1. Introduction
Bordetella bronchiseptica (Bb) is a Gram-negative coccobacillus of the genus Bordetella, which encompasses 16 other species, including
Bordetella pertussis.
B. bronchiseptica, can infect a wide range of mammals, including dogs, cats, rodents, and pigs [
1]. In dogs, Bb is the major causative agent of canine infectious respiratory disease complex (CIRDC) or kennel cough, while other pathogens such as
Mycoplasma cynos and canine parainfluenza virus (CPIV) are also frequently detected [
2]. Nevertheless, Bb was frequently identified as the causal agent of CIRDC, as seen in 3.3% to 78.7% of canine samples in Europe [
3]. CIRDC is characterized by tracheobronchitis, manifesting as a cough, lethargy, and intranasal discharge, and on rare occasions, it can lead to [
4] or lethal bronchopneumonia in puppies [
5]. Moreover, transmission to the owner is possible [
6,
7]. Bb has also been implicated in respiratory infections in immunocompromised humans [
8], and occasionally even in immunocompetent humans [
9]. In dogs, the current standard of care is based on treatment with antibiotics and supportive care for clinical signs. However, the attachment of the bacteria to the cilia may necessitate the nebulization of the antibiotics [
10]. Vaccines are an essential tool for the protection of animals and humans against this pathogen.
Several vaccines with various routes of administration (oral, intranasal, or injectable) are currently licensed for clinical use in dogs [
11]. The mucosal vaccines contain live attenuated strains, whereas the injectable vaccines consist of inactivated bacteria or antigen extracts [
11]. A number of studies have compared the efficacy of the vaccines licensed for dogs and the immune responses they induce [
12,
13,
14,
15,
16]. The parameters studied are generally limited to the observation of clinical signs (cough, fever, sneezing, and lethargy), colonization and bacterial shedding, and the induction of hemagglutinating antibodies in the serum. However, some studies have also considered mucosal immune responses, such as the induction of Bp-specific IgA [
15]. Mucosal vaccination against Bb via the oral or intranasal route has been reported to lead to better protection than the use of injectable vaccines does [
15,
16]. Nevertheless, it is not possible to conclude that the route of administration is solely responsible for these observed differences, because the formulation of the vaccines used also differs. However, in comparison to vaccination via the intranasal and oral routes, the superiority of one route over the other remains a matter of debate [
14]. The intranasal route more closely resembles the natural route of infection, but oral vaccines are easier to administer and can also induce mucosal immune responses. Notably, Larson et al. showed that intranasal and oral vaccines were similarly effective against Bb in dogs, both being superior to the inactivated parenteral vaccine [
16]. Another study comparing oral and intranasal vaccination reported no significant differences in protection following vaccination via these two routes [
12]. Moreover, vaccination via either of these routes can lead to more than 12 months of protection in dogs [
17,
18].
The main weakness of all the studies cited above is their lack of consideration of cellular responses, the colonization of other organs and the induction of immunological memory in the vaccinated animals. There is a critical need to develop a model that can enable an improved understanding of the mechanisms underlying the efficacy of each vaccine. This should facilitate the future development of new vaccines in dogs, for which an understanding of the best way to efficiently induce mucosal immune responses will be key. Novel candidates targeting both
B. bronchiseptica and
B. pertussis, thereby protecting both humans and animals, have been studied [
19,
20]. A relevant model for preclinical validation is required for the evaluation of these vaccines. Interestingly, the infection of rats with these bacterial species reproduces the lead clinical sign of CIRDC in dogs, the cough [
21]. For this reason, the rat model has been used to study the pathogenesis of Bordetella infections and responses to vaccination against these pathogens for more than a century [
21,
22,
23,
24,
25].
In this study, we used a rat model to investigate and compare established vaccine strains against a
B. bronchiseptica challenge infection dosed through various routes. We previously studied the pathogenesis and induction of disease in Wistar rats after challenge with Bb, as previously described [
21]. Here, a live attenuated vaccine strain was used. The vaccine strain was administered orally or intranasally for immunization of the animals, which were subsequently challenged. Numerous end points, including, cough, colonization, lesions, and humoral and cellular immune responses between groups were evaluated. Importantly, we observed that vaccine administration via either the oral or intranasal route prevented colonization and induced specific local and systemic antibody responses. However, cellular immune responses in the spleen and clinical signs differed between routes of administration. Overall, the observed data suggest that mucosal vaccination effectively protects rats against a Bb challenge infection. Such detailed understanding of the specific immunological and clinical outcomes influenced by mucosal routes of vaccination in translatable animal models have important implications in the field of vaccinology and infectious disease research.
2. Materials and Methods
2.1. Vaccine Strain, Growth, and Vaccination
The live attenuated strain of Bb present in the oral Recombitek vaccine [
17] was used for vaccination. The vaccine strain was grown in TSB-ST (BioMérieux SA, Marcy l’étoile, France) at 37 °C, with shaking at 180 rpm, for 14 h. The bacterium was cultured until an OD
600 of 2.5 (approximate equivalent to 1.5 × 10
9 CFU/mL) was reached. At D0 and D21, female Wistar rats (3–4 weeks old) were anesthetized with isoflurane for nasal administration. The live attenuated vaccine strain was then administered intranasally (10
6 CFU/animal) or orally (10
8 CFU/animal) with a micropipette. The bacterium was diluted in phosphate-buffered saline (PBS); the inoculation volume used was 50 µL/animal for intranasal administration (25 µL in each nostril), and 100 µL for oral micropipette-guided drug administration (MDA) [
26]. One group of animals received PBS under the same conditions, as a non-vaccinated control.
2.2. B. bronchiseptica Strain, Growth, and Challenge
A virulent Bb strain of canine origin was used for challenge. This challenge strain was grown in TSB-ST (BioMérieux SA, Marcy l’étoile, France) at 37 °C, with shaking at 180 rpm, for 6 h. The bacterial culture was collected when an OD600 of 1 was reached (approximately equivalent to 1 × 109 colony-forming units (CFU)/mL). On D35, all the study animals were anesthetized with isoflurane. Rats were inoculated with 25 µL of the challenge strain in each nostril, resulting in a total dose of 50 µL of the strain in PBS, or a titer of 1 × 103 CFU/animal. The diluted bacterial cultures were back-titrated to confirm the titers used (CFU/animal). Each suspension was serially diluted and 10 µL drops were dispensed onto Bordet Gengou agar containing 15% sheep blood (Becton Dickinson, Heidelberg, Germany).
2.3. Vaccine-Specific Antibody Detection via ELISA
Antibody titers were determined using an enzyme-linked immunosorbent assay (ELISA) to evaluate the levels of Bb-specific antibodies in the serum, intranasal washes, and feces of vaccinated and challenged rats. Blood was sampled via the intracardiac route into BD Microtainer SST Tube (Becton Dickinson, Heidelberg, Germany) and incubated for 30 min at room temperature. All blood samples were centrifuged at 10,000× g for 10 min at 4 °C. The supernatants were harvested and stored at −20 °C until analysis. Feces were added to an Eppendorf tube, with 1 mL of PBS and a 1% 1× protease inhibitor cocktail (Thermo Scientific, Rockford, IL, USA) per 100 mg of the sample, and shaken until a homogeneous suspension was obtained. Intranasal washes were sampled after the animals had been euthanized, by flushing 400 µL of PBS through the trachea and collecting it via the nares. Fifty microliters of each intranasal sample was added to a DNase- and RNase-free tube, which was then stored at −80 °C until analysis. We added 35 µL of a 10× protease inhibitor cocktail to the remaining volume. Fecal and intranasal samples were centrifuged at 10,000× g for 10 min at 4 °C, and the supernatants were harvested and stored at −20 °C until analysis.
Bb vaccine strain-specific ELISA plates were coated with 50 µL/well of a 5 µg/mL suspension of the sonicated live attenuated Bb strain. The coated plates were incubated overnight at 4 °C, washed three times with 200 µL per well of 1× PBS-0.05% Tween 20 and blocked via incubation with 1× PBS- 5% BSA (5% bovine serum albumin in PBS) for 1 h at 37 °C. The plates were washed again as described above, and the sera was diluted in a range of 1:1000 to 1:20,000 in 1× PBS for sera, while for intranasal washes and feces, a dilution of 1:2 to 1:4 was used. Fifty microliters of the resulting solutions were added per well. The plates were incubated for 2 h at 37 °C and then washed three times with 200 µL of 1× PBS-0,05% Tween 20. They were incubated with 50 µL/well of anti-rat IgG and IgM (1:6000 dilution) and IgA (1:10,000 dilution) in 1× PBS-BSA 5% (Bio-Rad Laboratories, Inc., Hercules, CA, USA) for detection. The plates were incubated for 1 h at 37 °C and washed three times, as previously described. In the next step 50 µL of 3,3′, 5,5”-tetramethylbenzidine (TMB)/well was added and the plates were incubated for 15 min in the dark at room temperature. The reaction was stopped by adding 50 µL of hydrochloric acid (HCl)/well. The titers of rat IgG, IgM, and IgA antibodies were determined on a TECAN microplate reader (Tecan Austria GmbH, Grödig, Austria), via a colorimetric evaluation of the stopped reactions on the ELISA plate at a wavelength of 450 nm.
2.4. Detection Challenge and Vaccine Strain
Bacterial load was determined via specific quantitative real time polymerase chain reactions (qPCR) and culture on blood agar plates. On D-1, 13 days post-boost and 7 and 14 days post-challenge (p.c.), 4 rats per group were euthanized with pentobarbital and their tracheas and lungs were removed, weighed, and minced in 5 mL of Dulbecco phosphate-buffered saline (D-PBS). Samples were homogenized with a syringe piston and passed through a cell strainer with 70 µm pores. The cell strainer was washed with D-PBS, and the suspension was collected and centrifuged at 6000×
g for 5 min at 22 °C. At this step, 200 µL of each sample was added to DNase- and RNase-free tubes for qPCR and frozen at −80 °C until analysis. The tissue suspension obtained was serially diluted in D-PBS and plated on blood agar plates. The bacteria were cultured at 37 °C for 2 days, and the number of CFU was determined. The number of viable colonies were counted manually, and the presence/absence of hemolysis was used to differentiate visually between the vaccine (non-hemolytic) and challenge (hemolytic) strains (
Supplementary Materials, Figure S1). The identity of the colonies corresponding to Bb was confirmed by mass spectrometry using MALDI-TOF (BRUKER, Billerica, MA, USA).
Analysis was performed to differentiate between the Bb-challenge or Bb-vaccine strain, employing strain specific qPCR using primers specifically targeting sequences of either the Bb-challenge or the Bb -vaccine strain. Frozen tracheal and lung homogenates and nasal lavages were thawed. DNA was extracted with DNeasy Blood and Tissue Kit (QIAGEN GmbH, Hilden, Germany) in accordance with the manufacturer’s protocol. DNA quantity and quality were assessed with NanoDrop 2000c (Thermo Fisher Scientific, Waltham, MA, USA). Reactions were then set up, in duplicate, on a 96-well qPCR plate. The reaction mixture consisted of 5 µL of the DNA-containing samples, 12.5 µL of 1× Master Mix Quantitect Multiplex PCR (QIAGEN GmbH, Hilden, Germany), 0.5 µL of each primer (listed in
Table 1), and 0.25 µL of a probe and RNAse-free water to obtain the total volume of 20 µL. The reaction mixture was subjected to an initial denaturation step at 95 °C for 15 min, followed by 40 cycles of 94 °C for 60 s, and then 60 °C (vaccine strain) or 64.5 °C (challenge strain) for 60 s.
2.5. Analysis of Cough
The clinical signs of three animals per group were monitored at three different timepoints: 6, 9, and 12 days post-challenge. Each animal was filmed separately, for 10 min, with 4K Video Camera VX980 (Panasonic, Osaka, Japan) and the noise was monitored with an MKE 600 microphone (Sennheiser, Wedemark, Germany). The number of coughs was analyzed with Adobe Premier Pro (Adobe Systems Inc., San José, CA, USA), based on the recorded images and sound waveforms.
2.6. Histology Trachea and Lung
For the histological analysis, 2 cm of each lung and trachea was dissected after challenge and added them to 3 mL of 1× PBS supplemented with 4% paraformaldehyde (PFA) (SIGMA-ALDRICH, St. Louis, MO, USA). These samples were sent to Vet Diagnostics (VetDiag, Lyon, France) for microscopical analysis. Sliced samples were stained with hematoxylin-eosin (H-E) in accordance with standard procedures. Microscopic lesions and/or the presence of bacteria were analyzed under a Nikon Eclipse Ci light microscope (Nikon Europe B.V.). The analysis was performed by an external certified veterinary pathologist who had no knowledge of the group allocation. The scoring system used included categories such as absence of lesions, minimal, mild, or moderate, based on the level of inflammation characterized by cellular infiltration, epithelial alteration, degeneration, and the presence of lymphoid follicles.
2.7. Isolation of Splenocytes and Flow Cytometry Analysis
Spleens were harvested before challenge, 7 and 14 days post-challenge. They were homogenized mechanically and passed through a Nylon Corning® cell strainer with 100 µm pores (Corning Incorporated, Durham, NC, USA). The cell suspension was layered into Ficoll-Paque™ PREMIUM (Cytiva, Uppsala, Sweden) and centrifuged at 400× g at 10 min at 20 °C without the brake on. Cells were harvested and washed twice with 1× PBS. Part of the splenocytes were stained with trypan blue and counted by eye under a microscope (XYZ). The cells were frozen in a solution containing 50% RPMI, 40% fetal bovine serum (FBS) (Eurobio scientific, Les Ulis, France), and 10% dimethyl sulfoxide (DMSO) (SIGMA-ALDRICH, St. Louis, MO, USA) and stored in liquid nitrogen until analysis.
The cells were thawed in a 37 °C water bath, transferred to tubes containing a medium supplemented with 10% RPMI, 5% FBS, and 1× Antibiotic Antimycotic Solution (SIGMA-ALDRICH, St. Louis, MO, USA) and centrifuged at 400×
g for 10 min at 20 °C. Splenocytes were counted, viability was assessed after trypan blue staining, and cells were washed twice as described above. The cells were then activated via incubation with Cell Activation Cocktail (with Brefeldin A) (BioLegend, San Diego, CA, USA) for 5.5 h at 37 °C. The activated splenocytes were stained for viability and incubated for 25 min in the dark. The cells were then washed twice as described above, and surface-stained via incubation with the antibodies indicated in
Table 2 for 20 min. Cells were washed as described above and fixed via incubation with a 1× FOXP3 Fix/Perm solution (Thermo Fisher Scientific, Waltham, MA, USA) for 30 min in the dark. Samples were washed twice with Permeabilization Buffer (Thermo Fisher Scientific, Waltham, MA, USA) and incubated with the indicated diluted intracellular antibodies for 30 min. The cells were then washed twice as described above with Permeabilization Buffer and fixed with 1% paraformaldehyde diluted in 1× PBS. The flow cytometry data obtained were analyzed with FlowJo version 10.8.1 (FlowJo, Becton, Dickinson and Company, NJ, USA).
2.8. Statistics
Statistical analyses were performed with GraphPad Prism version 9 (GraphPad, San Diego, CA, USA). Differences between groups were evaluated using one-way ANOVA with Tukey tests for multiple comparisons and using two-way ANOVA for the comparison of multiple variables. There were three biological replicates for the cough analysis, and four replicates for all other analyses. Correlograms were generated with R Studio Software Version 4.0.2 (RStudio, Boston, MA, USA), and the multiple R-squared was calculated for each group of variables.
2.9. Animal Care and Use
All rats were housed in identical conditions at the university hospital unit for animal testing (Saint-Etienne, France). The experimental protocol was designed in accordance with French law (decree number 2001-464 29/05/01) and the recommendations of the European Economic Community (86/609/CEE) for the care and use of laboratory animals (permit no. #23498-2020010715103375).
4. Discussion
B. bronchiseptica is the main causal agent of CIRDC in dogs, and several vaccines against this bacterium are available on the market [
11]. However, limited knowledge about the causality of the efficacy of these vaccines has been published. A bridging animal model in rats was investigated as a possible tool for exploring and studying the underlying mechanisms. To this end, the mucosal administration of a live attenuated
B. bronchiseptica vaccine strain for dogs was investigated in Wistar rats. The over-arching goals for this study were (1) to explore the differences in the immune responses elicited via either oral or intranasal routes in rats and (2) to investigate aspects not previously described in dogs.
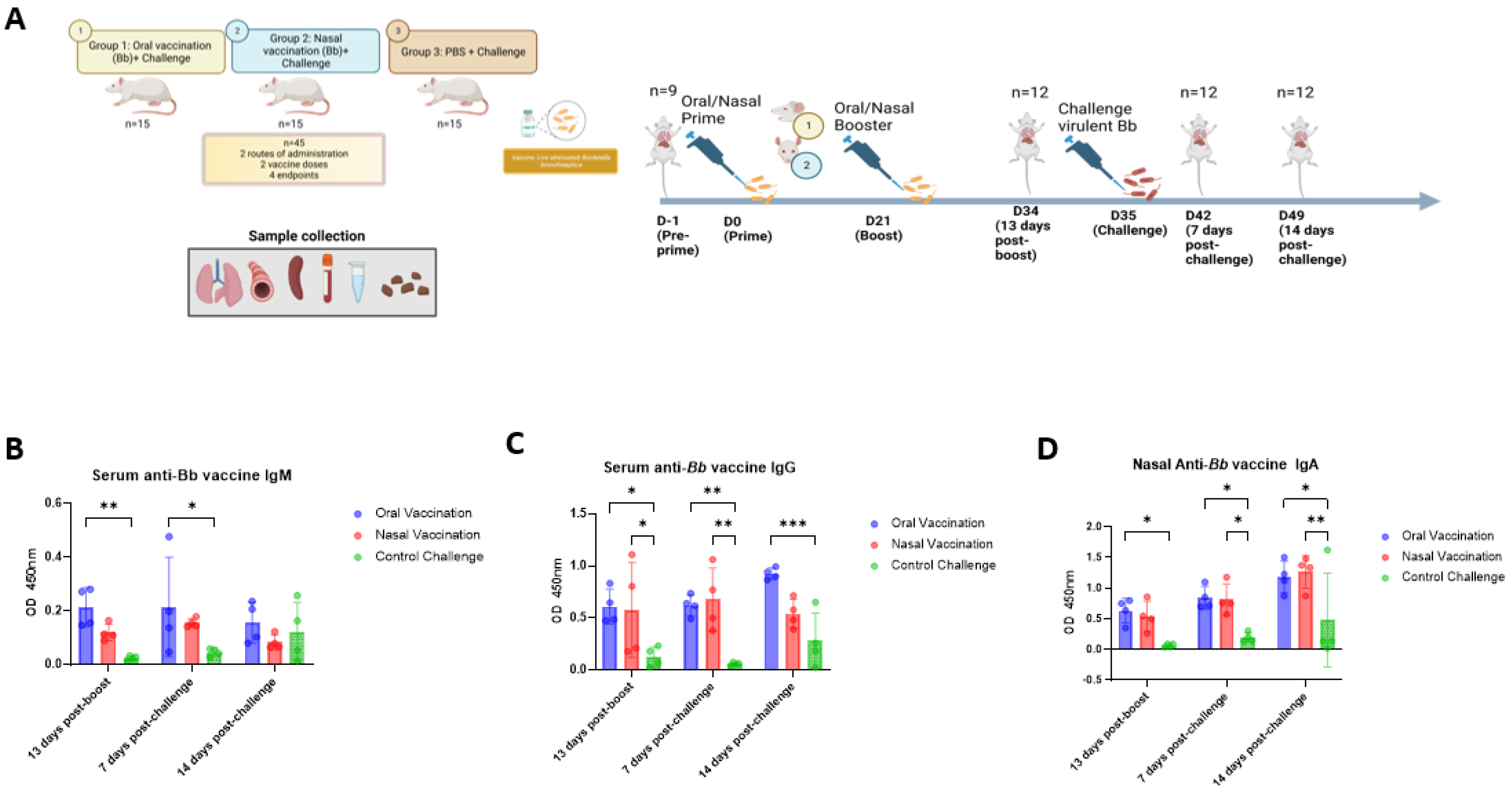
It was observed that vaccination via the oral and intranasal routes induced mucosal and systemic humoral responses while preventing bacterial colonization by a challenge strain and a lower number of lesions in the respiratory tract when compared with the non-vaccinated controls. However, intranasally vaccinated animals coughed significantly less frequently after challenge and had an altered ratio of inflammatory to regulatory cells in the spleen. Importantly, these findings are consistent with the immune response observed in dogs. Both oral and intranasal administrations of a
B. bronchiseptica vaccine in dogs elicit Bb-specific antibodies in the serum [
12,
14,
17,
18,
32]. A study comparing oral with intranasal vaccination in dogs reported no significant difference in median micro-agglutination titers (MAT) in the serum [
14], which is consistent with our observations (
Figure 1B,C). Antibody titers in the unvaccinated animals were heterogeneous, and lower than those in the vaccinated animals after challenge. This may reflect an immune evasion by and the persistence of
B. bronchiseptica, leading to a defective immune response.
B. bronchiseptica lacking the type III secretion system (T3SS) has been shown to elicit higher levels of Bp-specific antibodies [
33,
34]. It is likely that vaccination with a live attenuated strain will impede the modulation of the humoral response by virulent bacteria, while facilitating the recognition of all the immunostimulatory antigens for efficient antibody production [
35].
B. bronchiseptica is a primary respiratory pathogen; thus, mucosal immune responses are likely crucial for protection. This has already been reported for
B. pertussis [
28]. Pathogen-specific IgA is the most important antibody response in mucosal tissues [
27], and their detection serves as an indicator of protective mucosal immunity. In this study, it was observed that oral and intranasal vaccine administrations elicited comparable specific IgA responses in the nasal cavity (
Figure 1D). However, the binding strength of nasal IgA and IgA levels in serum were higher in the intranasal group (
Supplementary Materials, Figures S2 and S3). In dogs, intranasal administration results in a significant increase in IgA levels in the nose and faster induction of antibody production into the serum than subcutaneous administration does [
15]. Another study comparing the oral, intranasal, and injectable routes of vaccination reported IgA titers in nasal swabs that were low before challenge, but that were higher in the orally vaccinated group than in the nasal and control groups [
16]. Different observations were reported for an assessment of the same routes of administration of a vaccine against
B. pertussis in rats [
22]. Hall et al. investigated IgA levels in the lung and nose after challenge and found that only intranasal administration elicited IgA in the nose [
22]. This discrepancy may reflect the disadvantages of the administration of the vaccine via oral gavage compared to the method of micropipette-guided drug administration (MDA), used here. MDA involves delivering the vaccine directly into the mouth with a micropipette, after training the animals to ensure correct intake [
26]. This type of administration may lead to specific immune responses being elicited in the mouth or adjacent mucosae before arriving in the stomach, whereas gavage involves delivery directly to the stomach. MDA may constitute an alternative to gavage, more closely resembling administration in dogs and eliciting mucosal immune responses in the nose. MDA is also a simpler method of administration in rodents. Moreover, a review comparing administration in humans suggested that oral/gastrointestinal administration induced a lower immune response in the respiratory tract than administration though the sublingual route, whereas the efficacy of buccal administration remains debatable [
36].
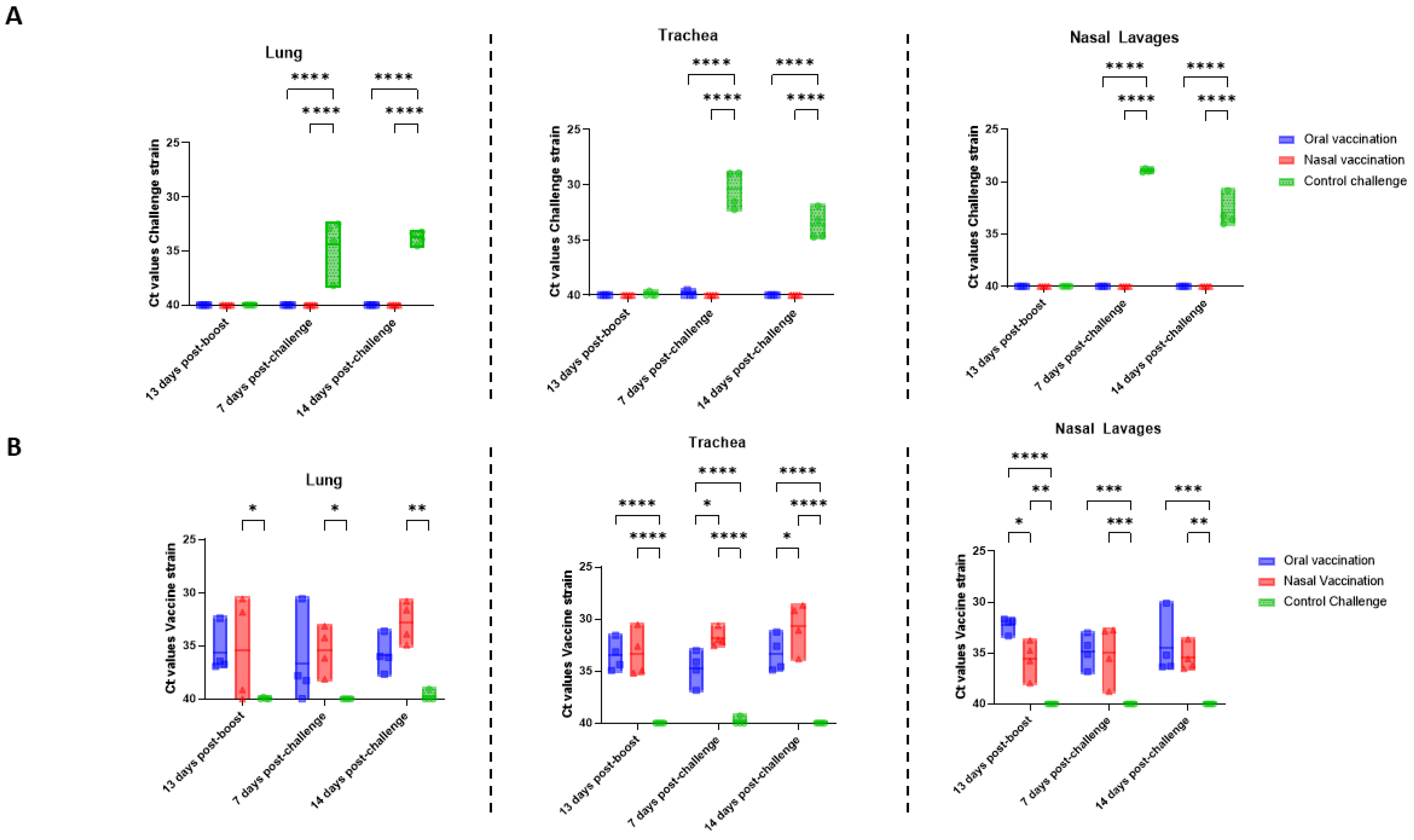
One important aspect of vaccine efficacy in dogs is the decrease in bacterial load and the shedding of
B. bronchiseptica. Both oral and intranasal vaccinations in dogs reduced bacterial load in either the trachea or nasal swabs for up to one year after vaccination [
17,
18]. Consistently with these findings, in our study, the challenge strain was not detected in the nose, trachea, or lungs for up to two weeks after challenge in vaccinated rats (
Figure 2A). Moreover, other
Bordetella vaccine studies in rodents also measured respiratory tract (lung, trachea, and nasal lavages) colonization after challenge and reported similar results [
20]. In contrast, certain published literature reports a lack of bacterial load reduction in dogs [
12,
14]. However, these studies only performed undifferentiated bacterial culture for detection, and it was done shortly after vaccination. This difference may therefore be due to a failure of these studies to distinguish between the challenge and vaccine strains. Indeed, in a study in which nasal swabs were collected after vaccination and before challenge, positive results for bacterial isolation were obtained for animals receiving vaccines via the mucosal route [
16]. One of the major strengths of our study was the availability of primers specifically targeting the challenge and vaccine strains, allowing the differentiation between these two strains via qPCR. The bacterial load of the challenge strain attributed to each animal cannot be confused with the persistence of the vaccine strain in the respiratory tract. The use of these specific primers enabled the revelation that vaccine strain persistence was similar in the nose, trachea, and lungs 28 days after vaccination by both the oral and intranasal route. It has been suggested that the propagation of the vaccine strain in the respiratory tract can stimulate the induction of mucosal IgA [
32], which is consistent with our findings. Indeed, there was an insignificant statistical trend towards a positive correlation between vaccine strain detection in the respiratory tract and the induction of IgA in the nose. Moreover, it was observed that antibody levels in the serum and nose were negatively correlated with trachea colonization of the challenge strain following challenge.
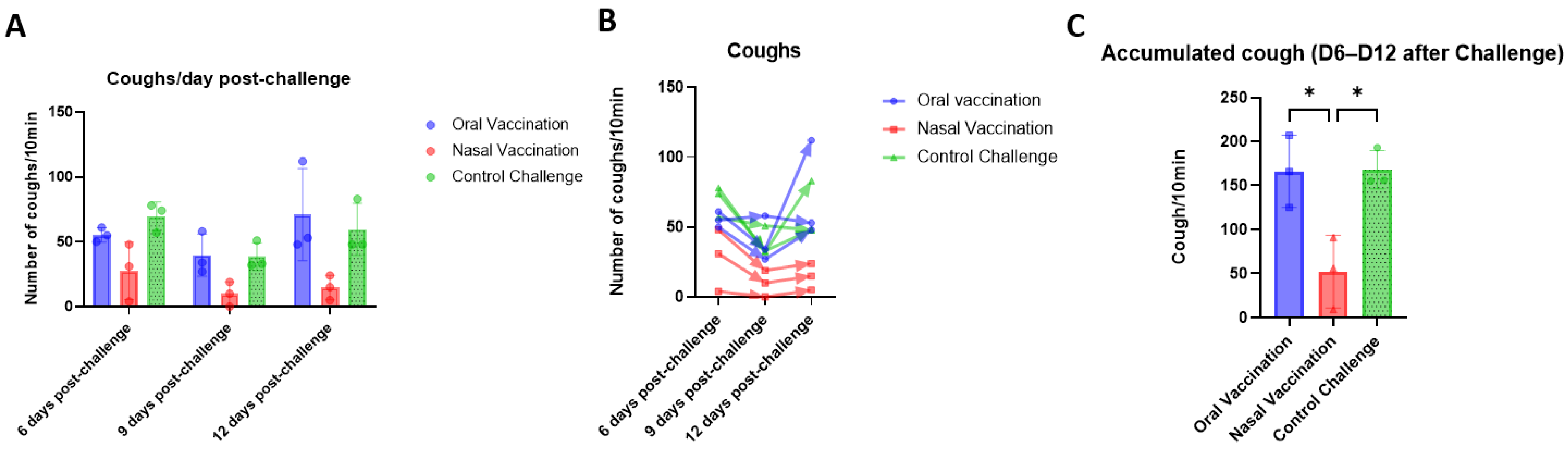
The main clinical sign of CIRDC in dogs is coughing, so an ability to decrease coughing is one of the requirements of any canine vaccine against
B. bronchiseptica. Both the oral and intranasal routes of vaccination decrease disease symptoms in dogs [
12]. Cough has been studied in rats since 1939 [
24] and more recent studies have focused on cough as a lead symptom for studying the pathogenicity of either
B. bronchiseptica [
21] or
B. pertussis [
22,
23]. Nakamura et al. studied the
B. bronchiseptica factors underlying the production of a cough in rats [
21]. Rats were infected with
B. bronchiseptica and the induced cough was evaluated, together with bacterial load. The number of coughs were counted and the bacterial load in the unvaccinated animals in the study described here were consistent with our findings.
On the other hand, vaccination against
B. pertussis via both the oral and intranasal routes decreases the number of coughs after challenge [
22]. Surprisingly, as described here, coughing was reduced significantly only in intranasally vaccinated rats. However, in studies on dogs, cases were defined based on a spontaneous cough for two or more consecutive days [
12]. One limitation of this study was that coughing was measured on separate days, from day 6 post-challenge. Measurements were not taken on consecutive days, so it is possible that some animals did not cough on consecutive days and therefore were not considered to have the disease. Another key consideration is that the mechanisms underlying the cough may differ between dogs and rats [
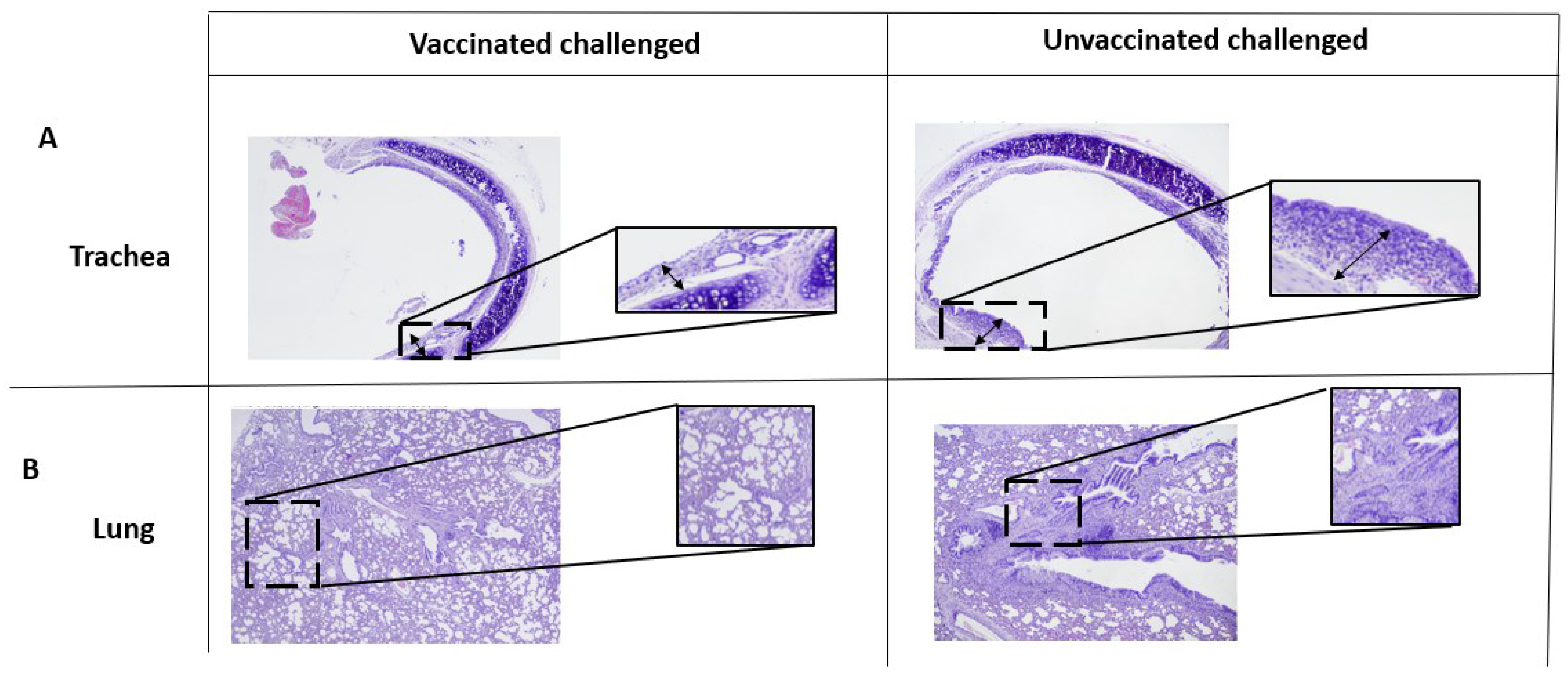
37], and this aspect warrants further exploration. In short, we observed similar immune responses, reductions in bacterial load, similar vaccine replication trends in the respiratory tract via oral or intranasal vaccination, with a varying frequency of coughing. One possible explanation for this could be the presence of the challenge strain in the respiratory tract of the orally vaccinated rats before the first day of sampling after challenge (7 days p.c.). It can be hypothesized that this might have led to the induction of lesions in the respiratory tract such that the cough persisted even though the bacteria were no longer present. However, analyses of the lesions in the respiratory tract demonstrated only that the two routes of vaccination led to a decrease in the severity of respiratory tract lesions two weeks after challenge when compared with that in the nonvaccinated control.
Protective immune responses against
Bordetella species are characterized by antibodies and Th17 cellular responses [
20,
38]. However, to our knowledge, no study of the cellular responses induced via vaccination against
B. bronchiseptica in dogs has ever been published. To gather more insights into the cellular immune responses, immune responses in the spleen, a major organized secondary lymphoid organ, was analyzed. We observed that the ratio of CD3+CD4+ (T helper cells) to CD3+CD8+ (cytotoxic T-cells) cells was similar in the three groups, but that the number of CD3+CD4-CD8-, double-negative (DN) T-cells was surprisingly higher in orally vaccinated animals than in the other groups, suggesting that vaccination could have an influence on their abundance. While there are no known studies directly comparing the percentage of double-negative (DN) T-cells in rat spleens after vaccination, the percentage of CD4 and CD8 out of the total splenocytes has been previously described after
B. pertussis vaccination in mice [
38] and in aging rats [
39], and our results are comparable. Additionally, while not significant, the percentage of these populations seems to be reduced by vaccination or challenge, as seen after
B. pertussis vaccination and challenge in mice [
38].
Double-negative T-cells have been identified as important cells for both tolerance and pathology [
39]. Among them, we can find natural-killer (NK)T-cells, inflammatory or regulatory DNT, DN gamma-delta T-cells (γδ T-cells) and mucosal-associated T (MAIT)-cells [
40]. A role of MAIT-cells could be implicated, based on their known involvement in the defense of the lung and the oral mucosa, and in infectious disease [
40]. However, additional markers will be required to differentiate between these populations and determine if they play a pathological or tolerogenic role in this case. Therefore, it is not yet possible to draw firm conclusions. In addition, the levels of inflammatory T-cells, CD3+CD4+IL17+ (Th17) and regulatory CD3+CD4+FOXP3+CD25+ (Treg) T-cells were assessed. IL-17-producing T helper (Th17) cells are a subset of effector T-cells characterized by the induction of inflammatory cytokines essential for some vaccine responses but also associated with autoimmune diseases [
41]. For instance, respiratory syncytial virus (RSV) infection is more severe if the Th17 response is excessive, but regulatory T-cells can help to counteract the inflammation, preventing disease exacerbation [
42]. Additionally, an imbalance of Th17:Treg cells has been observed to be associated with the severe inflammation seen in COVID-19 [
43], with acute coronary syndromes (ACS) [
44], or the pathogenesis of
Porphyromonas gingivalis [
45]. Hence, an imbalance in the proportions of inflammatory (Th17) and regulatory T-cells (Treg) might account for the clinical signs observed in this study. During data analysis, it was observed that the Th17:Treg ratio 7 days p.c. was significantly higher in orally vaccinated rats when compared with that in intranasally vaccinated rats. However, these data should be interpreted with caution because other mechanisms and cell populations may be at play in modulating these immune responses. Moreover, investigations of the local immune responses in the lung are required to confirm or refute this hypothesis. Differences in cellular responses to oral or nasal administration were previously observed in mice [
46]. Sudo et al. showed that the oral and nasal administration of bacterium-like particles (BLPs) conjugated to an antigen led to a differential secretion of cytokines in lymphoid tissues, such as the spleen, nasal-associated lymphoid tissue (NALT) or Peyer’s patches (PPs) [
46]. This evidence highlights the relevance of the route of administration based on each target, animal species or desired immune response.
The rat model is a valuable tool for the study of clinical symptoms, bacterial load, and humoral and cellular immune responses to
B. bronchiseptica. In addition, the reagents available for immunological studies in rats are superior to those available for canine studies, and rats are more easily accessible than dogs are for research studies. Rats and dogs also have several important anatomic and immunological features in common. A study comparing oral mucosae between species, performed by Thirion-Delalande et al., showed similarities in the immune cell populations present in the oral cavities of rats and dogs [
47]. Moreover, the naval cavity of the rat is similar anatomically to that of the dog, both these cavities being much less similar to the human nasal cavity [
48]. However, there are also major differences in immunological structures between these two species. For example, the rat spleen is considered to be involved in immune defenses, whereas the canine spleen specializes in blood storage and filtration and has underdeveloped immune structures [
49]. There are also differences in the organization and structure of gut-associated lymphoid tissue (GALT) and the nose-associated lymphoid tissue (NALT) between these two species [
49]. For example, tonsils, which play a key role in host defense and antibody responses to oral and nasal pathogens [
50], are present in dogs but not in rats [
51]. In our study, the vaccine was administered via the nose and the mouth. It is therefore possible that the differences in NALT organization between dogs and rats can account for the differences in clinical signs between these two species.
Importantly, more studies are required to characterize the cellular immune response against
B. bronchiseptica more fully. Furthermore, the organs involved in these responses are not only in the spleen but also in the local tissues of the respiratory tract and thus should be further investigated. Both the intranasal and oral vaccination routes have been shown to elicit protection for more than a year in dogs [
17,
18]. An understanding of the mechanisms involved will be required for the development of the next generation of vaccines. Finally, assessments of antibody functionality will also be required to evaluate the correlation between the presence of antibodies and disease correctly.
Notwithstanding these limitations, our findings suggest that mucosal vaccination against
B. bronchiseptica provides effective protection against colonization and shedding of the challenge strain in rats. The reasons for the differences in clinical signs observed remain unclear, but oral vaccine administration remains more convenient than nasal administration, which can be difficult in dogs. This study constitutes a first step towards enhancing the understanding of the immune response to
B. bronchiseptica and the immunological differences between routes of administration. It also highlights the rat model as a surrogate system suitable for use in the future development of mucosal vaccines for dogs.
B. bronchiseptica is the major pathogen responsible for CIRDC, but it is not the only pathogen implicated in this disease [
52]. This model could be used to assess protection against other respiratory pathogens involved, such as canine parainfluenza virus (CPIV). Rats are naturally susceptible to the Sendai virus, which is of the same family as CPIV, the Paramyxoviridae family [
53]. Therefore, in future studies, it would be of interest to demonstrate the susceptibility and induction of the disease of CPIV in rats. Thus, rats could potentially be used as a surrogate model for preliminary in vivo testing before conducting definitive tests in dogs.

 ,
, 






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}