
The Development of Gut Microbiota and Its Changes Following C. jejuni Infection in Broilers


Abstract
:1. Introduction
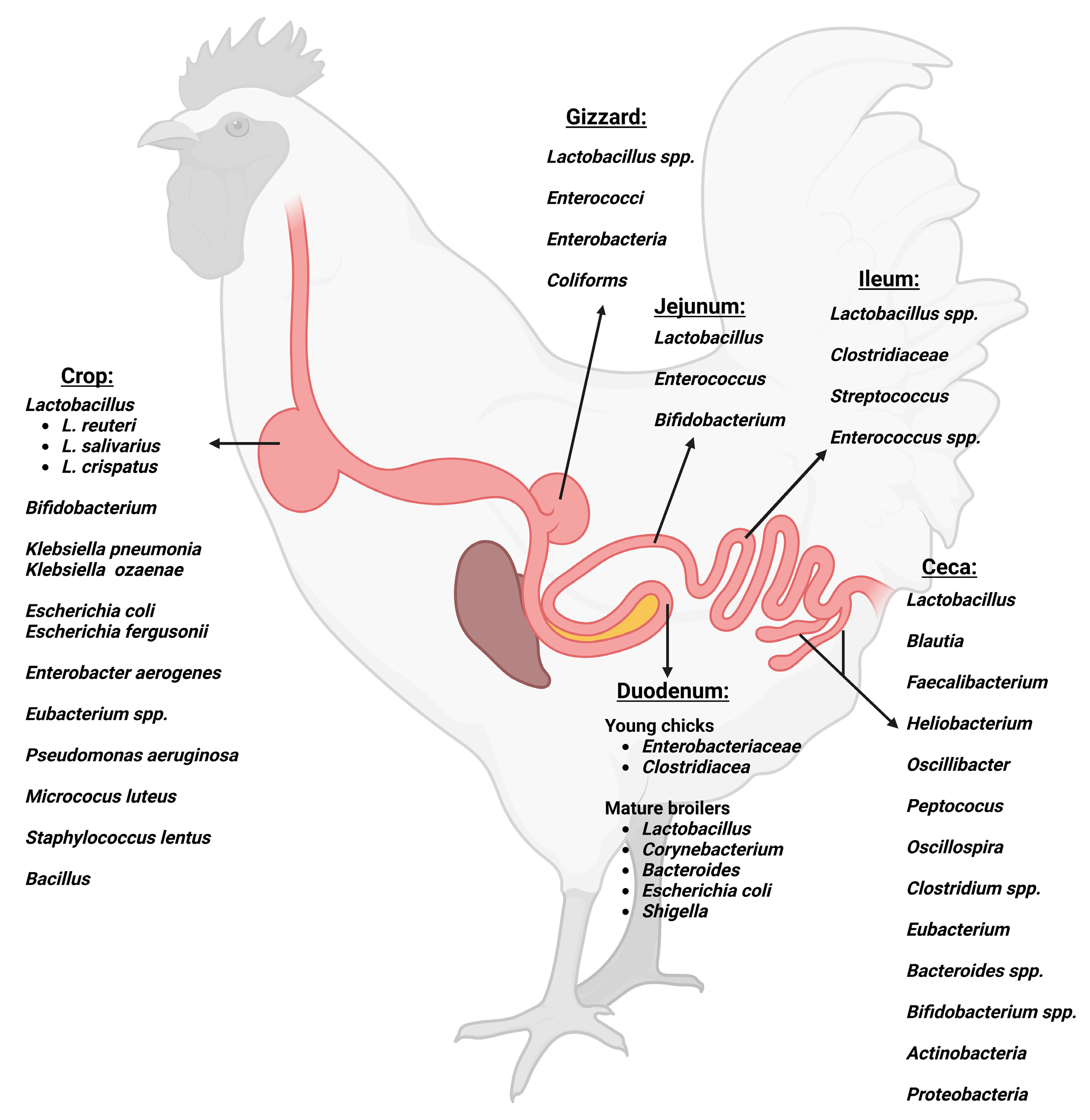
2. Gut Microbiota and Its Development in Commercial Broilers
2.1. Gastrointestinal Tract Microbial Composition in Commercial Broilers
2.1.1. Crop
2.1.2. Gizzard and Proventriculus
2.1.3. Small Intestine
Duodenum
Jejunum
Ileum
2.1.4. Ceca
3. The Impact of the Chicken Gut Microbiome on the Avian Immune System
3.1. The Chicken Gut Microbiome Impact on the Innate Immune System Components
3.1.1. Mucin Dynamics
3.1.2. Gene Expression and Macrophage-like Cells
3.1.3. Natural Killer Cells, Heterophils, and Defensins
3.2. The Chicken Gut Microbiome Impact on the Adaptative Immune System Components
3.2.1. B-Cells and Immunoglobulins
3.2.2. CD4+ and CD8+ T-Cells
3.2.3. T Regulatory Cells
3.2.4. Th17 Cells
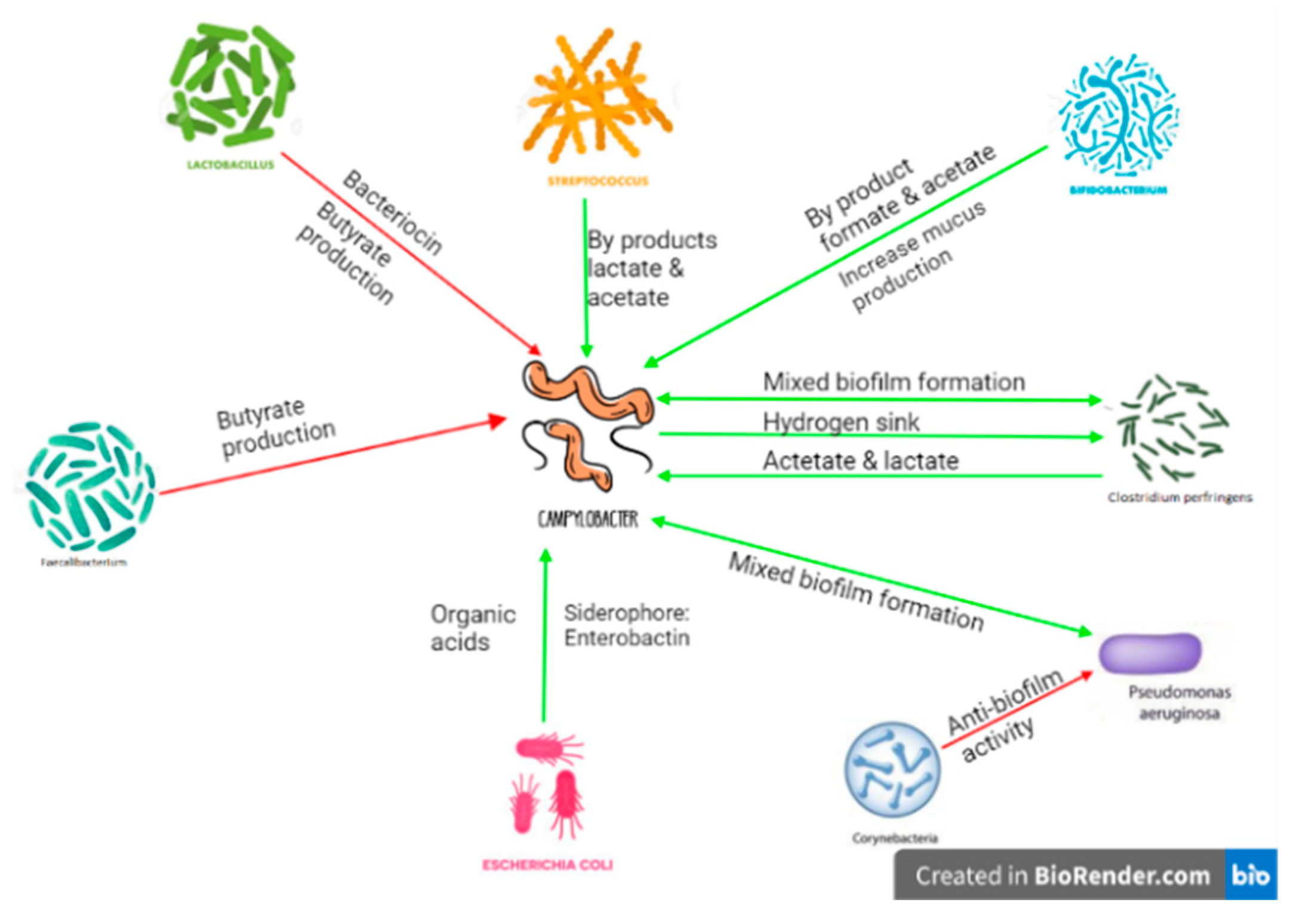
4. Campylobacter jejuni Modulation of the Avian Gut
5. Summary and Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Fathima, S.; Shanmugasundaram, R.; Adams, D.; Selvaraj, R. Gastrointestinal microbiota and their manipulation for improved growth and performance in chickens. Foods 2022, 11, 1401. [Google Scholar] [PubMed]
- Shang, Y.; Kumar, S.; Oakley, B.; Kim, W.K. Chicken gut microbiota: Importance and detection technology. Front. Vet. Sci. 2018, 5, 254. [Google Scholar] [PubMed] [Green Version]
- Amoroso, C.; Perillo, F.; Strati, F.; Fantini, M.; Caprioli, F.; Facciotti, F. The role of gut microbiota biomodulators on mucosal immunity and intestinal inflammation. Cells 2020, 9, 1234. [Google Scholar] [PubMed]
- Alexandratos, N.; Bruinsma, J. World Agriculture towards 2030/2050: The 2012 Revision; FAO: Rome, Italy, 2012. [Google Scholar]
- Al Hakeem, W.G.; Fathima, S.; Shanmugasundaram, R.; Selvaraj, R.K. Campylobacter jejuni in poultry: Pathogenesis and control strategies. Microorganisms 2022, 10, 2134. [Google Scholar]
- Price, K.R.; Hargis, B.M.; Barta, J.R. From the wild red jungle fowl to domesticated chickens: Modification of eimerian-microbiome-host interactions. World’s Poult. Sci. J. 2015, 71, 349–362. [Google Scholar] [CrossRef]
- Al-Nasser, A.; Al-Khalaifa, H.; Al-Saffar, A.; Khalil, F.; Albahouh, M.; Ragheb, G.; Al-Haddad, A.; Mashaly, M. Overview of chicken taxonomy and domestication. World’s Poult. Sci. J. 2007, 63, 285–300. [Google Scholar]
- Mignon-Grasteau, S.; Narcy, A.; Rideau, N.; Chantry-Darmon, C.; Boscher, M.-Y.; Sellier, N.; Chabault, M.; Konsak-Ilievski, B.; Le Bihan-Duval, E.; Gabriel, I. Impact of selection for digestive efficiency on microbiota composition in the chicken. PLoS ONE 2015, 10, e0135488. [Google Scholar]
- Ding, J.; Dai, R.; Yang, L.; He, C.; Xu, K.; Liu, S.; Zhao, W.; Xiao, L.; Luo, L.; Zhang, Y.; et al. Inheritance and establishment of gut microbiota in chickens. Front. Microbiol. 2017, 8, 1967. [Google Scholar]
- Lee, S.; La, T.-M.; Lee, H.-J.; Choi, I.-S.; Song, C.-S.; Park, S.-Y.; Lee, J.-B.; Lee, S.-W. Characterization of microbial communities in the chicken oviduct and the origin of chicken embryo gut microbiota. Sci. Rep. 2019, 9, 6838. [Google Scholar]
- Cisek, A.; Binek, M. Chicken intestinal microbiota function with a special emphasis on the role of probiotic bacteria. Pol. J. Vet. Sci. 2014, 17, 385–394. [Google Scholar]
- Maki, J.J.; Bobeck, E.A.; Sylte, M.J.; Looft, T. Eggshell and environmental bacteria contribute to the intestinal microbiota of growing chickens. J. Anim. Sci. Biotechnol. 2020, 11, 60. [Google Scholar] [CrossRef]
- Stanley, D.; Geier, M.S.; Hughes, R.J.; Denman, S.E.; Moore, R.J. Highly variable microbiota development in the chicken gastrointestinal tract. PLoS ONE 2014, 8, e84290. [Google Scholar] [CrossRef] [Green Version]
- Olsen, R.; Kudirkiene, E.; Thøfner, I.; Pors, S.; Karlskov-Mortensen, P.; Li, L.; Papasolomontos, S.; Angastiniotou, C.; Christensen, J. Impact of egg disinfection of hatching eggs on the eggshell microbiome and bacterial load. Poult. Sci. 2017, 96, 3901–3911. [Google Scholar] [CrossRef]
- Pedroso, A.; Menten, J.; Lambais, M. The structure of bacterial community in the intestines of Newly Hatched Chicks1. J. Appl. Poult. Res 2005, 14, 232–237. [Google Scholar] [CrossRef]
- Pan, D.; Yu, Z. Intestinal microbiome of poultry and its interaction with host and diet. Gut Microbes 2014, 5, 108–119. [Google Scholar] [CrossRef] [Green Version]
- Annett, C.; Viste, J.; Chirino-Trejo, M.; Classen, H.; Middleton, D.; Simko, E. Necrotic enteritis: Effect of barley, wheat and corn diets on proliferation of Clostridium perfringens type A. Avian Pathol. 2002, 31, 598–601. [Google Scholar] [CrossRef] [PubMed]
- Riddell, C.; Kong, X.-M. The influence of diet on necrotic enteritis in broiler chickens. Avian Dis. 1992, 36, 499–503. [Google Scholar] [CrossRef] [PubMed]
- Torok, V.A.; Hughes, R.J.; Ophel-Keller, K.; Ali, M.; MacAlpine, R. Influence of different litter materials on cecal microbiota colonization in broiler chickens. Poult. Sci. 2009, 88, 2474–2481. [Google Scholar] [CrossRef]
- Akerele, G.; Al Hakeem, W.G.; Lourenco, J.; Selvaraj, R.K. The effect of necrotic enteritis challenge on production performance, cecal microbiome, and cecal tonsil transcriptome in broilers. Pathogens 2022, 11, 839. [Google Scholar] [CrossRef]
- Macdonald, S.E.; Nolan, M.J.; Harman, K.; Boulton, K.; Hume, D.A.; Tomley, F.M.; Stabler, R.A.; Blake, D.P. Effects of Eimeria tenella infection on chicken caecal microbiome diversity, exploring variation associated with severity of pathology. PLoS ONE 2017, 12, e0184890. [Google Scholar] [CrossRef]
- Kubasova, T.; Kollarcikova, M.; Crhanova, M.; Karasova, D.; Cejkova, D.; Sebkova, A.; Matiasovicova, J.; Faldynova, M.; Pokorna, A.; Cizek, A.; et al. Contact with adult hen affects development of caecal microbiota in newly hatched chicks. PLoS ONE 2019, 14, e0212446. [Google Scholar] [CrossRef]
- Feye, K.M.; Baxter, M.F.A.; Tellez-Isaias, G.; Kogut, M.H.; Ricke, S.C. Influential factors on the composition of the conventionally raised broiler gastrointestinal microbiomes. Poult. Sci. 2020, 99, 653–659. [Google Scholar] [CrossRef] [PubMed]
- Hilmi, H.T.A.; Surakka, A.; Apajalahti, J.; Saris, P.E.J. Identification of the most abundant Lactobacillus species in the crop of 1- and 5-week-old broiler chickens. Appl. Env. Microbiol. 2007, 73, 7867–7873. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lev, M.; Briggs, C. The gut flora of the chick. II. The establishment of the flora. J. Appl. Bacteriol. 1956, 19, 224–230. [Google Scholar] [CrossRef]
- Rehman, H.U.; Vahjen, W.; Awad, W.A.; Zentek, J. Indigenous bacteria and bacterial metabolic products in the gastrointestinal tract of broiler chickens. Arch. Anim. Nutr. 2007, 61, 319–335. [Google Scholar] [CrossRef]
- Hinton, A., Jr.; Buhr, R.; Ingram, K. Physical, chemical, and microbiological changes in the crop of broiler chickens subjected to incremental feed withdrawal. Poult. Sci. 2000, 79, 212–218. [Google Scholar] [CrossRef] [PubMed]
- Svihus, B. The gizzard: Function, influence of diet structure and effects on nutrient availability. World’s Poult. Sci. J. 2011, 67, 207–224. [Google Scholar] [CrossRef]
- Gong, J.; Si, W.; Forster, R.J.; Huang, R.; Yu, H.; Yin, Y.; Yang, C.; Han, Y. 16S rRNA gene-based analysis of mucosa-associated bacterial community and phylogeny in the chicken gastrointestinal tracts: From crops to ceca. FEMS Microbiol. Ecol. 2007, 59, 147–157. [Google Scholar] [CrossRef] [Green Version]
- Engberg, R.; Hedemann, M.; Jensen, B. The influence of grinding and pelleting of feed on the microbial composition and activity in the digestive tract of broiler chickens. Br. Poult. Sci. 2002, 43, 569–579. [Google Scholar] [CrossRef]
- Xiao, Y.; Xiang, Y.; Zhou, W.; Chen, J.; Li, K.; Yang, H. Microbial community mapping in intestinal tract of broiler chicken. Poult. Sci. 2017, 96, 1387–1393. [Google Scholar] [CrossRef] [PubMed]
- Ballou, A.L.; Ali, R.A.; Mendoza, M.A.; Ellis, J.C.; Hassan, H.M.; Croom, W.J.; Koci, M.D. Development of the chick microbiome: How early exposure influences future microbial diversity. Front. Vet. Sci. 2016, 3, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jurburg, S.D.; Brouwer, M.S.M.; Ceccarelli, D.; van der Goot, J.; Jansman, A.J.M.; Bossers, A. Patterns of community assembly in the developing chicken microbiome reveal rapid primary succession. MicrobiologyOpen 2019, 8, e00821. [Google Scholar] [CrossRef]
- Lema, I.; Araújo, J.R.; Rolhion, N.; Demignot, S. Jejunum: The understudied meeting place of dietary lipids and the microbiota. Biochimie 2020, 178, 124–136. [Google Scholar] [CrossRef] [PubMed]
- Lavelle, A.; Sokol, H. Gut microbiota-derived metabolites as key actors in inflammatory bowel disease. Nat. Rev. Gastroenterol. Hepatol. 2020, 17, 223–237. [Google Scholar] [CrossRef]
- Yang, W.-Y.; Lee, Y.; Lu, H.; Chou, C.-H.; Wang, C. Analysis of gut microbiota and the effect of lauric acid against necrotic enteritis in Clostridium perfringens and Eimeria side-by-side challenge model. PLoS ONE 2019, 14, e0205784. [Google Scholar] [CrossRef] [Green Version]
- De Boever, P.; Wouters, R.; Verschaeve, L.; Berckmans, P.; Schoeters, G.; Verstraete, W. Protective effect of the bile salt hydrolase-active Lactobacillus reuteri against bile salt cytotoxicity. Appl. Microbiol. Biotechnol. 2000, 53, 709–714. [Google Scholar] [CrossRef] [PubMed]
- Portune, K.J.; Benítez-Páez, A.; Del Pulgar, E.M.G.; Cerrudo, V.; Sanz, Y. Gut microbiota, diet, and obesity-related disorders—The good, the bad, and the future challenges. Mol. Nutr. Food Res. 2017, 61, 1600252. [Google Scholar] [CrossRef] [Green Version]
- Lu, J.; Idris, U.; Harmon, B.; Hofacre, C.; Maurer, J.J.; Lee, M.D. Diversity and succession of the intestinal bacterial community of the maturing broiler chicken. Appl. Environ. Microbiol. 2003, 69, 6816–6824. [Google Scholar] [CrossRef] [Green Version]
- Shazali, N.; Loh, T.C.; Foo, H.L.; Samsudin, A.A. Gut microflora and intestinal morphology changes of broiler chickens fed reducing dietary protein supplemented with lysine, methionine, and threonine in tropical environment. Rev. Bras. De Zootec. 2019, 48, 1–15. [Google Scholar] [CrossRef]
- Oakley, B.B.; Lillehoj, H.S.; Kogut, M.H.; Kim, W.K.; Maurer, J.J.; Pedroso, A.; Lee, M.D.; Collett, S.R.; Johnson, T.J.; Cox, N.A. The chicken gastrointestinal microbiome. FEMS Microbiol. Lett. 2014, 360, 100–112. [Google Scholar] [CrossRef]
- Józefiak, D.; Rutkowski, A.; Martin, S. Carbohydrate fermentation in the avian ceca: A review. Anim. Feed. Sci. Technol. 2004, 113, 1–15. [Google Scholar] [CrossRef]
- Lee, K.-C.; Kil, D.Y.; Sul, W.J. Cecal microbiome divergence of broiler chickens by sex and body weight. J. Microbiol. 2017, 55, 939–945. [Google Scholar] [CrossRef] [PubMed]
- Ocejo, M.; Oporto, B.; Hurtado, A. 16S rRNA amplicon sequencing characterization of caecal microbiome composition of broilers and free-range slow-growing chickens throughout their productive lifespan. Sci. Rep. 2019, 9, 2506. [Google Scholar] [CrossRef] [Green Version]
- Gilbert, J.A.; Blaser, M.J.; Caporaso, J.G.; Jansson, J.K.; Lynch, S.V.; Knight, R. Current understanding of the human microbiome. Nat. Med. 2018, 24, 392–400. [Google Scholar] [CrossRef]
- Jovel, J.; Dieleman, L.A.; Kao, D.; Mason, A.L.; Wine, E. The human gut microbiome in health and disease. Metagenomics 2018, 13, 197–213. [Google Scholar]
- Miani, M.; Le Naour, J.; Waeckel-Enée, E.; chand Verma, S.; Straube, M.; Emond, P.; Ryffel, B.; Van Endert, P.; Sokol, H.; Diana, J. Gut microbiota-stimulated innate lymphoid cells support β-defensin 14 expression in pancreatic endocrine cells, preventing autoimmune diabetes. Cell Metab. 2018, 28, 557–572.e556. [Google Scholar] [CrossRef] [Green Version]
- Vaishnava, S.; Behrendt, C.L.; Ismail, A.S.; Eckmann, L.; Hooper, L.V. Paneth cells directly sense gut commensals and maintain homeostasis at the intestinal host-microbial interface. Proc. Natl. Acad. Sci. USA 2008, 105, 20858–20863. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Menendez, A.; Willing, B.; Montero, M.; Wlodarska, M.; So, C.; Bhinder, G.; Vallance, B.; Finlay, B. Bacterial stimulation of the TLR-MyD88 pathway modulates the homeostatic expression of ileal Paneth cell α-defensins. J. Innate Immun. 2013, 5, 39–49. [Google Scholar] [CrossRef] [PubMed]
- Franchi, L.; Kamada, N.; Nakamura, Y.; Burberry, A.; Kuffa, P.; Suzuki, S.; Shaw, M.H.; Kim, Y.-G.; Núñez, G. NLRC4-driven production of IL-1β discriminates between pathogenic and commensal bacteria and promotes host intestinal defense. Nat. Immunol. 2012, 13, 449–456. [Google Scholar] [CrossRef]
- Levy, M.; Thaiss, C.A.; Katz, M.N.; Suez, J.; Elinav, E. Inflammasomes and the Microbiota—Partners in the Preservation of Mucosal Homeostasis. Semin. Immunopathol. 2015, 37, 39–46. [Google Scholar]
- Nielsen, M.M.; Witherden, D.A.; Havran, W.L. γδ T cells in homeostasis and host defence of epithelial barrier tissues. Nat. Rev. Immunol. 2017, 17, 733–745. [Google Scholar] [CrossRef] [Green Version]
- Ueda, Y.; Kayama, H.; Jeon, S.G.; Kusu, T.; Isaka, Y.; Rakugi, H.; Yamamoto, M.; Takeda, K. Commensal microbiota induce LPS hyporesponsiveness in colonic macrophages via the production of IL-10. Int. Immunol. 2010, 22, 953–962. [Google Scholar] [CrossRef] [Green Version]
- Acevedo-Villanueva, K.Y.; Akerele, G.O.; Al Hakeem, W.G.; Renu, S.; Shanmugasundaram, R.; Selvaraj, R.K. A novel approach against Salmonella: A review of polymeric nanoparticle vaccines for broilers and layers. Vaccines 2021, 9, 1041. [Google Scholar] [CrossRef] [PubMed]
- Belova, A.V.; Smutka, L.; Rosochatecká, E. World chicken meat market–its development and current status. Acta Univ. Agric. Et Silvic. Mendel. Brun. 2012, 60, 15–30. [Google Scholar] [CrossRef] [Green Version]
- Cheled-Shoval, S.; Gamage, N.W.; Amit-Romach, E.; Forder, R.; Marshal, J.; Van Kessel, A.; Uni, Z. Differences in intestinal mucin dynamics between germ-free and conventionally reared chickens after mannan-oligosaccharide supplementation. Poult. Sci. 2014, 93, 636–644. [Google Scholar] [CrossRef] [PubMed]
- Turck, D.; Feste, A.S.; Lifschitz, C.H. Age and diet affect the composition of porcine colonic mucins. Pediatr. Res. 1993, 33, 564–567. [Google Scholar] [CrossRef] [Green Version]
- Forder, R.; Howarth, G.; Tivey, D.; Hughes, R. Bacterial modulation of small intestinal goblet cells and mucin composition during early posthatch development of poultry. Poult. Sci. 2007, 86, 2396–2403. [Google Scholar] [CrossRef]
- Smirnov, A.; Perez, R.; Amit-Romach, E.; Sklan, D.; Uni, Z. Mucin dynamics and microbial populations in chicken small intestine are changed by dietary probiotic and antibiotic growth promoter supplementation. J. Nutr. 2005, 135, 187–192. [Google Scholar] [CrossRef] [Green Version]
- Schokker, D.; Jansman, A.J.; Veninga, G.; de Bruin, N.; Vastenhouw, S.A.; de Bree, F.M.; Bossers, A.; Rebel, J.M.; Smits, M.A. Perturbation of microbiota in one-day old broiler chickens with antibiotic for 24 hours negatively affects intestinal immune development. BMC Genom. 2017, 18, 241. [Google Scholar] [CrossRef] [Green Version]
- Abbas, A.K.; Lichtman, A.H.; Pillai, S.; Baker, D.; Baker, A. Cellular and Molecular Immunology; Elsevier: Philadelphia, PA, USA, 2015; Volume 199. [Google Scholar]
- Meijerink, N.; Kers, J.G.; Velkers, F.C.; Van Haarlem, D.A.; Lamot, D.M.; De Oliveira, J.E.; Smidt, H.; Stegeman, J.A.; Rutten, V.P.; Jansen, C.A. Early life inoculation with adult-derived microbiota accelerates maturation of intestinal microbiota and enhances NK cell activation in broiler chickens. Front. Vet. Sci. 2020, 7, 584561. [Google Scholar] [CrossRef] [PubMed]
- Davison, F.; Kaspers, B.; Schat, K.A. Avian Immunology; Elsevier: Philadelphia, PA, USA, 2008. [Google Scholar]
- Harmon, B.G. Avian heterophils in inflammation and disease resistance. Poult. Sci. 1998, 77, 972–977. [Google Scholar] [CrossRef] [PubMed]
- Farnell, M.; Donoghue, A.; De Los Santos, F.S.; Blore, P.; Hargis, B.; Tellez, G.; Donoghue, D. Upregulation of oxidative burst and degranulation in chicken heterophils stimulated with probiotic bacteria. Poult. Sci. 2006, 85, 1900–1906. [Google Scholar] [CrossRef]
- Vos, J.B.; Datson, N.A.; Rabe, K.F.; Hiemstra, P.S. Exploring host-pathogen interactions at the epithelial surface: Application of transcriptomics in lung biology. Am. J. Physiol. -Lung Cell. Mol. Physiol. 2006, 292, L367–L377. [Google Scholar] [CrossRef]
- van Dijk, A.; Veldhuizen, E.J.; Haagsman, H.P. Avian defensins. Vet. Immunol. Immunopathol. 2008, 124, 1–18. [Google Scholar] [CrossRef]
- Butler, V.L.; Mowbray, C.A.; Cadwell, K.; Niranji, S.S.; Bailey, R.; Watson, K.A.; Ralph, J.; Hall, J. Effects of rearing environment on the gut antimicrobial responses of two broiler chicken lines. Vet. Immunol. Immunopathol. 2016, 178, 29–36. [Google Scholar] [CrossRef]
- Zhao, Q.; Elson, C.O. Adaptive immune education by gut microbiota antigens. Immunology 2018, 154, 28–37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, J.H.; Wang, K.-W.; Zhang, D.; Zhan, X.; Wang, T.; Bu, C.-H.; Behrendt, C.L.; Zeng, M.; Wang, Y.; Misawa, T. IgD class switching is initiated by microbiota and limited to mucosa-associated lymphoid tissue in mice. Proc. Natl. Acad. Sci. USA 2017, 114, E1196–E1204. [Google Scholar] [CrossRef] [Green Version]
- Woof, J.; Russell, M. Structure and function relationships in IgA. Mucosal Immunol. 2011, 4, 590–597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaspers, B.; Lettmann, S.; Roell, S. Development of the Gut Associated Immune System. In Proceedings of the 20th European Symposium on Poultry Nutrition, Prague, Czech Republic, 24–27 August 2015; p. 12. [Google Scholar]
- Zenner, C.; Hitch, T.C.; Riedel, T.; Wortmann, E.; Tiede, S.; Buhl, E.M.; Abt, B.; Neuhaus, K.; Velge, P.; Overmann, J. Early-life immune system maturation in chickens using a synthetic community of cultured gut bacteria. Msystems 2021, 6, e01300–e01320. [Google Scholar] [CrossRef]
- Vandaveer, S.; Erf, G.; Durdik, J. Avian T helper one/two immune response balance can be shifted toward inflammation by antigen delivery to scavenger receptors. Poult. Sci. 2001, 80, 172–181. [Google Scholar] [CrossRef] [PubMed]
- Shanmugasundaram, R.; Selvaraj, R.K. Regulatory T cell properties of chicken CD4+ CD25+ cells. J. Immunol. 2011, 186, 1997–2002. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, I.K.; Gu, M.J.; Ko, K.H.; Bae, S.; Kim, G.; Jin, G.-D.; Kim, E.B.; Kong, Y.-Y.; Park, T.S.; Park, B.-C. Regulation of CD4+ CD8− CD25+ and CD4+ CD8+ CD25+ T cells by gut microbiota in chicken. Sci. Rep. 2018, 8, 8627. [Google Scholar] [CrossRef] [Green Version]
- Atarashi, K.; Tanoue, T.; Shima, T.; Imaoka, A.; Kuwahara, T.; Momose, Y.; Cheng, G.; Yamasaki, S.; Saito, T.; Ohba, Y. Induction of colonic regulatory T cells by indigenous Clostridium species. Science 2011, 331, 337–341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Josefowicz, S.Z.; Niec, R.E.; Kim, H.Y.; Treuting, P.; Chinen, T.; Zheng, Y.; Umetsu, D.T.; Rudensky, A.Y. Extrathymically generated regulatory T cells control mucosal TH2 inflammation. Nature 2012, 482, 395–399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, W.H.; Chaudhari, A.A.; Lillehoj, H.S. Involvement of T cell immunity in avian coccidiosis. Front. Immunol. 2019, 10, 2732. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, W.H.; Lillehoj, H.S.; Min, W. Indole treatment alleviates intestinal tissue damage induced by chicken coccidiosis through activation of the aryl hydrocarbon receptor. Front. Immunol. 2019, 10, 560. [Google Scholar] [CrossRef]
- Reid, W.D.K.; Close, A.J.; Humphrey, S.; Chaloner, G.; Lacharme-Lora, L.; Rothwell, L.; Kaiser, P.; Williams, N.J.; Humphrey, T.J.; Wigley, P.; et al. Cytokine responses in birds challenged with the human food-borne pathogen Campylobacter jejuni implies a Th17 response. R. Soc. Open Sci. 2016, 3, 150541. [Google Scholar] [CrossRef] [Green Version]
- Ivanov, I.I.; Atarashi, K.; Manel, N.; Brodie, E.L.; Shima, T.; Karaoz, U.; Wei, D.; Goldfarb, K.C.; Santee, C.A.; Lynch, S.V.; et al. Induction of intestinal Th17 cells by segmented filamentous bacteria. Cell 2009, 139, 485–498. [Google Scholar] [CrossRef] [Green Version]
- Skirrow, M.B. Diseases due to Campylobacter, Helicobacter and related bacteria. J. Comp. Pathol. 1994, 111, 113–149. [Google Scholar] [CrossRef]
- Awad, W.A.; Hess, C.; Hess, M. Re-thinking the chicken–Campylobacter jejuni interaction: A review. Avian Pathol. 2018, 47, 352–363. [Google Scholar] [CrossRef]
- Cason, E.E.; Al Hakeem, W.G.; Adams, D.; Shanmugasundaram, R.; Selvaraj, R. Effects of synbiotic supplementation as an antibiotic growth promoter replacement on cecal Campylobacter jejuni load in broilers challenged with C. jejuni. J. Appl. Poult. Res. 2022, 32, 100315. [Google Scholar] [CrossRef]
- Kaakoush, N.O.; Castaño-Rodríguez, N.; Mitchell, H.M.; Man, S.M. Global epidemiology of Campylobacter infection. Clin. Microbiol. Rev. 2015, 28, 687–720. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Acevedo-Villanueva, K.; Akerele, G.; Al-Hakeem, W.; Adams, D.; Gourapura, R.; Selvaraj, R. Immunization of broiler chickens with a killed chitosan nanoparticle Salmonella vaccine decreases Salmonella enterica serovar enteritidis load. Front. Physiol. 2022, 13, 920777. [Google Scholar] [CrossRef] [PubMed]
- Sahin, O.; Kassem, I.I.; Shen, Z.; Lin, J.; Rajashekara, G.; Zhang, Q. Campylobacter in poultry: Ecology and potential interventions. Avian Dis. 2015, 59, 185–200. [Google Scholar] [CrossRef] [PubMed]
- Skarp, C.; Hänninen, M.-L.; Rautelin, H. Campylobacteriosis: The role of poultry meat. Clin. Microbiol. Infect. 2016, 22, 103–109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wagenaar, J.A.; French, N.P.; Havelaar, A.H. Preventing Campylobacter at the source: Why is it so difficult? Clin. Infect. Dis. 2013, 57, 1600–1606. [Google Scholar] [CrossRef] [Green Version]
- Pielsticker, C.; Glünder, G.; Rautenschlein, S. Colonization properties of Campylobacter jejuni in chickens. Eur. J. Microbiol. Immunol. 2012, 2, 61–65. [Google Scholar] [CrossRef] [Green Version]
- Parkhill, J.; Wren, B.W.; Mungall, K.; Ketley, J.M.; Churcher, C.; Basham, D.; Chillingworth, T.; Davies, R.M.; Feltwell, T.; Holroyd, S.; et al. The genome sequence of the food-borne pathogen Campylobacter jejuni reveals hypervariable sequences. Nature 2000, 403, 665–668. [Google Scholar] [CrossRef] [Green Version]
- Wright, J.A.; Grant, A.J.; Hurd, D.; Harrison, M.; Guccione, E.J.; Kelly, D.J.; Maskell, D.J. Metabolite and transcriptome analysis of Campylobacter jejuni in vitro growth reveals a stationary-phase physiological switch. Microbiology 2009, 155, 80–94. [Google Scholar] [CrossRef] [Green Version]
- Thomas, M.T.; Shepherd, M.; Poole, R.K.; van Vliet, A.H.M.; Kelly, D.J.; Pearson, B.M. Two respiratory enzyme systems in Campylobacter jejuni NCTC 11168 contribute to growth on l-lactate. Environ. Microbiol. 2011, 13, 48–61. [Google Scholar] [CrossRef]
- Stahl, M.; Butcher, J.; Stintzi, A. Nutrient acquisition and metabolism by Campylobacter jejuni. Front. Cell. Infect. Microbiol. 2012, 2, 5. [Google Scholar] [CrossRef] [Green Version]
- Awad, W.A.; Mann, E.; Dzieciol, M.; Hess, C.; Schmitz-Esser, S.; Wagner, M.; Hess, M. Age-related differences in the luminal and mucosa-associated gut microbiome of broiler chickens and shifts associated with Campylobacter jejuni infection. Front. Cell. Infect. Microbiol. 2016, 6, 154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Louis, P.; Flint, H.J. Diversity, metabolism and microbial ecology of butyrate-producing bacteria from the human large intestine. FEMS Microbiol. Lett. 2009, 294, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Parnell, J.; Reimer, R.A. Prebiotic fiber modulation of the gut microbiota improves risk factors for obesity and the metabolic syndrome. Gut Microbes 2012, 3, 29–34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Q.; Lan, F.; Li, X.; Yan, W.; Sun, C.; Li, J.; Yang, N.; Wen, C. The spatial and temporal characterization of gut microbiota in broilers. Front. Vet. Sci. 2021, 8, 712226. [Google Scholar] [CrossRef]
- Awad, W.A.; Dublecz, F.; Hess, C.; Dublecz, K.; Khayal, B.; Aschenbach, J.R.; Hess, M. Campylobacter jejuni colonization promotes the translocation of Escherichia coli to extra-intestinal organs and disturbs the short-chain fatty acids profiles in the chicken gut. Poult. Sci. 2016, 95, 2259–2265. [Google Scholar] [CrossRef]
- Kaakoush, N.O.; Sodhi, N.; Chenu, J.W.; Cox, J.M.; Riordan, S.M.; Mitchell, H.M. The interplay between Campylobacter and Helicobacter species and other gastrointestinal microbiota of commercial broiler chickens. Gut Pathog. 2014, 6, 18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belenguer, A.; Holtrop, G.; Duncan, S.H.; Anderson, S.E.; Calder, A.G.; Flint, H.J.; Lobley, G.E. Rates of production and utilization of lactate by microbial communities from the human colon. FEMS Microbiol. Ecol. 2011, 77, 107–119. [Google Scholar] [CrossRef] [Green Version]
- Connerton, P.L.; Richards, P.J.; Lafontaine, G.M.; O’Kane, P.M.; Ghaffar, N.; Cummings, N.J.; Smith, D.L.; Fish, N.M.; Connerton, I.F. The effect of the timing of exposure to Campylobacter jejuni on the gut microbiome and inflammatory responses of broiler chickens. Microbiome 2018, 6, 88. [Google Scholar] [CrossRef]
- Dempsey, E.; Corr, S.C. Lactobacillus spp. for gastrointestinal health: Current and future perspectives. Front. Immunol. 2022, 13, 840245. [Google Scholar] [CrossRef] [PubMed]
- Mortada, M.; Cosby, D.E.; Shanmugasundaram, R.; Selvaraj, R.K. In vivo and in vitro assessment of commercial probiotic and organic acid feed additives in broilers challenged with Campylobacter coli. J. Appl. Poult. Res. 2020, 29, 435–446. [Google Scholar] [CrossRef]
- Thibodeau, A.; Fravalo, P.; Yergeau, É.; Arsenault, J.; Lahaye, L.; Letellier, A. Chicken caecal microbiome modifications induced by Campylobacter jejuni colonization and by a non-antibiotic feed additive. PLoS ONE 2015, 10, e0131978. [Google Scholar] [CrossRef] [Green Version]
- Fathima, S.; Hakeem, W.G.A.; Shanmugasundaram, R.; Selvaraj, R.K. Necrotic enteritis in broiler chickens: A review on the pathogen, pathogenesis, and prevention. Microorganisms 2022, 10, 1958. [Google Scholar] [CrossRef]
- Molnár, A.; Hess, C.; Pál, L.; Wágner, L.; Awad, W.; Husvéth, F.; Hess, M.; Dublecz, K. Composition of diet modifies colonization dynamics of Campylobacter jejuni in broiler chickens. J. Appl. Microbiol. 2015, 118, 245–254. [Google Scholar] [CrossRef] [PubMed]
- Indikova, I.; Humphrey, T.J.; Hilbert, F. Survival with a Helping Hand: Campylobacter and Microbiota. Front. Microbiol. 2015, 6, 1266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ica, T.; Caner, V.; Istanbullu, O.; Nguyen, H.D.; Ahmed, B.; Call, D.R.; Beyenal, H. Characterization of mono-and mixed-culture Campylobacter jejuni biofilms. Appl. Environ. Microbiol. 2012, 78, 1033–1038. [Google Scholar]
- Hakansson, A.P.; Orihuela, C.J.; Bogaert, D. Bacterial-host interactions: Physiology and pathophysiology of respiratory infection. Physiol. Rev. 2018, 98, 781–811. [Google Scholar] [CrossRef] [Green Version]
- Dalili, D.; Amini, M.; Faramarzi, M.A.; Fazeli, M.R.; Khoshayand, M.R.; Samadi, N. Isolation and structural characterization of Coryxin, a novel cyclic lipopeptide from Corynebacterium xerosis NS5 having emulsifying and anti-biofilm activity. Colloids Surf. B Biointerfaces 2015, 135, 425–432. [Google Scholar] [CrossRef]
- Menberu, M.A.; Liu, S.; Cooksley, C.; Hayes, A.J.; Psaltis, A.J.; Wormald, P.-J.; Vreugde, S. Corynebacterium accolens has antimicrobial activity against Staphylococcus aureus and methicillin-resistant S. aureus pathogens isolated from the sinonasal niche of chronic rhinosinusitis patients. Pathogens 2021, 10, 207. [Google Scholar] [PubMed]
- Fernandez-Julia, P.; Commane, D.M.; van Sinderen, D.; Munoz-Munoz, J. Cross-feeding interactions between human gut commensals belonging to the Bacteroides and Bifidobacterium genera when grown on dietary glycans. Microbiome Res. Rep. 2022, 1, 12. [Google Scholar] [CrossRef]
- Van Deun, K.; Pasmans, F.; Van Immerseel, F.; Ducatelle, R.; Haesebrouck, F. Butyrate protects Caco-2 cells from Campylobacter jejuni invasion and translocation. Br. J. Nutr. 2008, 100, 480–484. [Google Scholar]
- O’Callaghan, A.; van Sinderen, D. Bifidobacteria, and Their Role as Members of the Human Gut Microbiota. Front. Microbiol. 2016, 7, 925. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruas-Madiedo, P.; Gueimonde, M.; Fernández-García, M.; de los Reyes-Gavilán, C.G.; Margolles, A. Mucin degradation by Bifidobacterium strains isolated from the human intestinal microbiota. Appl. Environ. Microbiol. 2008, 74, 1936–1940. [Google Scholar] [CrossRef] [Green Version]
- Muraoka, W.T.; Zhang, Q. Phenotypic and genotypic evidence for L-fucose utilization by Campylobacter jejuni. J. Bacteriol. 2011, 193, 1065–1075. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stahl, M.; Friis, L.M.; Nothaft, H.; Liu, X.; Li, J.; Szymanski, C.M.; Stintzi, A. L-fucose utilization provides Campylobacter jejuni with a competitive advantage. Proc. Natl. Acad. Sci. USA 2011, 108, 7194–7199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopez-Siles, M.; Duncan, S.H.; Garcia-Gil, L.J.; Martinez-Medina, M. Faecalibacterium prausnitzii: From microbiology to diagnostics and prognostics. ISME J. 2017, 11, 841–852. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Innate Immune System | Reference | |
| Mucosal components |
| [56] |
| [58] | |
| [59] | |
| Gene expression and Macrophage-like cells |
| [60] |
| Natural Killer Cells |
| [62] |
| Heterophils |
| [65] |
| Defensins |
| [68] |
| Adaptative Immune System | Reference | |
| B-cells and Immunoglobulins |
| [72] |
| [30,73] | |
| CD4+ and CD8+ T-cells |
| [72] |
| [60] | |
| [62] | |
| T-regulatory Cells |
| [76] |
| Th-17 Cells |
| [80] |
| [81] | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Al Hakeem, W.G.; Acevedo Villanueva, K.Y.; Selvaraj, R.K. The Development of Gut Microbiota and Its Changes Following C. jejuni Infection in Broilers. Vaccines 2023, 11, 595. https://doi.org/10.3390/vaccines11030595
Al Hakeem WG, Acevedo Villanueva KY, Selvaraj RK. The Development of Gut Microbiota and Its Changes Following C. jejuni Infection in Broilers. Vaccines. 2023; 11(3):595. https://doi.org/10.3390/vaccines11030595
Chicago/Turabian StyleAl Hakeem, Walid Ghazi, Keila Y. Acevedo Villanueva, and Ramesh K. Selvaraj. 2023. "The Development of Gut Microbiota and Its Changes Following C. jejuni Infection in Broilers" Vaccines 11, no. 3: 595. https://doi.org/10.3390/vaccines11030595
APA StyleAl Hakeem, W. G., Acevedo Villanueva, K. Y., & Selvaraj, R. K. (2023). The Development of Gut Microbiota and Its Changes Following C. jejuni Infection in Broilers. Vaccines, 11(3), 595. https://doi.org/10.3390/vaccines11030595