Plant-Derived Bioactive Compounds Luteolin and Luteolin 7-Glucoside. Nutrients 2022, 14, 1155. [Google Scholar] [CrossRef] [PubMed]
- Harborne, J.B.; Williams, C.A. Advances in Flavonoid Research since 1992. Phytochemistry 2000, 55, 481–504. [Google Scholar] [CrossRef]
- Muruganathan, N.; Dhanapal, A.R.; Baskar, V.; Muthuramalingam, P.; Selvaraj, D.; Aara, H.; Shiek Abdullah, M.Z.; Sivanesan, I. Recent Updates on Source, Biosynthesis, and Therapeutic Potential of Natural Flavonoid Luteolin: A Review. Metabolites 2022, 12, 1145. [Google Scholar] [CrossRef]
- Cai, Q.; Rahn, R.O.; Zhang, R. Dietary Flavonoids, Quercetin, Luteolin and Genistein, Reduce Oxidative DNA Damage and Lipid Peroxidation and Quench Free Radicals. Cancer Lett. 1997, 119, 99–107. [Google Scholar] [CrossRef] [PubMed]
- Choi, C.-W.; Jung, H.A.; Kang, S.S.; Choi, J.S. Antioxidant Constituents and a New Triterpenoid Glycoside from Flos Lonicerae. Arch. Pharm. Res. 2007, 30, 1–7. [Google Scholar] [CrossRef] [PubMed]
- López-Lázaro, M. Distribution and Biological Activities of the Flavonoid Luteolin. Mini. Rev. Med. Chem. 2009, 9, 31–59. [Google Scholar] [CrossRef]
- Lin, Y.; Shi, R.; Wang, X.; Shen, H.-M. Luteolin, a Flavonoid with Potential for Cancer Prevention and Therapy. Curr. Cancer Drug Targets 2008, 8, 634–646. [Google Scholar] [CrossRef]
- Aziz, N.; Kim, M.-Y.; Cho, J.Y. Anti-Inflammatory Effects of Luteolin: A Review of in Vitro, in Vivo, and in Silico Studies. J. Ethnopharmacol. 2018, 225, 342–358. [Google Scholar] [CrossRef] [PubMed]
- Singh Tuli, H.; Rath, P.; Chauhan, A.; Sak, K.; Aggarwal, D.; Choudhary, R.; Sharma, U.; Vashishth, K.; Sharma, S.; Kumar, M.; et al. Luteolin, a Potent Anticancer Compound: From Chemistry to Cellular Interactions and Synergetic Perspectives. Cancers 2022, 14, 5373. [Google Scholar] [CrossRef]
- Ganai, S.A.; Sheikh, F.A.; Baba, Z.A.; Mir, M.A.; Mantoo, M.A.; Yatoo, M.A. Anticancer Activity of the Plant Flavonoid Luteolin against Preclinical Models of Various Cancers and Insights on Different Signalling Mechanisms Modulated. Phytother. Res. 2021, 35, 3509–3532. [Google Scholar] [CrossRef]
- Yang, K.; Song, Y.; Ge, L.; Su, J.; Wen, Y.; Long, Y. Measurement and Correlation of the Solubilities of Luteolin and Rutin in Five Imidazole-Based Ionic Liquids. Fluid Phase Equilibria 2013, 344, 27–31. [Google Scholar] [CrossRef]
- Marín, L.; Gutiérrez-del-Río, I.; Yagüe, P.; Manteca, Á.; Villar, C.J.; Lombó, F. De Novo Biosynthesis of Apigenin, Luteolin, and Eriodictyol in the Actinomycete Streptomyces Albus and Production Improvement by Feeding and Spore Conditioning. Front. Microbiol. 2017, 8, 921. [Google Scholar] [CrossRef] [Green Version]
- le Marchand, L. Cancer Preventive Effects of Flavonoids—A Review. Biomed. Pharmacother. 2002, 56, 296–301. [Google Scholar] [CrossRef]
- Baroni, L.; Sarni, A.R.; Zuliani, C. Plant Foods Rich in Antioxidants and Human Cognition: A Systematic Review. Antioxidants 2021, 10, 714. [Google Scholar] [CrossRef]
- Zang, Y.; Igarashi, K.; Li, Y. Anti-Diabetic Effects of Luteolin and Luteolin-7-O-Glucoside on KK-A y Mice. Biosci. Biotechnol. Biochem. 2016, 80, 1580–1586. [Google Scholar] [CrossRef] [Green Version]
- Park, C.M.; Song, Y.-S. Luteolin and Luteolin-7-O-Glucoside Inhibit Lipopolysaccharide-Induced Inflammatory Responses through Modulation of NF-ΚB/AP-1/PI3K-Akt Signaling Cascades in RAW 264.7 Cells. Nutr. Res. Pract. 2013, 7, 423–429. [Google Scholar] [CrossRef] [Green Version]
- Sayre, C.L.; Gerde, K.D.; Yáñez, J.A.; Davies, N.M. Clinical Pharmacokinetics of Flavonoids. In Flavonoid Pharmacokinetics; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2012; pp. 195–247. ISBN 9781118468524. [Google Scholar]
- Chen, T.; Li; Lu, X.-Y.; Jiang, H.-D.; Zeng, S. Absorption and Excretion of Luteolin and Apigenin in Rats after Oral Administration of Chrysanthemum Morifolium Extract. J. Agric. Food Chem. 2007, 55, 273–277. [Google Scholar] [CrossRef]
- Yasuda, M.T.; Fujita, K.; Hosoya, T.; Imai, S.; Shimoi, K. Absorption and Metabolism of Luteolin and Its Glycosides from the Extract of Chrysanthemum Morifolium Flowers in Rats and Caco-2 Cells. J. Agric. Food Chem. 2015, 63, 7693–7699. [Google Scholar] [CrossRef]
- Shimoi, K.; Okada, H.; Furugori, M.; Goda, T.; Takase, S.; Suzuki, M.; Hara, Y.; Yamamoto, H.; Kinae, N. Intestinal Absorption of Luteolin and Luteolin 7-O-β-Glucoside in Rats and Humans. FEBS Lett. 1998, 438, 220–224. [Google Scholar] [CrossRef]
- Yin, L.; Shi, M.; Sun, Y.; Sun, X.; Meng, H.; Fawcett, J.P.; Yang, Y.; Gu, J. A Liquid Chromatography–Tandem Mass Spectrometric Method for the Simultaneous Quantitation of Five Components of Ixeris Sonchifoliain (Bge.) Hance in Rat Plasma and Its Application to a Pharmacokinetic Study. J. Chromatogr. B 2013, 931, 12–16. [Google Scholar] [CrossRef] [PubMed]
- Kure, A.; Nakagawa, K.; Kondo, M.; Kato, S.; Kimura, F.; Watanabe, A.; Shoji, N.; Hatanaka, S.; Tsushida, T.; Miyazawa, T. Metabolic Fate of Luteolin in Rats: Its Relationship to Anti-Inflammatory Effect. J. Agric. Food Chem. 2016, 64, 4246–4254. [Google Scholar] [CrossRef]
- Wang, L.; Chen, Q.; Zhu, L.; Li, Q.; Zeng, X.; Lu, L.; Hu, M.; Wang, X.; Liu, Z. Metabolic Disposition of Luteolin Is Mediated by the Interplay of UDP-Glucuronosyltransferases and Catechol-O-methyltransferases in rats. Drug Metab. Dispos. 2017, 45, 306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaikwad, A.; Ahire, K.; Gosavi, A.; Salunkhe, K. Phytosome as a Novel Drug Delivery System for Bioavailability Enhancement of Phytoconstituents and Its Applications: A Review. J. Drug Deliv. Ther. 2021, 11, 138–152. [Google Scholar] [CrossRef]
- Teng, Z.; Yuan, C.; Zhang, F.; Huan, M.; Cao, W.; Li, K.; Yang, J.; Cao, D.; Zhou, S.; Mei, Q. Intestinal Absorption and First-Pass Metabolism of Polyphenol Compounds in Rat and Their Transport Dynamics in Caco-2 Cells. PLoS ONE 2012, 7, e29647. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Wang, Q.; Morris, M.E. Pharmacokinetic Interaction between the Flavonoid Luteolin and γ-Hydroxybutyrate in Rats: Potential Involvement of Monocarboxylate Transporters. AAPS J. 2008, 10, 47–55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dang, H.; Meng, M.H.W.; Zhao, H.; Iqbal, J.; Dai, R.; Deng, Y.; Lv, F. Luteolin-Loaded Solid Lipid Nanoparticles Synthesis, Characterization, & Improvement of Bioavailability, Pharmacokinetics in Vitro and Vivo Studies. J. Nanopart. Res. 2014, 16, 2347. [Google Scholar] [CrossRef]
- Qing, W.; Wang, Y.; Li, H.; Ma, F.; Zhu, J.; Liu, X. Preparation and Characterization of Copolymer Micelles for the Solubilization and In Vitro Release of Luteolin and Luteoloside. AAPS PharmSciTech 2017, 18, 2095–2101. [Google Scholar] [CrossRef]
- Khan, J.; Saraf, S.; Saraf, S. Preparation and Evaluation of Luteolin–Phospholipid Complex as an Effective Drug Delivery Tool against GalN/LPS Induced Liver Damage. Pharm. Dev. Technol. 2016, 21, 475–486. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.; Li, K.; Zhao, C.; Ran, X.; Zhang, Y.; Zhang, T. A Rapid HPLC–MS/MS Method for the Simultaneous Determination of Luteolin, Resveratrol and Their Metabolites in Rat Plasma and Its Application to Pharmacokinetic Interaction Studies. J. Chromatogr. B 2022, 1191, 123118. [Google Scholar] [CrossRef]
- Tuorkey, M.J. Molecular Targets of Luteolin in Cancer. Eur. J. Cancer Prev. 2016, 25, 65–76. [Google Scholar] [CrossRef] [Green Version]
- Seelinger, G.; Merfort, I.; Wölfle, U.; Schempp, C.M. Anti-Carcinogenic Effects of the Flavonoid Luteolin. Molecules 2008, 13, 2628–2651. [Google Scholar] [CrossRef] [Green Version]
- Selvendiran, K.; Koga, H.; Ueno, T.; Yoshida, T.; Maeyama, M.; Torimura, T.; Yano, H.; Kojiro, M.; Sata, M. Luteolin Promotes Degradation in Signal Transducer and Activator of Transcription 3 in Human Hepatoma Cells: An Implication for the Antitumor Potential of Flavonoids. Cancer Res. 2006, 66, 4826–4834. [Google Scholar] [CrossRef] [Green Version]
- Pandurangan, A.K.; Kumar, S.A.S.; Dharmalingam, P.; Ganapasam, S. Luteolin, a Bioflavonoid Inhibits Azoxymethane-Induced Colon Carcinogenesis: Involvement of INOS and COX-2. Pharmacogn. Mag. 2014, 10, S306. [Google Scholar]
- Pandurangan, A.K.; Dharmalingam, P.; Sadagopan, S.K.A.; Ganapasam, S. Luteolin Inhibits Matrix Metalloproteinase 9 and 2 in Azoxymethane-Induced Colon Carcinogenesis. Hum. Exp. Toxicol. 2014, 33, 1176–1185. [Google Scholar] [CrossRef]
- Pandurangan, A.K.; Ananda Sadagopan, S.K.; Dharmalingam, P.; Ganapasam, S. Luteolin, a Bioflavonoid Inhibits Azoxymethane-Induced Colorectal Cancer through Activation of Nrf2 Signaling. Toxicol. Mech. Methods 2014, 24, 13–20. [Google Scholar] [CrossRef] [PubMed]
- Pandurangan, A.K.; Dharmalingam, P.; Sadagopan, S.K.A.; Ramar, M.; Munusamy, A. Luteolin Induces Growth Arrest in Colon Cancer Cells through Involvement of Wnt/β-Catenin/GSK-3β Signaling. J. Environ. Pathol. Toxicol. Oncol. 2013, 32, 131–139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pandurangan, A.K.; Ganapsam, S. Luteolin Induces Apoptosis in Azoxymethane-Induced Colon Carcinogenesis through the Involvement of Bcl-2, Bax and Caspase-3. J. Chem. Pharm. Res. 2013, 5, 143–148. [Google Scholar]
- Kang, K.A.; Piao, M.J.; Ryu, Y.S.; Hyun, Y.J.; Park, J.E.; Shilnikova, K.; Zhen, A.X.; Kang, H.K.; Koh, Y.S.; Jeong, Y.J. Luteolin Induces Apoptotic Cell Death via Antioxidant Activity in Human Colon Cancer Cells. Int. J. Oncol. 2017, 51, 1169–1178. [Google Scholar] [CrossRef] [Green Version]
- Kang, K.A.; Piao, M.J.; Hyun, Y.J.; Zhen, A.X.; Cho, S.J.; Ahn, M.J.; Yi, J.M.; Hyun, J.W. Luteolin Promotes Apoptotic Cell Death via Upregulation of Nrf2 Expression by DNA Demethylase and the Interaction of Nrf2 with P53 in Human Colon Cancer Cells. Exp. Mol. Med. 2019, 51, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Yoo, H.S.; Won, S.B.; Kwon, Y.H. Luteolin Induces Apoptosis and Autophagy in HCT116 Colon Cancer Cells via P53-Dependent Pathway. Nutr. Cancer 2022, 74, 677–686. [Google Scholar] [CrossRef]
- Chen, Z.; Zhang, B.; Gao, F.; Shi, R. Modulation of G2/M Cell Cycle Arrest and Apoptosis by Luteolin in Human Colon Cancer Cells and Xenografts. Oncol. Lett. 2018, 15, 1559–1565. [Google Scholar] [CrossRef] [Green Version]
- Song, Y.; Yu, J.; Li, L.; Wang, L.; Dong, L.; Xi, G.; Lu, Y.J.; Li, Z. Luteolin Impacts Deoxyribonucleic Acid Repair by Modulating the Mitogen-Activated Protein Kinase Pathway in Colorectal Cancer. Bioengineered 2022, 13, 10998–11011. [Google Scholar] [CrossRef]
- Jang, C.H.; Moon, N.; Oh, J.; Kim, J.-S. Adverse Effect of Luteolin on the Anticancer Ability of Oxaliplatin in HCT116 Human Colorectal Carcinoma Cells. FASEB J. 2019, 33, lb602. [Google Scholar] [CrossRef]
- Aromokeye, R.; Si, H. Combined Curcumin and Luteolin Synergistically Inhibit Colon Cancer Associated with Notch1 and TGF-β Signaling Pathways in Cultured Cells and Xenograft Mice. Cancers 2022, 14, 3001. [Google Scholar] [CrossRef]
- Jiang, J.; Zhu, F.; Zhang, H.; Sun, T.; Fu, F.; Chen, X.; Zhang, Y. Luteolin Suppresses the Growth of Colon Cancer Cells by Inhibiting the IL-6/STAT3 Signaling Pathway. J. Gastrointest. Oncol. 2022, 13, 1722–1732. [Google Scholar] [CrossRef] [PubMed]
- Yao, Y.; Rao, C.; Zheng, G.; Wang, S. Luteolin Suppresses Colorectal Cancer Cell Metastasis via Regulation of the MiR-384/Pleiotrophin Axis. Oncol. Rep. 2019, 42, 131–141. [Google Scholar] [CrossRef]
- Meng, G.; Chai, K.; Li, X.; Zhu, Y.; Huang, W. Luteolin Exerts Pro-Apoptotic Effect and Anti-Migration Effects on A549 Lung Adenocarcinoma Cells through the Activation of MEK/ERK Signaling Pathway. Chem. Biol. Interact. 2016, 257, 26–34. [Google Scholar] [CrossRef]
- Park, S.-H.; Park, H.S.; Lee, J.H.; Chi, G.Y.; Kim, G.-Y.; Moon, S.-K.; Chang, Y.-C.; Hyun, J.W.; Kim, W.-J.; Choi, Y.H. Induction of Endoplasmic Reticulum Stress-Mediated Apoptosis and Non-Canonical Autophagy by Luteolin in NCI-H460 Lung Carcinoma Cells. Food Chem. Toxicol. 2013, 56, 100–109. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.; Peng, H.; Li, K.; Zhao, R.; Li, L.; Yu, Y.; Wang, X.; Han, Z. Luteolin Exerts an Anticancer Effect on NCI-H460 Human Non-Small Cell Lung Cancer Cells through the Induction of Sirt1-Mediated Apoptosis. Mol. Med. Rep. 2015, 12, 4196–4202. [Google Scholar] [CrossRef] [Green Version]
- Zhang, M.; Wang, R.; Tian, J.; Song, M.; Zhao, R.; Liu, K.; Zhu, F.; Shim, J.-H.; Dong, Z.; Lee, M.-H. Targeting LIMK1 with Luteolin Inhibits the Growth of Lung Cancer in Vitro and in Vivo. J. Cell Mol. Med. 2021, 25, 5560–5571. [Google Scholar] [CrossRef]
- Dallavalasa, S.; Beeraka, N.M.; Basavaraju, C.G.; Tulimilli, S.V.; Sadhu, S.P.; Rajesh, K.; Aliev, G.; Madhunapantula, S.V. The Role of Tumor Associated Macrophages (TAMs) in Cancer Progression, Chemoresistance, Angiogenesis and Metastasis-Current Status. Curr. Med. Chem. 2021, 28, 8203–8236. [Google Scholar] [CrossRef]
- Choi, H.-J.; Choi, H.-J.; Chung, T.-W.; Ha, K.-T. Luteolin Inhibits Recruitment of Monocytes and Migration of Lewis Lung Carcinoma Cells by Suppressing Chemokine (C–C Motif) Ligand 2 Expression in Tumor-Associated Macrophage. Biochem. Biophys. Res. Commun. 2016, 470, 101–106. [Google Scholar] [CrossRef] [PubMed]
- Pardoll, D.M. The Blockade of Immune Checkpoints in Cancer Immunotherapy. Nat. Rev. Cancer 2012, 12, 252–264. [Google Scholar] [CrossRef] [Green Version]
- Yadav, D.; Kwak, M.; Chauhan, P.S.; Puranik, N.; Lee, P.C.W.; Jin, J.-O. Cancer Immunotherapy by Immune Checkpoint Blockade and Its Advanced Application Using Bio-Nanomaterials. Semin. Cancer Biol. 2022, 86, 909–922. [Google Scholar] [CrossRef]
- Jiang, Z.-B.; Wang, W.-J.; Xu, C.; Xie, Y.-J.; Wang, X.-R.; Zhang, Y.-Z.; Huang, J.-M.; Huang, M.; Xie, C.; Liu, P.; et al. Luteolin and Its Derivative Apigenin Suppress the Inducible PD-L1 Expression to Improve Anti-Tumor Immunity in KRAS-Mutant Lung Cancer. Cancer Lett. 2021, 515, 36–48. [Google Scholar] [CrossRef] [PubMed]
- Begg, A.C.; Stewart, F.A.; Vens, C. Strategies to Improve Radiotherapy with Targeted Drugs. Nat. Rev. Cancer 2011, 11, 239–253. [Google Scholar] [CrossRef]
- Cho, H.; Ahn, K.; Choi, J.Y.; Hwang, S.; Kim, W.; Um, H.; Park, J.K. Luteolin Acts as a Radiosensitizer in Non-small Cell Lung Cancer Cells by Enhancing Apoptotic Cell Death through Activation of a P38/ROS/Caspase Cascade. Int. J. Oncol. 2015, 46, 1149–1158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, S.; El-Deiry, W.S. TRAIL and Apoptosis Induction by TNF-Family Death Receptors. Oncogene 2003, 22, 8628–8633. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hellwig, C.T.; Rehm, M. TRAIL Signaling and Synergy Mechanisms Used in TRAIL-Based Combination Therapies. Mol. Cancer Ther. 2012, 11, 3–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, B.; Xiong, J.; Zhou, Y.; Wu, Y.; Song, Y.; Wang, N.; Chen, L.; Zhang, J. Luteolin Enhances TRAIL Sensitivity in Non-Small Cell Lung Cancer Cells through Increasing DR5 Expression and Drp1-Mediated Mitochondrial Fission. Arch. Biochem. Biophys. 2020, 692, 108539. [Google Scholar] [CrossRef]
- Chen, K.-C.; Chen, C.-Y.; Lin, C.-J.; Yang, T.-Y.; Chen, T.-H.; Wu, L.-C.; Wu, C.-C. Luteolin Attenuates TGF-Β1-Induced Epithelial–Mesenchymal Transition of Lung Cancer Cells by Interfering in the PI3K/Akt–NF-ΚB–Snail Pathway. Life Sci. 2013, 93, 924–933. [Google Scholar] [CrossRef]
- Masraksa, W.; Tanasawet, S.; Hutamekalin, P.; Wongtawatchai, T.; Sukketsiri, W. Luteolin Attenuates Migration and Invasion of Lung Cancer Cells via Suppressing Focal Adhesion Kinase and Non-Receptor Tyrosine Kinase Signaling Pathway. Nutr. Res. Pract. 2020, 14, 127–133. [Google Scholar] [CrossRef]
- Çetinkaya, M.; Baran, Y. MicroRNAs and Long Non-Coding RNAs as Novel Targets in Anti-Cancer Drug Development. Curr. Pharm. Biotechnol. 2022, 23. [Google Scholar] [CrossRef]
- Jiang, Z.-Q.; Li, M.-H.; Qin, Y.-M.; Jiang, H.-Y.; Zhang, X.; Wu, M.-H. Luteolin Inhibits Tumorigenesis and Induces Apoptosis of Non-Small Cell Lung Cancer Cells via Regulation of MicroRNA-34a-5p. Int. J. Mol. Sci. 2018, 19, 447. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pan, J.; Cai, X.; Zheng, X.; Zhu, X.; Feng, J.; Wang, X. Luteolin Inhibits Viability, Migration, Angiogenesis and Invasion of Non-Small Cell Lung Cancer Vascular Endothelial Cells via MiR-133a-3p/Purine Rich Element Binding Protein B-Mediated MAPK and PI3K/Akt Signaling Pathways. Tissue Cell 2022, 75, 101740. [Google Scholar] [CrossRef] [PubMed]
- Zheng, H.; Zhu, X.; Gong, E.; Lv, Y.; Li, Y.; Cai, X. Luteolin Suppresses Lung Cancer Progression through Targeting the Circ_0000190/MiR-130a-3p/Notch-1 Signaling Pathway. J. Chemother. 2022, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Q.; Yan, B.; Hu, X.; Li, X.-B.; Zhang, J.; Fang, J. Luteolin Inhibits Invasion of Prostate Cancer PC3 Cells through E-Cadherin. Mol. Cancer Ther. 2009, 8, 1684–1691. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsai, P.-H.; Cheng, C.-H.; Lin, C.-Y.; Huang, Y.-T.; Lee, L.-T.; Kandaswami, C.C.; Lin, Y.-C.; Lee, K.P.-H.; Hung, C.-C.; Hwang, J.-J. Dietary Flavonoids Luteolin and Quercetin Suppressed Cancer Stem Cell Properties and Metastatic Potential of Isolated Prostate Cancer Cells. Anticancer. Res. 2016, 36, 6367–6380. [Google Scholar] [CrossRef] [Green Version]
- Pratheeshkumar, P.; Son, Y.-O.; Budhraja, A.; Wang, X.; Ding, S.; Wang, L.; Hitron, A.; Lee, J.-C.; Kim, D.; Divya, S.P. Luteolin Inhibits Human Prostate Tumor Growth by Suppressing Vascular Endothelial Growth Factor Receptor 2-Mediated Angiogenesis. PLoS ONE 2012, 7, e52279. [Google Scholar] [CrossRef] [PubMed]
- Han, K.; Lang, T.; Zhang, Z.; Zhang, Y.; Sun, Y.; Shen, Z.; Beuerman, R.W.; Zhou, L.; Min, D. Luteolin Attenuates Wnt Signaling via Upregulation of FZD6 to Suppress Prostate Cancer Stemness Revealed by Comparative Proteomics. Sci. Rep. 2018, 8, 8537. [Google Scholar] [CrossRef] [Green Version]
- Tsui, K.-H.; Chung, L.-C.; Feng, T.-H.; Chang, P.-L.; Juang, H.-H. Upregulation of Prostate-Derived Ets Factor by Luteolin Causes Inhibition of Cell Proliferation and Cell Invasion in Prostate Carcinoma Cells. Int. J. Cancer 2012, 130, 2812–2823. [Google Scholar] [CrossRef] [PubMed]
- Markaverich, B.M.; Vijjeswarapu, M. Multiple Sites of Type II Site Ligand (Luteolin and BMHPC) Regulation of Gene Expression in PC-3 Cells. Int. J. Biomed. Sci. 2012, 8, 219. [Google Scholar]
- Wang, L.; Li, W.; Lin, M.; Garcia, M.; Mulholland, D.; Lilly, M.; Martins-Green, M. Luteolin, Ellagic Acid and Punicic Acid Are Natural Products That Inhibit Prostate Cancer Metastasis. Carcinogenesis 2014, 35, 2321–2330. [Google Scholar] [CrossRef] [Green Version]
- Gray, A.L.; Stephens, C.A.; Bigelow, R.L.H.; Coleman, D.T.; Cardelli, J.A. The Polyphenols (−)-Epigallocatechin-3-Gallate and Luteolin Synergistically Inhibit TGF-β-Induced Myofibroblast Phenotypes through RhoA and ERK Inhibition. PLoS ONE 2014, 9, e109208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakurai, M.A.; Ozaki, Y.; Okuzaki, D.; Naito, Y.; Sasakura, T.; Okamoto, A.; Tabara, H.; Inoue, T.; Hagiyama, M.; Ito, A. Gefitinib and Luteolin Cause Growth Arrest of Human Prostate Cancer PC-3 Cells via Inhibition of Cyclin G-Associated Kinase and Induction of MiR-630. PLoS ONE 2014, 9, e100124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, K.; Meng, W.; Zhang, J.; Zhou, Y.; Wang, Y.; Su, Y.; Lin, S.; Gan, Z.; Sun, Y.; Min, D. Luteolin Inhibited Proliferation and Induced Apoptosis of Prostate Cancer Cells through MiR-301. Oncol. Targets Ther. 2016, 9, 3085. [Google Scholar] [CrossRef] [Green Version]
- Pu, Y.; Zhang, T.; Wang, J.; Mao, Z.; Duan, B.; Long, Y.; Xue, F.; Liu, D.; Liu, S.; Gao, Z. Luteolin Exerts an Anticancer Effect on Gastric Cancer Cells through Multiple Signaling Pathways and Regulating MiRNAs. J. Cancer 2018, 9, 3669–3675. [Google Scholar] [CrossRef] [Green Version]
- Lu, X.Y.; Li, Y.H.; Xiao, X.W.; Li, X.B. Inhibitory Effects of Luteolin on Human Gastric Carcinoma Xenografts in Nude Mice and Its Mechanism. Zhonghua Yi Xue Za Zhi 2013, 93, 142–146. [Google Scholar] [PubMed]
- Lu, J.; Li, G.; He, K.; Jiang, W.; Xu, C.; Li, Z.; Wang, H.; Wang, W.; Wang, H.; Teng, X.; et al. Luteolin Exerts a Marked Antitumor Effect in CMet-Overexpressing Patient-Derived Tumor Xenograft Models of Gastric Cancer. J. Transl. Med. 2015, 13, 42. [Google Scholar] [CrossRef] [Green Version]
- Zang, M.; Hu, L.; Fan, Z.; Wang, H.; Zhu, Z.; Cao, S.; Wu, X.; Li, J.; Su, L.; Li, C.; et al. Luteolin Suppresses Gastric Cancer Progression by Reversing Epithelial-Mesenchymal Transition via Suppression of the Notch Signaling Pathway. J. Transl. Med. 2017, 15, 52. [Google Scholar] [CrossRef] [Green Version]
- Zang, M.; Hu, L.; Zhang, B.; Zhu, Z.; Li, J.; Zhu, Z.; Yan, M.; Liu, B. Luteolin Suppresses Angiogenesis and Vasculogenic Mimicry Formation through Inhibiting Notch1-VEGF Signaling in Gastric Cancer. Biochem. Biophys. Res. Commun. 2017, 490, 913–919. [Google Scholar] [CrossRef]
- Zhang, L.; Liao, Y.; Tang, L. MicroRNA-34 Family: A Potential Tumor Suppressor and Therapeutic Candidate in Cancer. J. Exp. Clin. Cancer Res. 2019, 38, 53. [Google Scholar] [CrossRef] [Green Version]
- Wu, H.; Huang, M.; Liu, Y.; Shu, Y.; Liu, P. Luteolin Induces Apoptosis by Up-Regulating MiR-34a in Human Gastric Cancer Cells. Technol. Cancer Res. Treat. 2014, 14, 747–755. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.; Ding, B.-Z.; Lin, Y.-P.; Wang, H.-B. MiR-34a, as a Suppressor, Enhance the Susceptibility of Gastric Cancer Cell to Luteolin by Directly Targeting HK1. Gene 2018, 644, 56–65. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Wan, L.; Guo, Y.; Cheng, N.; Cheng, W.; Sun, Q.; Zhu, J. Radiosensitization Effect of Luteolin on Human Gastric Cancer SGC-7901 Cells. J. Biol. Regul. Homeost. Agents 2009, 23, 71–78. [Google Scholar] [PubMed]
- Ren, L.-Q.; Li, Q.; Zhang, Y. Luteolin Suppresses the Proliferation of Gastric Cancer Cells and Acts in Synergy with Oxaliplatin. Biomed. Res. Int. 2020, 2020, 9396512. [Google Scholar] [CrossRef] [Green Version]
- Sawmiller, D.; Li, S.; Shahaduzzaman, M.; Smith, A.J.; Obregon, D.; Giunta, B.; Borlongan, C.V.; Sanberg, P.R.; Tan, J. Luteolin Reduces Alzheimer’s Disease Pathologies Induced by Traumatic Brain Injury. Int. J. Mol. Sci. 2014, 15, 895–904. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, H.-S.; Park, B.-S.; Kang, H.-M.; Kim, J.-H.; Shin, S.-H.; Kim, I.-R. Role of Luteolin-Induced Apoptosis and Autophagy in Human Glioblastoma Cell Lines. Medicina 2021, 57, 879. [Google Scholar] [CrossRef]
- Franco, Y.E.M.; de Lima, C.A.; Rosa, M.N.; Silva, V.A.O.; Reis, R.M.; Priolli, D.G.; Carvalho, P.O.; do Nascimento, J.R.; da Rocha, C.Q.; Longato, G.B. Investigation of U-251 Cell Death Triggered by Flavonoid Luteolin: Towards a Better Understanding on Its Anticancer Property against Glioblastomas. Nat. Prod. Res. 2021, 35, 4807–4813. [Google Scholar] [CrossRef]
- Anson, D.M.; Wilcox, R.M.; Huseman, E.D.; Stump, T.A.; Paris, R.L.; Darkwah, B.O.; Lin, S.; Adegoke, A.O.; Gryka, R.J.; Jean-Louis, D.S.; et al. Luteolin Decreases Epidermal Growth Factor Receptor-Mediated Cell Proliferation and Induces Apoptosis in Glioblastoma Cell Lines. Basic. Clin. Pharmacol. Toxicol. 2018, 123, 678–686. [Google Scholar] [CrossRef]
- Powe, E.; Parschauer, D.; Istifan, J.; Lin, S.; Duan, H.; Gryka, R.; Jean-Louis, D.; Tiwari, A.K.; Amos, S. Luteolin Enhances Erlotinib’s Cell Proliferation Inhibitory and Apoptotic Effects in Glioblastoma Cell Lines. Front. Pharmacol. 2022, 13, 3963. [Google Scholar] [CrossRef]
- Cheng, W.-Y.; Chiao, M.-T.; Liang, Y.-J.; Yang, Y.-C.; Shen, C.-C.; Yang, C.-Y. Luteolin Inhibits Migration of Human Glioblastoma U-87 MG and T98G Cells through Downregulation of Cdc42 Expression and PI3K/AKT Activity. Mol. Biol. Rep. 2013, 40, 5315–5326. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Wang, H.; Jia, Y.; Ding, H.; Zhang, L.; Pan, H. Luteolin Reduces Migration of Human Glioblastoma Cell Lines via Inhibition of the P-IGF-1R/PI3K/AKT/MTOR Signaling Pathway. Oncol. Lett. 2017, 14, 3545–3551. [Google Scholar] [CrossRef] [Green Version]
- Chakrabarti, M.; Ray, S.K. Synergistic Anti-Tumor Actions of Luteolin and Silibinin Prevented Cell Migration and Invasion and Induced Apoptosis in Glioblastoma SNB19 Cells and Glioblastoma Stem Cells. Brain. Res. 2015, 1629, 85–93. [Google Scholar] [CrossRef] [PubMed]
- Chakrabarti, M.; Ray, S.K. Anti-Tumor Activities of Luteolin and Silibinin in Glioblastoma Cells: Overexpression of MiR-7-1-3p Augmented Luteolin and Silibinin to Inhibit Autophagy and Induce Apoptosis in Glioblastoma in Vivo. Apoptosis 2016, 21, 312–328. [Google Scholar] [CrossRef] [PubMed]
- Ding, S.; Hu, A.; Hu, Y.; Ma, J.; Weng, P.; Dai, J. Anti-Hepatoma Cells Function of Luteolin through Inducing Apoptosis and Cell Cycle Arrest. Tumor Biol. 2014, 35, 3053–3060. [Google Scholar] [CrossRef] [PubMed]
- Hwang, J.-T.; Park, O.J.; Lee, Y.K.; Sung, M.J.; Hur, H.J.; Kim, M.S.; Ha, J.H.; Kwon, D.Y. Anti-Tumor Effect of Luteolin Is Accompanied by AMP-Activated Protein Kinase and Nuclear Factor-ΚB Modulation in HepG2 Hepatocarcinoma Cells. Int. J. Mol. Med. 2011, 28, 25–31. [Google Scholar] [PubMed] [Green Version]
- Im, E.; Yeo, C.; Lee, E.-O. Luteolin Induces Caspase-Dependent Apoptosis via Inhibiting the AKT/Osteopontin Pathway in Human Hepatocellular Carcinoma SK-Hep-1 Cells. Life Sci. 2018, 209, 259–266. [Google Scholar] [CrossRef]
- Chakraborty, G.; Jain, S.; Behera, R.; Ahmed, M.; Sharma, P.; Kumar, V.; Kundu, G.C. The Multifaceted Roles of Osteopontin in Cell Signaling, Tumor Progression and Angiogenesis. Curr. Mol. Med. 2006, 6, 819–830. [Google Scholar] [CrossRef]
- Saleh, S.; Thompson, D.E.; McConkey, J.; Murray, P.; Moorehead, R.A. Osteopontin Regulates Proliferation, Apoptosis, and Migration of Murine Claudin-Low Mammary Tumor Cells. BMC Cancer 2016, 16, 359. [Google Scholar] [CrossRef] [Green Version]
- Niu, J.X.; Guo, H.P.; Gan, H.M.; Bao, L.D.; Ren, J.J. Effect of Luteolin on Gene Expression in Mouse H22 Hepatoma Cells. Genet. Mol. Res. 2015, 14, 14448–14456. [Google Scholar] [CrossRef]
- Xu, H.; Yang, T.; Liu, X.; Tian, Y.; Chen, X.; Yuan, R.; Su, S.; Lin, X.; Du, G. Luteolin Synergizes the Antitumor Effects of 5-Fluorouracil against Human Hepatocellular Carcinoma Cells through Apoptosis Induction and Metabolism. Life Sci. 2016, 144, 138–147. [Google Scholar] [CrossRef]
- Nazim, U.M.; Park, S. Luteolin Sensitizes Human Liver Cancer Cells to TRAIL-induced Apoptosis via Autophagy and JNK-mediated Death Receptor 5 Upregulation. Int. J. Oncol. 2019, 54, 665–672. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, X.-Y.; Li, H.-G.; Zhang, K.-J.; Gu, J.-F. Strategy of Cancer Targeting Gene-Viro-Therapy (CTGVT) a Trend in Both Cancer Gene Therapy and Cancer Virotherapy. Curr. Pharm. Biotechnol. 2012, 13, 1761–1767. [Google Scholar] [CrossRef]
- Wang, C.; Li, Q.; Xiao, B.; Fang, H.; Huang, B.; Huang, F.; Wang, Y. Luteolin Enhances the Antitumor Efficacy of Oncolytic Vaccinia Virus That Harbors IL-24 Gene in Liver Cancer Cells. J. Clin. Lab. Anal. 2021, 35, e23677. [Google Scholar] [CrossRef]
- Cao, Z.; Zhang, H.; Cai, X.; Fang, W.; Chai, D.; Wen, Y.; Chen, H.; Chu, F.; Zhang, Y. Luteolin Promotes Cell Apoptosis by Inducing Autophagy in Hepatocellular Carcinoma. Cell. Physiol. Biochem. 2017, 43, 1803–1812. [Google Scholar] [CrossRef]
- Lee, Y.; Kwon, Y.H. Regulation of Apoptosis and Autophagy by Luteolin in Human Hepatocellular Cancer Hep3B Cells. Biochem. Biophys. Res. Commun. 2019, 517, 617–622. [Google Scholar] [CrossRef]
- Ahmed, S.; Khan, H.; Fratantonio, D.; Hasan, M.M.; Sharifi, S.; Fathi, N.; Ullah, H.; Rastrelli, L. Apoptosis Induced by Luteolin in Breast Cancer: Mechanistic and Therapeutic Perspectives. Phytomedicine 2019, 59, 152883. [Google Scholar] [CrossRef]
- Sui, J.-Q.; Xie, K.-P.; Xie, M.-J. Inhibitory Effect of Luteolin on the Proliferation of Human Breast Cancer Cell Lines Induced by Epidermal Growth Factor. Sheng Li Xue Bao Acta Physiol. Sin. 2016, 68, 27–34. [Google Scholar]
- Wang, L.-M.; Xie, K.-P.; Huo, H.-N.; Shang, F.; Zou, W.; Xie, M.-J. Luteolin Inhibits Proliferation Induced by IGF-1 Pathway Dependent ERα in Human Breast Cancer MCF-7 Cells. Asian Pac. J. Cancer Prev. 2012, 13, 1431–1437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monti, E.; Marras, E.; Prini, P.; Gariboldi, M.B. Luteolin Impairs Hypoxia Adaptation and Progression in Human Breast and Colon Cancer Cells. Eur. J. Pharmacol. 2020, 881, 173210. [Google Scholar] [CrossRef] [PubMed]
- Lee, E.-J.; Oh, S.-Y.; Sung, M.-K. Luteolin Exerts Anti-Tumor Activity through the Suppression of Epidermal Growth Factor Receptor-Mediated Pathway in MDA-MB-231 ER-Negative Breast Cancer Cells. Food Chem. Toxicol. 2012, 50, 4136–4143. [Google Scholar] [CrossRef] [PubMed]
- Park, S.-H.; Ham, S.; Kwon, T.H.; Kim, M.S.; Lee, D.H.; Kang, J.-W.; Oh, S.-R.; Yoon, D.-Y. Luteolin Induces Cell Cycle Arrest and Apoptosis through Extrinsic and Intrinsic Signaling Pathways in MCF-7 Breast Cancer Cells. J. Environ. Pathol. Toxicol. Oncol. 2014, 33, 219–231. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Jin, K.; Lan, H. Luteolin Inhibits Cell Cycle Progression and Induces Apoptosis of Breast Cancer Cells through Downregulation of Human Telomerase Reverse Transcriptase. Oncol. Lett. 2019, 17, 3842–3850. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, H.-T.; Liu, Y.-E.; Hsu, K.-W.; Wang, Y.-F.; Chan, Y.-C.; Chen, Y.; Chen, D.-R. MLL3 Induced by Luteolin Causes Apoptosis in Tamoxifen-Resistant Breast Cancer Cells through H3K4 Monomethylation and Suppression of the PI3K/AKT/MTOR Pathway. Am. J. Chin. Med. 2020, 48, 1221–1241. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Xie, K.-P.; Huo, H.-N.; Wang, L.-M.; Zou, W.; Xie, M.-J. Inhibitory Effect of Luteolin on the Angiogenesis of Chick Chorioallantoic Membrane and Invasion of Breast Cancer Cells via Downregulation of AEG-1 and MMP-2. Sheng Li Xue Bao Acta Physiol. Sin. 2013, 65, 513–518. [Google Scholar]
- Park, S.-H.; Kim, J.-H.; Lee, D.-H.; Kang, J.-W.; Song, H.-H.; Oh, S.-R.; Yoon, D.-Y. Luteolin 8-C-β-Fucopyranoside Inhibits Invasion and Suppresses TPA-Induced MMP-9 and IL-8 via ERK/AP-1 and ERK/NF-ΚB Signaling in MCF-7 Breast Cancer Cells. Biochimie 2013, 95, 2082–2090. [Google Scholar] [CrossRef] [PubMed]
- Cook, M.T.; Liang, Y.; Besch-Williford, C.; Goyette, S.; Mafuvadze, B.; Hyder, S.M. Luteolin Inhibits Progestin-Dependent Angiogenesis, Stem Cell-like Characteristics, and Growth of Human Breast Cancer Xenografts. Springerplus 2015, 4, 444. [Google Scholar] [CrossRef] [Green Version]
- Lin, D.; Kuang, G.; Wan, J.; Zhang, X.; Li, H.; Gong, X.; Li, H. Luteolin Suppresses the Metastasis of Triple-Negative Breast Cancer by Reversing Epithelial-to-Mesenchymal Transition via Downregulation of β-Catenin Expression. Oncol. Rep. 2017, 37, 895–902. [Google Scholar] [CrossRef] [Green Version]
- Cao, D.; Zhu, G.-Y.; Lu, Y.; Yang, A.; Chen, D.; Huang, H.-J.; Peng, S.-X.; Chen, L.-W.; Li, Y.-W. Luteolin Suppresses Epithelial-Mesenchymal Transition and Migration of Triple-Negative Breast Cancer Cells by Inhibiting YAP/TAZ Activity. Biomed. Pharmacother. 2020, 129, 110462. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.-T.; Lin, J.; Liu, Y.-E.; Chen, H.-F.; Hsu, K.-W.; Lin, S.-H.; Peng, K.-Y.; Lin, K.-J.; Hsieh, C.-C.; Chen, D.-R. Luteolin Suppresses Androgen Receptor-Positive Triple-Negative Breast Cancer Cell Proliferation and Metastasis by Epigenetic Regulation of MMP9 Expression via the AKT/MTOR Signaling Pathway. Phytomedicine 2021, 81, 153437. [Google Scholar] [CrossRef]
- Sun, D.-W.; Zhang, H.-D.; Mao, L.; Mao, C.-F.; Chen, W.; Cui, M.; Ma, R.; Cao, H.-X.; Jing, C.-W.; Wang, Z. Luteolin Inhibits Breast Cancer Development and Progression in Vitro and in Vivo by Suppressing Notch Signaling and Regulating MiRNAs. Cell. Physiol. Biochem. 2015, 37, 1693–1711. [Google Scholar] [CrossRef] [Green Version]
- Gao, G.; Ge, R.; Li, Y.; Liu, S. Luteolin Exhibits Anti-Breast Cancer Property through up-Regulating MiR-203. Artif. Cells Nanomed. Biotechnol. 2019, 47, 3265–3271. [Google Scholar] [CrossRef] [Green Version]
- Shih, Y.-L.; Liu, H.-C.; Chen, C.-S.; Hsu, C.-H.; Pan, M.-H.; Chang, H.-W.; Chang, C.-H.; Chen, F.-C.; Ho, C.-T.; Yang, Y.-Y.; et al. Combination Treatment with Luteolin and Quercetin Enhances Antiproliferative Effects in Nicotine-Treated MDA-MB-231 Cells by Down-Regulating Nicotinic Acetylcholine Receptors. J. Agric. Food Chem. 2010, 58, 235–241. [Google Scholar] [CrossRef]
- Yang, Y.; Zheng, R.; Zhang, P.; Wen, J.; Luo, Y.; Zhao, Z.; You, L.; Ho, C.-T. Combination Effects of Polyphenols Present in Sugarcane on Proliferation in MCF-7 Human Breast Cancer Cells. Sugar Tech 2022, 24, 832–840. [Google Scholar] [CrossRef]
- Sato, Y.; Sasaki, N.; Saito, M.; Endo, N.; Kugawa, F.; Ueno, A. Luteolin Attenuates Doxorubicin-Induced Cytotoxicity to MCF-7 Human Breast Cancer Cells. Biol. Pharm. Bull. 2015, 38, 703–709. [Google Scholar] [CrossRef] [Green Version]
- Shi, Y.; Li, F.; Shen, M.; Sun, C.; Hao, W.; Wu, C.; Xie, Y.; Zhang, S.; Gao, H.; Yang, J. Luteolin Prevents Cardiac Dysfunction and Improves the Chemotherapeutic Efficacy of Doxorubicin in Breast Cancer. Front. Cardiovasc. Med. 2021, 8, 750186. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Yang, F.; Huang, L.; Liu, A.; Zhang, J. Luteolin Enhances the Antitumor Activity of Lapatinib in Human Breast Cancer Cells. Biomed. Res. 2017, 28, 4902–4907. [Google Scholar]
- Jeon, Y.-W.; Suh, Y.J. Synergistic Apoptotic Effect of Celecoxib and Luteolin on Breast Cancer Cells. Oncol. Rep. 2013, 29, 819–825. [Google Scholar] [CrossRef] [Green Version]
- Jeon, Y.W.; Ahn, Y.E.; Chung, W.S.; Choi, H.J.; Suh, Y.J. Synergistic Effect between Celecoxib and Luteolin Is Dependent on Estrogen Receptor in Human Breast Cancer Cells. Tumor Biol. 2015, 36, 6349–6359. [Google Scholar] [CrossRef]
- Yang, M.-Y.; Wang, C.-J.; Chen, N.-F.; Ho, W.-H.; Lu, F.-J.; Tseng, T.-H. Luteolin Enhances Paclitaxel-Induced Apoptosis in Human Breast Cancer MDA-MB-231 Cells by Blocking STAT3. Chem. Biol. Interact. 2014, 213, 60–68. [Google Scholar] [CrossRef] [PubMed]
- Tsai, K.-J.; Tsai, H.-Y.; Tsai, C.-C.; Chen, T.-Y.; Hsieh, T.-H.; Chen, C.-L.; Mbuyisa, L.; Huang, Y.-B.; Lin, M.-W. Luteolin Inhibits Breast Cancer Stemness and Enhances Chemosensitivity through the Nrf2-Mediated Pathway. Molecules 2021, 26, 6452. [Google Scholar] [CrossRef]
- Luo, Y.; Chen, S.; Zhou, J.; Chen, J.; Tian, L.; Gao, W.; Zhang, Y.; Ma, A.; Li, L.; Zhou, Z. Luteolin Cocrystals: Characterization, Evaluation of Solubility, Oral Bioavailability and Theoretical Calculation. J. Drug Deliv. Sci. Technol. 2019, 50, 248–254. [Google Scholar] [CrossRef]
- Sindhu, R.K.; Verma, R.; Salgotra, T.; Rahman, M.H.; Shah, M.; Akter, R.; Murad, W.; Mubin, S.; Bibi, P.; Qusti, S. Impacting the Remedial Potential of Nano Delivery-Based Flavonoids for Breast Cancer Treatment. Molecules 2021, 26, 5163. [Google Scholar] [CrossRef] [PubMed]
- Bernabeu, E.; Cagel, M.; Lagomarsino, E.; Moretton, M.; Chiappetta, D.A. Paclitaxel: What Has Been Done and the Challenges Remain Ahead. Int. J. Pharm. 2017, 526, 474–495. [Google Scholar] [CrossRef] [PubMed]
- Zheng, S.; Cheng, Y.; Teng, Y.; Liu, X.; Yu, T.; Wang, Y.; Liu, J.; Hu, Y.; Wu, C.; Wang, X. Application of Luteolin Nanomicelles Anti-Glioma Effect with Improvement in Vitro and in Vivo. Oncotarget 2017, 8, 61146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, C.; Xu, Q.; Chen, X.; Liu, J. Delivery Luteolin with Folacin-Modified Nanoparticle for Glioma Therapy. Int. J. Nanomed. 2019, 14, 7515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sabzichi, M.; Hamishehkar, H.; Ramezani, F.; Sharifi, S.; Tabasinezhad, M.; Pirouzpanah, M.; Ghanbari, P.; Samadi, N. Luteolin-Loaded Phytosomes Sensitize Human Breast Carcinoma MDA-MB 231 Cells to Doxorubicin by Suppressing Nrf2 Mediated Signalling. Asian Pac. J. Cancer Prev. 2014, 15, 5311–5316. [Google Scholar] [CrossRef] [Green Version]
- Kollur, S.P.; Prasad, S.K.; Pradeep, S.; Veerapur, R.; Patil, S.S.; Amachawadi, R.G.; Lamraoui, G.; Al-Kheraif, A.A.; Elgorban, A.M.; Syed, A. Luteolin-Fabricated ZnO Nanostructures Showed PLK-1 Mediated Anti-Breast Cancer Activity. Biomolecules 2021, 11, 385. [Google Scholar] [CrossRef]
- Altamimi, M.A.; Hussain, A.; AlRajhi, M.; Alshehri, S.; Imam, S.S.; Qamar, W. Luteolin-Loaded Elastic Liposomes for Transdermal Delivery to Control Breast Cancer: In Vitro and Ex Vivo Evaluations. Pharmaceuticals 2021, 14, 1143. [Google Scholar] [CrossRef]
- Ding, J.; Li, Q.; He, S.; Xie, J.; Liang, X.; Wu, T.; Li, D. Luteolin-Loading of Her-2-Poly (Lactic-Co-Glycolic Acid) Nanoparticles and Proliferative Inhibition of Gastric Cancer Cells via Targeted Regulation of Forkhead Box Protein O1. J. Cancer Res. Ther. 2020, 16, 263. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Çetinkaya, M.; Baran, Y. Therapeutic Potential of Luteolin on Cancer. Vaccines 2023, 11, 554. https://doi.org/10.3390/vaccines11030554
Çetinkaya M, Baran Y. Therapeutic Potential of Luteolin on Cancer. Vaccines. 2023; 11(3):554. https://doi.org/10.3390/vaccines11030554
Chicago/Turabian StyleÇetinkaya, Melisa, and Yusuf Baran. 2023. "Therapeutic Potential of Luteolin on Cancer" Vaccines 11, no. 3: 554. https://doi.org/10.3390/vaccines11030554
APA StyleÇetinkaya, M., & Baran, Y. (2023). Therapeutic Potential of Luteolin on Cancer. Vaccines, 11(3), 554. https://doi.org/10.3390/vaccines11030554