
Long-Term Cross Immune Response in Mice following Heterologous Prime-Boost COVID-19 Vaccination with Full-Length Spike mRNA and Recombinant S1 Protein
 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cells and Virus
2.2. SARS-CoV-2 Protein Vaccine and mRNA Vaccine
2.3. Animal Ethical Approval
2.4. Animal Immune Study
2.5. Neutralization Assay for Wild-Type and Variant Strains of SARS-CoV-2
2.6. ELISA Assay for Wild-Type and Variant Strains of SARS-CoV-2
2.7. ELISpot Assays for Wild-Type and Variant Strains of SARS-CoV-2
2.8. Statistical Analysis
3. Results
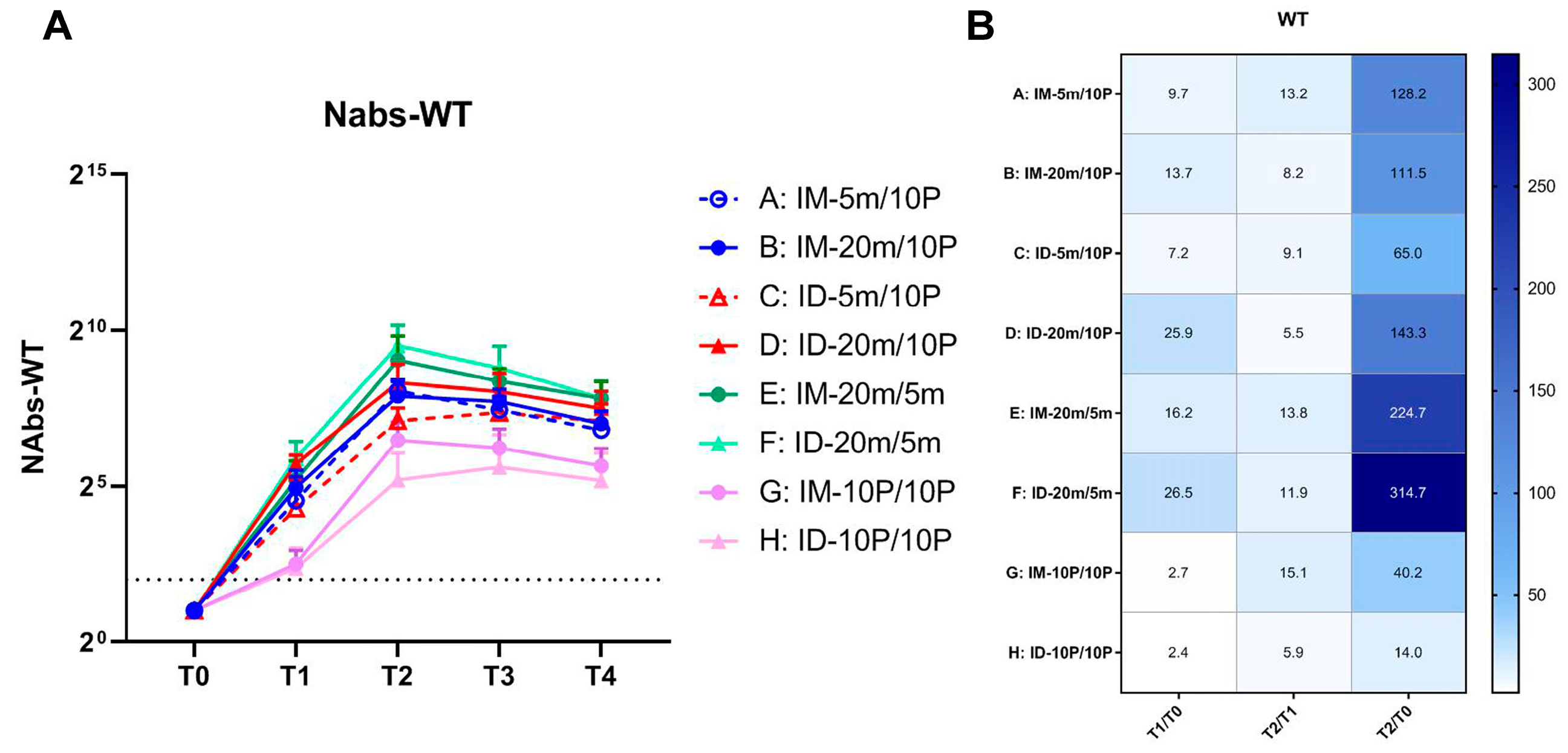
3.1. Neutralization of SARS-CoV-2 WT Strain by Sera from S mRNA/S1 Protein Heterologous Immunization
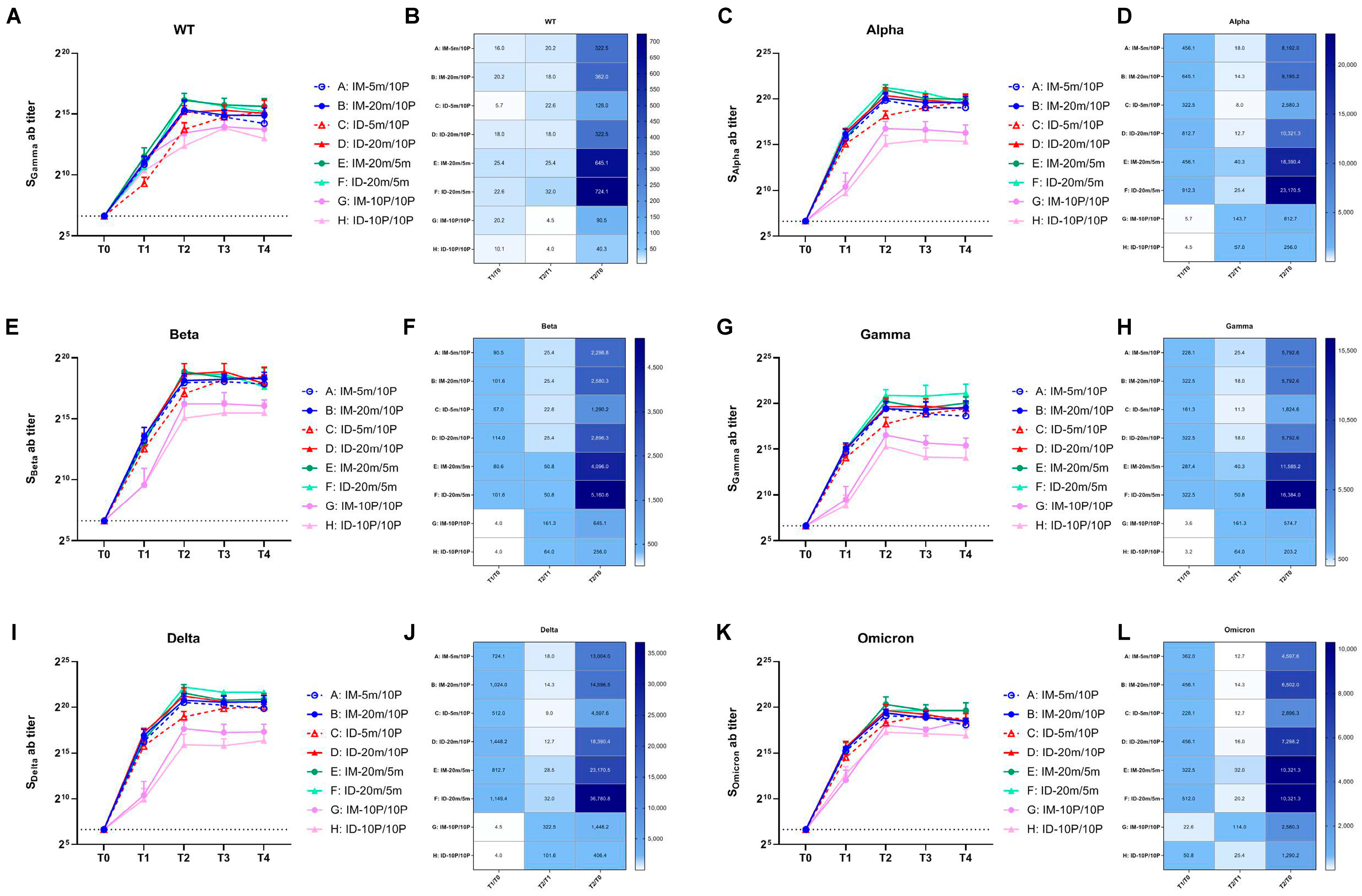
3.2. Cross-Reactive Antibodies Targeting VOCs Promoted via Boosting with Recombinant S1 Protein
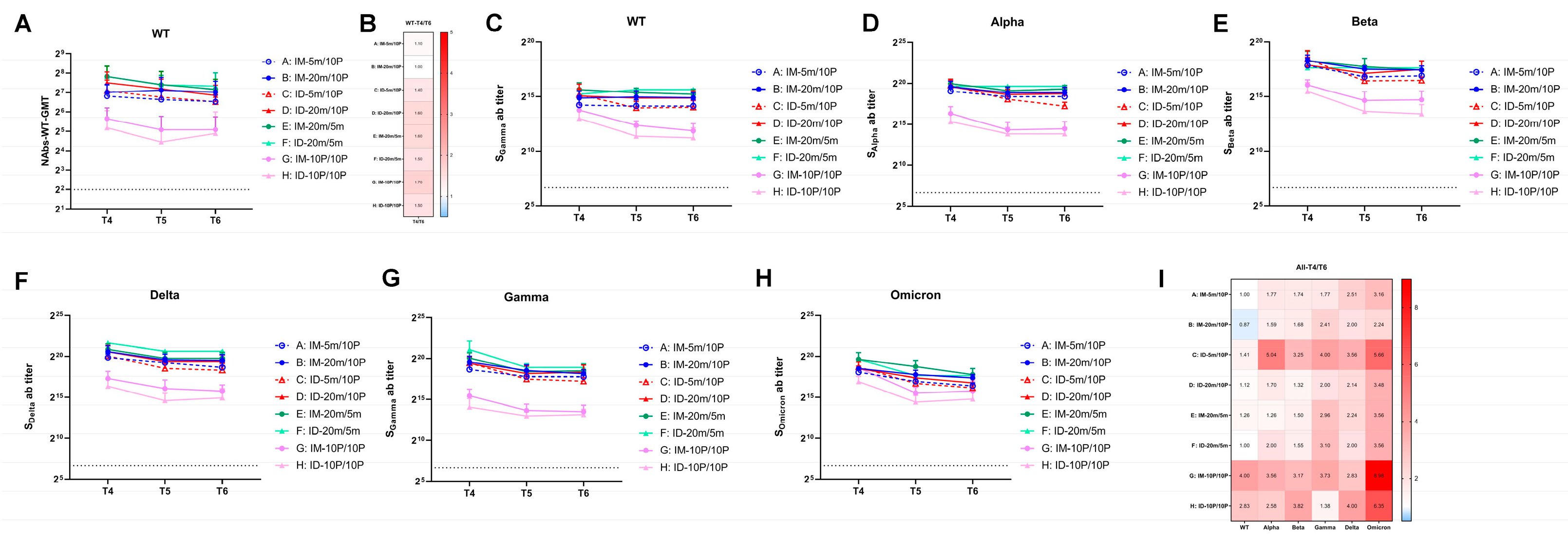
3.3. Over Seven-Month, Long-Term Antibody Response Induced by S mRNA/S1 Protein Heterologous Immunization
3.4. Over Seven-Month, Long-Term Cellular Immune Responses Induced by S mRNA/S1 Protein Heterologous Vaccine
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- WHO. Coronavirus Disease (COVID-19) Dashboard 2023; WHO: Geneva, Switzerland, 2023. [Google Scholar]
- Chiu, N.C.; Chi, H.; Tu, Y.K.; Huang, Y.N.; Tai, Y.L.; Weng, S.L.; Chang, L.; Huang, D.T.-N.; Huang, F.-Y.; Lin, C.-Y. To mix or not to mix? A rapid systematic review of heterologous prime-boost COVID-19 vaccination. Expert Rev. Vaccines 2021, 20, 1211–1220. [Google Scholar] [PubMed]
- He, Q.; Mao, Q.; An, C.; Zhang, J.; Gao, F.; Bian, L.; Li, C.; Liang, Z.; Xu, M.; Wang, J. Heterologous prime-boost: Breaking the protective immune response bottleneck of COVID-19 vaccine candidates. Emerg. Microbes Infect. 2021, 10, 629–637. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.T.; Quach, T.H.T.; Tran, T.M.; Phuoc, H.N.; Nguyen, H.T.; Vo, T.K.; Vo, G.V. Reactogenicity and immunogenicity of heterologous prime-boost immunization with COVID-19 vaccine. Biomed. Pharmacother. 2022, 147, 112650. [Google Scholar] [CrossRef] [PubMed]
- Shaw, R.H.; Stuart, A.; Greenland, M.; Liu, X.; Nguyen Van-Tam, J.S.; Snape, M.D.; Com-COV Study Group. Heterologous prime-boost COVID-19 vaccination: Initial reactogenicity data. Lancet 2021, 397, 2043–2046. [Google Scholar] [CrossRef]
- Fan, S.; Liao, Y.; Lian, Y.; Jiang, G.; Jiang, L.; Dong, C.; Yang, E.; Wang, L.; Xu, X.; Feng, M.; et al. Role of innate lymphoid cells and dendritic cells in intradermal immunization of the enterovirus antigen. NPJ Vaccines 2019, 4, 14. [Google Scholar] [CrossRef]
- Fan, S.; Liao, Y.; Jiang, G.; Jiang, L.; Wang, L.; Xu, X.; Feng, M.; Yang, E.; Zhang, Y.; Cui, W.; et al. Study of integrated protective immunity induced in rhesus macaques by the intradermal administration of a bivalent EV71-CA16 inactivated vaccine. Vaccine 2020, 38, 2034–2044. [Google Scholar] [CrossRef]
- Fan, S.; Liao, Y.; Jiang, G.; Wang, L.; Zhao, H.; Yu, L.; Xu, X.; Li, D.; Zhang, Y.; Li, Q.; et al. Efficacy of an inactivated bivalent vaccine for enterovirus 71 and coxsackievirus A16 in mice immunized intradermally. Vaccine 2021, 39, 596–604. [Google Scholar] [CrossRef]
- Anand, A.; Zaman, K.; Estivariz, C.F.; Yunus, M.; Gary, H.E.; Weldon, W.C.; Bari, T.; Oberste, S.; Wassilak, S.; Luby, S.; et al. Early priming with inactivated poliovirus vaccine (IPV) and intradermal fractional dose IPV administered by a microneedle device: A randomized controlled trial. Vaccine 2015, 33, 6816–6822. [Google Scholar] [CrossRef]
- Fan, S.; Xiao, K.; Li, D.; Zhao, H.; Zhang, J.; Yu, L.; Chang, P.; Zhu, S.; Xu, X.; Liao, Y.; et al. Preclinical immunological evaluation of an intradermal heterologous vaccine against SARS-CoV-2 variants. Emerg. Microbes Infect. 2022, 11, 212–226. [Google Scholar] [CrossRef]
- Fan, S.; Li, D.; Zhao, H.; Yu, L.; Cui, P.; Wang, L.; Zhang, Y.; Liao, Y.; Xu, X.; Jiang, G.; et al. Ameliorated immunity elicited by intradermal inoculation in individuals vaccinated with inactivated SARS-CoV-2 vaccine. Vaccine 2021, 39, 6980–6983. [Google Scholar] [CrossRef]
- Pu, J.; Yu, Q.; Yin, Z.; Zhang, Y.; Li, X.; Yin, Q.; Chen, H.; Long, R.; Zhao, Z.; Mou, T.; et al. The safety and immunogenicity of an inactivated SARS-CoV-2 vaccine in Chinese adults aged 18–59 years: A phase I randomized, double-blinded, controlled trial. Vaccine 2021, 39, 2746–2754. [Google Scholar] [CrossRef] [PubMed]
- Li, R. Transient transfection of CHO cells using linear polyethylenimine is a simple and effective means of producing rainbow trout recombinant IFN-gamma protein. Cytotechnology 2015, 67, 987–993. [Google Scholar] [CrossRef] [PubMed]
- Xie, T.; Lu, S.; He, Z.; Liu, H.; Wang, J.; Tang, C.; Yang, T.; Yu, W.; Li, H.; Yang, Y.; et al. Three doses of prototypic SARS-CoV-2 inactivated vaccine induce cross-protection against its variants of concern. Signal Transduct. Target. Ther. 2022, 7, 61. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Liao, Y.; Jiang, G.; Yao, W.; Duan, S.; Xiao, K.; Ding, X.; Zhao, H.; Zhang, Y.; Zhang, A.; et al. Immunological evaluation of an mRNA vaccine booster in individuals fully immunized with an inactivated SARS-CoV-2 vaccine. Clin. Transl. Med. 2022, 12, e875. [Google Scholar] [CrossRef] [PubMed]
- Gagne, M.; Moliva, J.I.; Foulds, K.E.; Andrew, S.F.; Flynn, B.J.; Werner, A.P.; Wagner, D.; Teng, I.-T.; Lin, B.; Moore, C.; et al. mRNA-1273 or mRNA-Omicron boost in vaccinated macaques elicits similar B cell expansion, neutralizing responses, and protection from Omicron. Cell 2022, 185, 1556–1571.e18. [Google Scholar] [CrossRef] [PubMed]
- Vanshylla, K.; Di Cristanziano, V.; Kleipass, F.; Dewald, F.; Schommers, P.; Gieselmann, L.; Gruell, H.; Schlotz, M.; Ercanoglu, M.; Stumpf, R.; et al. Kinetics and correlates of the neutralizing antibody response to SARS-CoV-2 infection in humans. Cell Host Microbe 2021, 29, 917–929.e14. [Google Scholar] [CrossRef]
- Dan, J.M.; Mateus, J.; Kato, Y.; Hastie, K.M.; Yu, E.D.; Faliti, C.E.; Grifoni, A.; Ramirez, S.; Haupt, S.; Frazier, A.; et al. Immunological memory to SARS-CoV-2 assessed for up to 8 months after infection. Science 2021, 371, eabf4063. [Google Scholar] [CrossRef]
- Sokal, A.; Chappert, P.; Barba-Spaeth, G.; Roeser, A.; Fourati, S.; Azzaoui, I.; Vandenberghe, A.; Fernandez, I.; Meola, A.; Bouvier-Alias, M.; et al. Maturation and persistence of the anti-SARS-CoV-2 memory B cell response. Cell 2021, 184, 1201–1213.e14. [Google Scholar] [CrossRef]
- Sepand, M.R.; Bigdelou, B.; Ho, J.Q.; Sharaf, M.; Lannigan, A.J.; Sullivan, I.M.; da Silva, A.; Barrett, L.; McGoldrick, S.; Lnu, Y.; et al. Long-Term Immunity and Antibody Response: Challenges for Developing Efficient COVID-19 Vaccines. Antibodies 2022, 11, 35. [Google Scholar] [CrossRef]
- Doria-Rose, N.; Suthar, M.S.; Makowski, M.; O’Connell, S.; McDermott, A.B.; Flach, B.; Ledgerwood, J.; Mascola, J.; Graham, B.; Lin, B.; et al. Antibody Persistence through 6 Months after the Second Dose of mRNA-1273 Vaccine for Covid-19. N. Engl. J. Med. 2021, 384, 2259–2261. [Google Scholar] [CrossRef]
- Lee, I.J.; Sun, C.P.; Wu, P.Y.; Lan, Y.H.; Wang, I.H.; Liu, W.C.; Yuan, J.P.-Y.; Chang, Y.-W.; Tseng, S.-C.; Tsung, S.-I.; et al. A booster dose of Delta x Omicron hybrid mRNA vaccine produced broadly neutralizing antibody against Omicron and other SARS-CoV-2 variants. J. Biomed. Sci. 2022, 29, 49. [Google Scholar] [CrossRef] [PubMed]
- Tarke, A.; Sidney, J.; Methot, N.; Yu, E.D.; Zhang, Y.; Dan, J.M.; Goodwin, B.; Rubiro, P.; Sutherland, A.; Wang, E.; et al. Impact of SARS-CoV-2 variants on the total CD4(+) and CD8(+) T cell reactivity in infected or vaccinated individuals. Cell Rep. Med. 2021, 2, 100355. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.J.; Kim, D.U.; Noh, J.Y.; Kim, S.; Park, S.H.; Jeong, H.W.; Shin, E.-C. T cell epitopes in SARS-CoV-2 proteins are substantially conserved in the Omicron variant. Cell Mol. Immunol. 2022, 19, 447–448. [Google Scholar] [CrossRef]
- van Doremalen, N.; Fischer, R.J.; Schulz, J.E.; Holbrook, M.G.; Smith, B.J.; Lovaglio, J.; Petsch, B.; Munster, V.J. Immunogenicity of Low-Dose Prime-Boost Vaccination of mRNA Vaccine CV07050101 in Non-Human Primates. Viruses 2021, 13, 1645. [Google Scholar] [CrossRef] [PubMed]
- Mateus, J.; Dan, J.M.; Zhang, Z.; Rydyznski Moderbacher, C.; Lammers, M.; Goodwin, B.; Sette, A.; Crotty, S.; Weiskopf, D. Low-dose mRNA-1273 COVID-19 vaccine generates durable memory enhanced by cross-reactive T cells. Science 2021, 374, eabj9853. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; He, Q.; Yan, X.; Liu, J.; Bai, Y.; An, C.; Cui, B.; Gao, F.; Mao, Q.; Wang, J.; et al. Mixed formulation of mRNA and protein-based COVID-19 vaccines triggered superior neutralizing antibody responses. MedComm 2022, 3, e188. [Google Scholar] [CrossRef]
- Levin, Y.; Kochba, E.; Hung, I.; Kenney, R. Intradermal vaccination using the novel microneedle device MicronJet600: Past, present, and future. Hum. Vaccin. Immunother. 2015, 11, 991–997. [Google Scholar] [CrossRef]
- Hung, I.F.; Levin, Y.; To, K.K.; Chan, K.H.; Zhang, A.J.; Li, P.; Li, C.; Xu, T.; Wong, T.-Y.; Yuen, K.-Y. Dose sparing intradermal trivalent influenza (2010/2011) vaccination overcomes reduced immunogenicity of the 2009 H1N1 strain. Vaccine 2012, 30, 6427–6435. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, D.; Zhao, H.; Liao, Y.; Jiang, G.; Cui, P.; Zhang, Y.; Yu, L.; Fan, S.; Li, H.; Li, Q. Long-Term Cross Immune Response in Mice following Heterologous Prime-Boost COVID-19 Vaccination with Full-Length Spike mRNA and Recombinant S1 Protein. Vaccines 2023, 11, 963. https://doi.org/10.3390/vaccines11050963
Li D, Zhao H, Liao Y, Jiang G, Cui P, Zhang Y, Yu L, Fan S, Li H, Li Q. Long-Term Cross Immune Response in Mice following Heterologous Prime-Boost COVID-19 Vaccination with Full-Length Spike mRNA and Recombinant S1 Protein. Vaccines. 2023; 11(5):963. https://doi.org/10.3390/vaccines11050963
Chicago/Turabian StyleLi, Dandan, Heng Zhao, Yun Liao, Guorun Jiang, Pingfang Cui, Ying Zhang, Li Yu, Shengtao Fan, Hangwen Li, and Qihan Li. 2023. "Long-Term Cross Immune Response in Mice following Heterologous Prime-Boost COVID-19 Vaccination with Full-Length Spike mRNA and Recombinant S1 Protein" Vaccines 11, no. 5: 963. https://doi.org/10.3390/vaccines11050963
APA StyleLi, D., Zhao, H., Liao, Y., Jiang, G., Cui, P., Zhang, Y., Yu, L., Fan, S., Li, H., & Li, Q. (2023). Long-Term Cross Immune Response in Mice following Heterologous Prime-Boost COVID-19 Vaccination with Full-Length Spike mRNA and Recombinant S1 Protein. Vaccines, 11(5), 963. https://doi.org/10.3390/vaccines11050963