

Haemato-Immunological Response of Immunized Atlantic Salmon (Salmo salar) to Moritella viscosa Challenge and Antigens



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fish Holding
2.2. Infection Trials
2.2.1. Inoculum Preparation
2.2.2. Challenge Assay in Vaccinated Atlantic Salmon
2.3. Flow Cytometry
2.4. Immune Confocal Microscopy
2.5. IgM Titer Determination Using Indirect Enzyme-Linked Immunosorbent Assay (ELISA)
2.6. M. visosa Antigenicity in Farmed Atlantic Salmon
2.6.1. Vaccine Preparations
2.6.2. Antigenicity of M. viscosa Components in Farmed Atlantic Salmon
2.6.3. M. viscosa Challenge in Atlantic Salmon
2.7. Statistical Analysis
3. Results
3.1. Susceptibility of Vaccinated Farmed Atlantic Salmon to M. viscosa Challenge
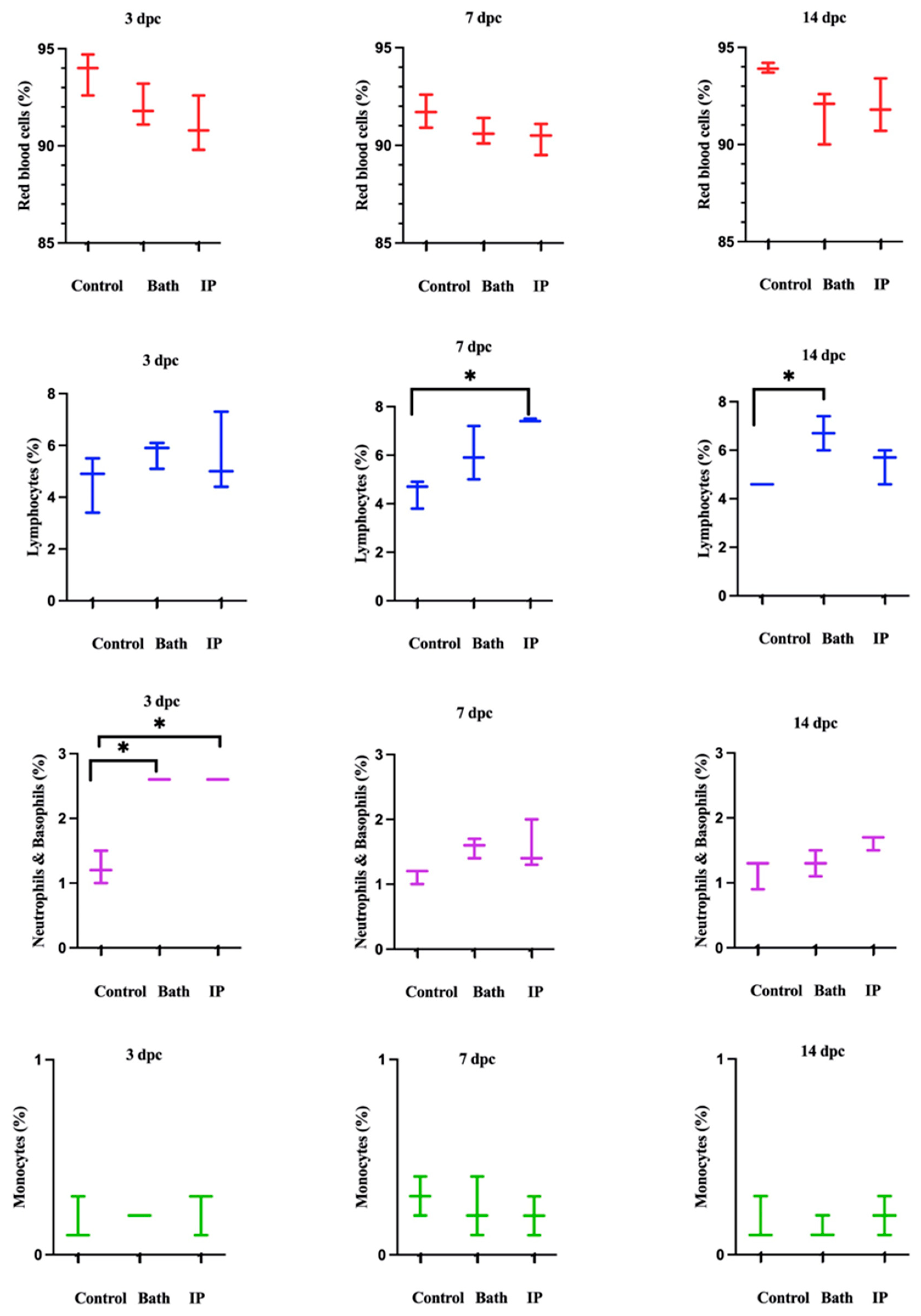
3.2. Cell Populations in the Challenged Atlantic Salmon
3.3. Confocal Microscopy Analysis
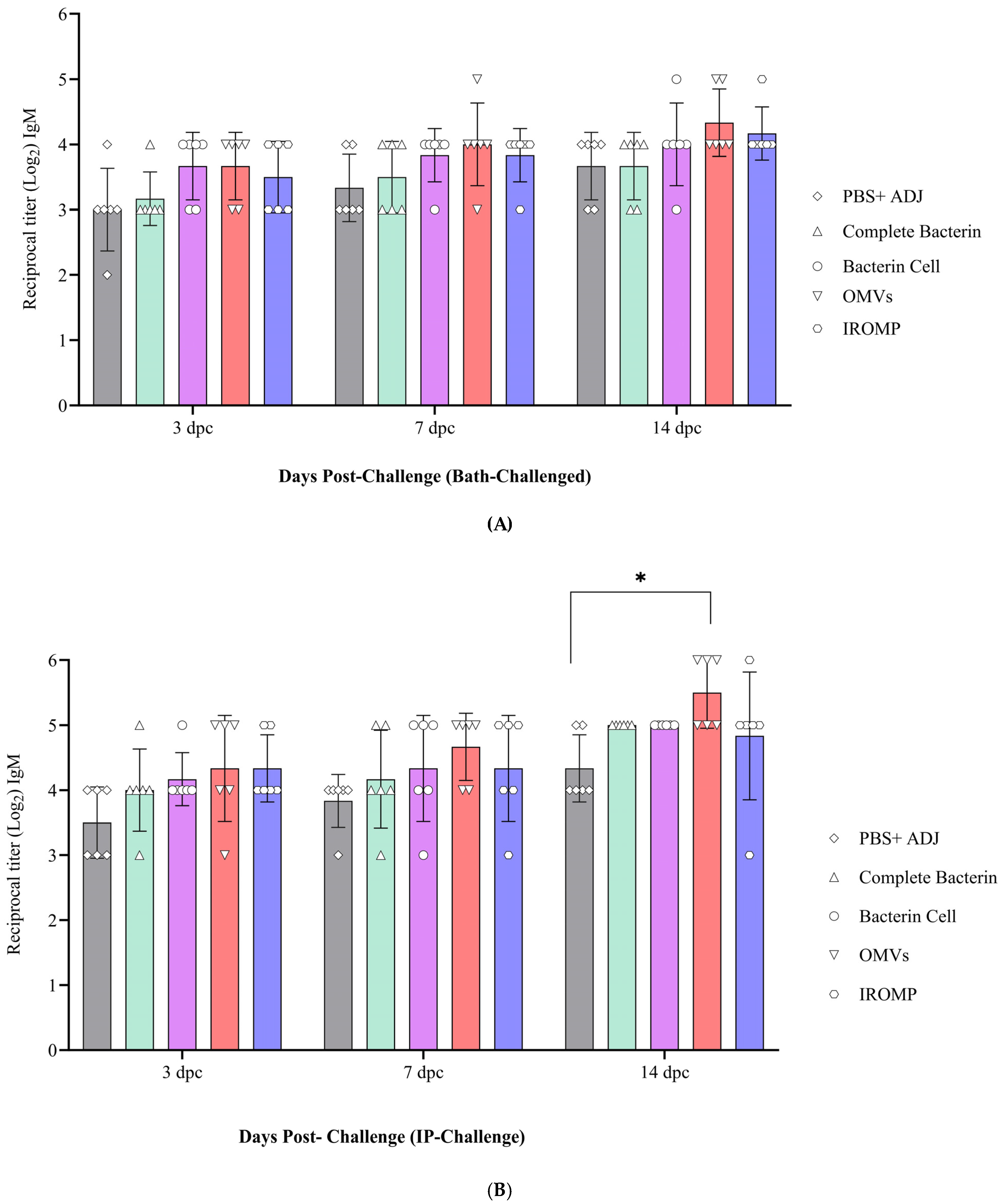
3.4. IgM Titer Levels in the Challenged Atlantic Salmon
3.5. Detection of Antibody after Booster Dose by Prepared Vaccines
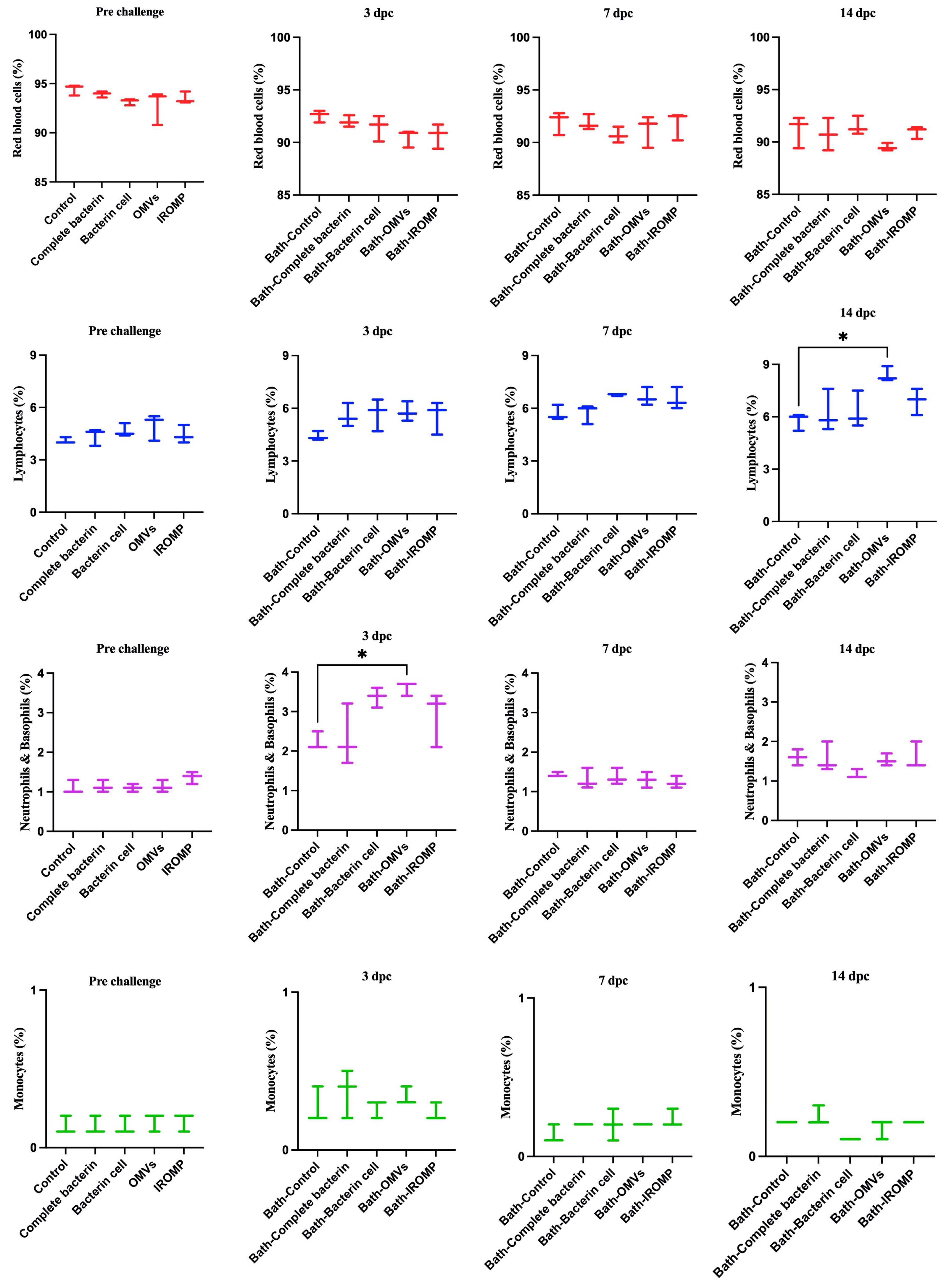
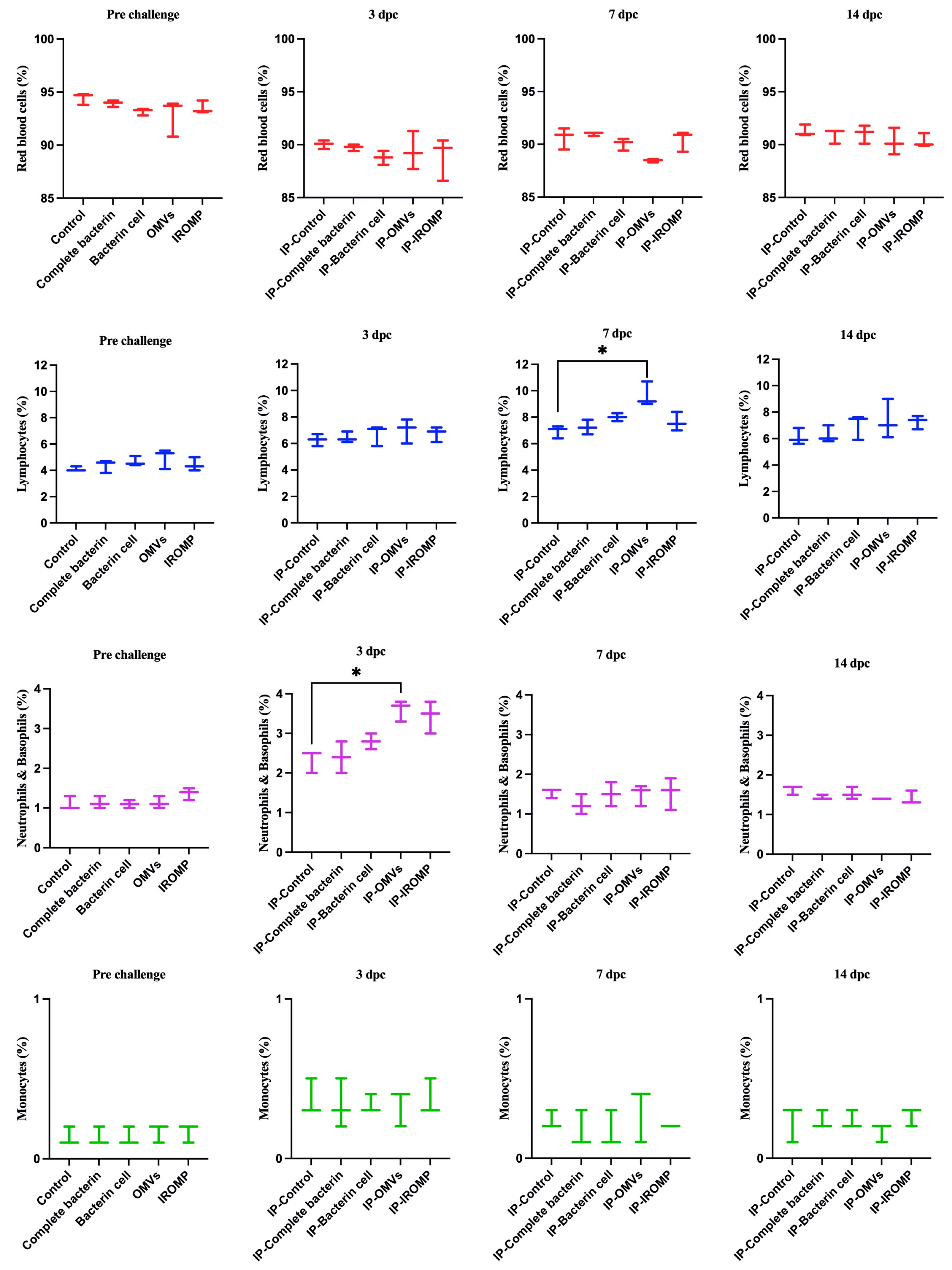
3.6. Cell Population after Challenge in the Boosted Groups
3.7. Detection of Antibody Levels after Challenge in the Boosted Groups
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Løvoll, M.; Wiik-Nielsen, C.; Tunsjø, H.S.; Colquhoun, D.; Lunder, T.; Sørum, H.; Grove, S. Atlantic salmon bath challenged with Moritella viscosa–pathogen invasion and host response. Fish Shellfish Immunol. 2009, 26, 877–884. [Google Scholar] [CrossRef] [PubMed]
- Grove, S.; Wiik-Nielsen, C.; Lunder, T.; Tunsjø, H.; Tandstad, N.; Reitan, L.; Marthinussen, A.; Sørgaard, M.; Olsen, A.; Colquhoun, D. Previously unrecognised division within Moritella viscosa isolated from fish farmed in the North Atlantic. Dis. Aquat. Org. 2010, 93, 51–61. [Google Scholar] [CrossRef] [PubMed]
- Karlsen, C.; Thorarinsson, R.; Wallace, C.; Salonius, K.; Midtlyng, P.J. Atlantic salmon winter-ulcer disease: Combining mortality and skin ulcer development as clinical efficacy criteria against Moritella viscosa infection. Aquaculture 2017, 473, 538–544. [Google Scholar] [CrossRef]
- MacKinnon, B.; Groman, D.; Fast, M.D.; Manning, A.J.; Jones, P.; St-Hilaire, S. Atlantic salmon challenged with extracellular products from Moritella viscosa. Dis. Aquat. Org. 2019, 133, 119–125. [Google Scholar] [CrossRef]
- MacKinnon, B.; Jones, P.; Hawkins, L.; Dohoo, I.; Stryhn, H.; Vanderstichel, R.; St-Hilaire, S. The epidemiology of skin ulcers in saltwater reared Atlantic salmon (Salmo salar) in Atlantic Canada. Aquaculture 2019, 501, 230–238. [Google Scholar] [CrossRef]
- Mondal, H.; Thomas, J. A review on the recent advances and application of vaccines against fish pathogens in aquaculture. Aquac. Int. 2022, 30, 1971–2000. [Google Scholar] [CrossRef]
- Shoemaker, C.A.; Klesius, P.H.; Evans, J.J.; Arias, C.R. Use of modified live vaccines in aquaculture. J. World Aquac. Soc. 2009, 40, 573–585. [Google Scholar] [CrossRef]
- Ghasemieshkaftaki, M.; Vasquez, I.; Eshraghi, A.; Gamperl, A.K.; Santander, J. Comparative Genomic Analysis of a Novel Vibrio sp. Isolated from an Ulcer Disease Event in Atlantic Salmon (Salmo salar). Microorganisms 2023, 11, 1736. [Google Scholar] [CrossRef]
- Secombes, C. Will advances in fish immunology change vaccination strategies? Fish Shellfish Immunol. 2008, 25, 409–416. [Google Scholar] [CrossRef]
- Tafalla, C.; Bøgwald, J.; Dalmo, R.A. Adjuvants and immunostimulants in fish vaccines: Current knowledge and future perspectives. Fish Shellfish Immunol. 2013, 35, 1740–1750. [Google Scholar] [CrossRef]
- Yamaguchi, T.; Quillet, E.; Boudinot, P.; Fischer, U. What could be the mechanisms of immunological memory in fish? Fish Shellfish Immunol. 2019, 85, 3–8. [Google Scholar] [CrossRef] [PubMed]
- Kayansamruaj, P.; Areechon, N.; Unajak, S. Development of fish vaccine in Southeast Asia: A challenge for the sustainability of SE Asia aquaculture. Fish Shellfish Immunol. 2020, 103, 73–87. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Bruce, T.J.; Jones, E.M.; Cain, K.D. A review of fish vaccine development strategies: Conventional methods and modern biotechnological approaches. Microorganisms 2019, 7, 569. [Google Scholar] [CrossRef] [PubMed]
- Evensen, Ø. Development of fish vaccines: Focusing on methods. In Fish Vaccines; Springer: Basel, Switzerland, 2016; pp. 53–74. [Google Scholar]
- Durbin, M.; McIntosh, D.; Smith, P.; Wardle, R.; Austin, B. Immunization against furunculosis in rainbow trout with iron-regulated outer membrane protein vaccines: Relative efficacy of immersion, oral, and injection delivery. J. Aquat. Anim. Health 1999, 11, 68–75. [Google Scholar] [CrossRef]
- Björnsson, H.; Marteinsson, V.; Friðjónsson, Ó.; Linke, D.; Benediktsdottir, E. Isolation and characterization of an antigen from the fish pathogen Moritella viscosa. J. Appl. Microbiol. 2011, 111, 17–25. [Google Scholar] [CrossRef] [PubMed]
- Sartorio, M.G.; Pardue, E.J.; Feldman, M.F.; Haurat, M.F. Bacterial outer membrane vesicles: From discovery to applications. Annu. Rev. Microbiol. 2021, 75, 609–630. [Google Scholar] [CrossRef] [PubMed]
- Prior, J.T.; Davitt, C.; Kurtz, J.; Gellings, P.; McLachlan, J.B.; Morici, L.A. Bacterial-derived outer membrane vesicles are potent adjuvants that drive humoral and cellular immune responses. Pharmaceutics 2021, 13, 131. [Google Scholar] [CrossRef]
- Anand, D.; Chaudhuri, A. Bacterial outer membrane vesicles: New insights and applications. Mol. Membr. Biol. 2016, 33, 125–137. [Google Scholar] [CrossRef]
- Balhuizen, M.D.; Veldhuizen, E.J.; Haagsman, H.P. Outer membrane vesicle induction and isolation for vaccine development. Front. Microbiol. 2021, 12, 629090. [Google Scholar] [CrossRef]
- Mehanny, M.; Lehr, C.-M.; Fuhrmann, G. Extracellular vesicles as antigen carriers for novel vaccination avenues. Adv. Drug Deliv. Rev. 2021, 173, 164–180. [Google Scholar] [CrossRef]
- Gomez, D.; Sunyer, J.O.; Salinas, I. The mucosal immune system of fish: The evolution of tolerating commensals while fighting pathogens. Fish Shellfish Immunol. 2013, 35, 1729–1739. [Google Scholar] [CrossRef] [PubMed]
- Koshio, S. Immunotherapies targeting fish mucosal immunity–Current knowledge and future perspectives. Front. Immunol. 2016, 6, 643. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Wang, Q.; Huang, Z.; Ding, L.; Xu, Z. Immunoglobulins, mucosal immunity and vaccination in teleost fish. Front. Immunol. 2020, 11, 567941. [Google Scholar] [CrossRef] [PubMed]
- Mokhtar, D.M.; Zaccone, G.; Alesci, A.; Kuciel, M.; Hussein, M.T.; Sayed, R.K. Main components of fish immunity: An overview of the fish immune system. Fishes 2023, 8, 93. [Google Scholar] [CrossRef]
- Smith, N.C.; Rise, M.L.; Christian, S.L. A comparison of the innate and adaptive immune systems in cartilaginous fish, ray-finned fish, and lobe-finned fish. Front. Immunol. 2019, 10, 2292. [Google Scholar] [CrossRef]
- Bengtén, E.; Clem, L.W.; Miller, N.W.; Warr, G.W.; Wilson, M. Channel catfish immunoglobulins: Repertoire and expression. Dev. Comp. Immunol. 2006, 30, 77–92. [Google Scholar] [CrossRef]
- Solem, S.T.; Stenvik, J. Antibody repertoire development in teleosts—A review with emphasis on salmonids and Gadus morhua L. Dev. Comp. Immunol. 2006, 30, 57–76. [Google Scholar] [CrossRef]
- Mu, Q.; Dong, Z.; Kong, W.; Wang, X.; Yu, J.; Ji, W.; Su, J.; Xu, Z. Response of immunoglobulin M in gut mucosal immunity of common carp (Cyprinus carpio) infected with Aeromonas hydrophila. Front. Immunol. 2022, 13, 1037517. [Google Scholar] [CrossRef]
- Isla, A.; Sánchez, P.; Ruiz, P.; Albornoz, R.; Pontigo, J.P.; Rauch, M.C.; Hawes, C.; Vargas-Chacoff, L.; Yáñez, A.J. Effect of low-dose Piscirickettsia salmonis infection on haematological-biochemical blood parameters in Atlantic salmon (Salmo salar). J. Fish Biol. 2022, 101, 1021–1032. [Google Scholar] [CrossRef]
- Monir, M.S.; Yusoff, S.b.M.; Zulperi, Z.b.M.; Hassim, H.b.A.; Mohamad, A.; Ngoo, M.S.b.M.H.; Ina-Salwany, M.Y. Haemato-immunological responses and effectiveness of feed-based bivalent vaccine against Streptococcus iniae and Aeromonas hydrophila infections in hybrid red tilapia (Oreochromis mossambicus × O. niloticus). BMC Vet. Res. 2020, 16, 226. [Google Scholar] [CrossRef]
- Fazio, F. Fish hematology analysis as an important tool of aquaculture: A review. Aquaculture 2019, 500, 237–242. [Google Scholar] [CrossRef]
- Gordeev, I.I.; Mikryakov, D.V.; Balabanova, L.V.; Mikryakov, V.R. Composition of leucocytes in peripheral blood of Patagonian toothfish (Dissostichus eleginoides, Smitt, 1898)(Nototheniidae). Polar Res. 2017, 36, 1374126. [Google Scholar] [CrossRef]
- Witeska, M.; Kondera, E.; Ługowska, K.; Bojarski, B. Hematological methods in fish–Not only for beginners. Aquaculture 2022, 547, 737498. [Google Scholar] [CrossRef]
- Soto-Dávila, M.; Hossain, A.; Chakraborty, S.; Rise, M.L.; Santander, J. Aeromonas salmonicida subsp. salmonicida early infection and immune response of Atlantic cod (Gadus morhua L.) primary macrophages. Front. Immunol. 2019, 10, 1237. [Google Scholar] [CrossRef] [PubMed]
- Connors, E.; Soto-Dávila, M.; Hossain, A.; Vasquez, I.; Gnanagobal, H.; Santander, J. Identification and validation of reliable Aeromonas salmonicida subspecies salmonicida reference genes for differential gene expression analyses. Infect. Genet. Evol. 2019, 73, 314–321. [Google Scholar] [CrossRef]
- Leboffe, M.J.; Pierce, B.E. Microbiology: Laboratory Theory and Application; Morton Publishing Company: Englewood, CO, USA, 2015. [Google Scholar]
- Gross, A.; Schoendube, J.; Zimmermann, S.; Steeb, M.; Zengerle, R.; Koltay, P. Technologies for single-cell isolation. Int. J. Mol. Sci. 2015, 16, 16897–16919. [Google Scholar] [CrossRef]
- Inoue, T.; Moritomo, T.; Tamura, Y.; Mamiya, S.; Fujino, H.; Nakanishi, T. A new method for fish leucocyte counting and partial differentiation by flow cytometry. Fish Shellfish Immunol. 2002, 13, 379–390. [Google Scholar] [CrossRef]
- Erkinharju, T.; Lundberg, M.R.; Isdal, E.; Hordvik, I.; Dalmo, R.A.; Seternes, T. Studies on the antibody response and side effects after intramuscular and intraperitoneal injection of Atlantic lumpfish (Cyclopterus lumpus L.) with different oil-based vaccines. J. Fish Dis. 2017, 40, 1805–1813. [Google Scholar]
- Rønneseth, A.; Haugland, G.T.; Colquhoun, D.J.; Brudal, E.; Wergeland, H.I. Protection and antibody reactivity following vaccination of lumpfish (Cyclopterus lumpus L.) against atypical Aeromonas salmonicida. Fish Shellfish Immunol. 2017, 64, 383–391. [Google Scholar] [CrossRef]
- Crowther, J.R. The ELISA Guidebook; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2008; Volume 149. [Google Scholar]
- Dang, M.; Cao, T.; Vasquez, I.; Hossain, A.; Gnanagobal, H.; Kumar, S.; Hall, J.R.; Monk, J.; Boyce, D.; Westcott, J. Oral immunization of larvae and juvenile of lumpfish (Cyclopterus lumpus) against Vibrio anguillarum does not influence systemic immunity. Vaccines 2021, 9, 819. [Google Scholar] [CrossRef]
- Vasquez, I.; Cao, T.; Hossain, A.; Valderrama, K.; Gnanagobal, H.; Dang, M.; Leeuwis, R.H.; Ness, M.; Campbell, B.; Gendron, R. Aeromonas salmonicida infection kinetics and protective immune response to vaccination in sablefish (Anoplopoma fimbria). Fish Shellfish Immunol. 2020, 104, 557–566. [Google Scholar] [CrossRef] [PubMed]
- Eslamloo, K.; Kumar, S.; Caballero-Solares, A.; Gnanagobal, H.; Santander, J.; Rise, M.L. Profiling the transcriptome response of Atlantic salmon head kidney to formalin-killed Renibacterium salmoninarum. Fish Shellfish Immunol. 2020, 98, 937–949. [Google Scholar] [CrossRef] [PubMed]
- Hirst, I.; Ellis, A.E. Iron-regulated outer membrane proteins of Aeromonas salmonicida are important protective antigens in Atlantic salmon against furunculosis. Fish Shellfish Immunol. 1994, 4, 29–45. [Google Scholar] [CrossRef]
- Santander, J.; Golden, G.; Wanda, S.-Y.; Curtiss III, R. Fur-regulated iron uptake system of Edwardsiella ictaluri and its influence on pathogenesis and immunogenicity in the catfish host. Infect. Immun. 2012, 80, 2689–2703. [Google Scholar] [CrossRef] [PubMed]
- Sambrook, J.; Russell, D. Molecular Cloning: A Laboratory Manual, 3rd ed.; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 2001; Volume 413. [Google Scholar]
- Brudal, E.; Lampe, E.O.; Reubsaet, L.; Roos, N.; Hegna, I.K.; Thrane, I.M.; Koppang, E.O.; Winther-Larsen, H.C. Vaccination with outer membrane vesicles from Francisella noatunensis reduces development of francisellosis in a zebrafish model. Fish Shellfish Immunol. 2015, 42, 50–57. [Google Scholar] [CrossRef] [PubMed]
- Karlsen, C.; Vanberg, C.; Mikkelsen, H.; Sørum, H. Co-infection of Atlantic salmon (Salmo salar), by Moritella viscosa and Aliivibrio wodanis, development of disease and host colonization. Vet. Microbiol. 2014, 171, 112–121. [Google Scholar] [CrossRef] [PubMed]
- Poppe, T.T.; Bæverfjord, G.; Hansen, T. Effects of intensive production with emphasis on on-growing production: Fast growth, deformities and production-related diseases. In Aquaculture Research: From Cage to Consumption; Norges Forskningsråd: Oslo, Norway, 2007; pp. 120–135. [Google Scholar]
- Adams, A. Progress, challenges and opportunities in fish vaccine development. Fish Shellfish Immunol. 2019, 90, 210–214. [Google Scholar] [CrossRef]
- Rozas-Serri, M.; Correa, R.; Walker-Vergara, R.; Coñuecar, D.; Barrientos, S.; Leiva, C.; Ildefonso, R.; Senn, C.; Peña, A. Reference Intervals for Blood Biomarkers in Farmed Atlantic Salmon, Coho Salmon and Rainbow Trout in Chile: Promoting a Preventive Approach in Aquamedicine. Biology 2022, 11, 1066. [Google Scholar] [CrossRef]
- Ley, K.; Laudanna, C.; Cybulsky, M.I.; Nourshargh, S. Getting to the site of inflammation: The leukocyte adhesion cascade updated. Nat. Rev. Immunol. 2007, 7, 678–689. [Google Scholar] [CrossRef]
- Chavakis, E.; Choi, E.Y.; Chavakis, T. Novel aspects in the regulation of the leukocyte adhesion cascade. Thromb. Haemost. 2009, 102, 191–197. [Google Scholar] [CrossRef]
- Janeway Jr, C.A. Immunobiology the Immune System in Health and Disease; Garland Science: New York, NY, USA, 1997. [Google Scholar]
- Mutoloki, S.; Jørgensen, J.B.; Evensen, Ø. The adaptive immune response in fish. In Fish Vaccination; Wiley: Hoboken, NJ, USA, 2014; pp. 104–115. [Google Scholar]
- Harikrishnan, R.; Rani, M.N.; Balasundaram, C. Hematological and biochemical parameters in common carp, Cyprinus carpio, following herbal treatment for Aeromonas hydrophila infection. Aquaculture 2003, 221, 41–50. [Google Scholar] [CrossRef]
- Afiyanti, A.D.; Yuliani, M.G.A.; Handijatno, D. Leukocyte count and differential leukocyte count of carp (Cyprinus carpio Linn) after infected by Aeromonas salmonicida. Cell 2018, 2000, 3. [Google Scholar]
- Martins, M.; Mouriño, J.; Amaral, G.; Vieira, F.; Dotta, G.; Jatobá, A.; Pedrotti, F.; Jerônimo, G.; Buglione-Neto, C. Haematological changes in Nile tilapia experimentally infected with Enterococcus sp. Braz. J. Biol. 2008, 68, 657–661. [Google Scholar] [CrossRef]
- Sheng, X.-Z.; Xu, G.-J.; Tang, X.-Q.; Zhan, W.-B. Monoclonal antibodies recognizing mucus immunoglobulin and surface immunoglobulin-positive cells of flounder (Paralichthys olivaceus). Vet. Immunol. Immunopathol. 2012, 145, 143–150. [Google Scholar] [CrossRef] [PubMed]
- Sheng, X.; Qian, X.; Tang, X.; Xing, J.; Zhan, W. Polymeric immunoglobulin receptor mediates immune excretion of mucosal IgM–antigen complexes across intestinal epithelium in flounder (Paralichthys olivaceus). Front. Immunol. 2018, 9, 1562. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Barreda, D.R.; Zhang, Y.-A.; Boshra, H.; Gelman, A.E.; LaPatra, S.; Tort, L.; Sunyer, J.O. B lymphocytes from early vertebrates have potent phagocytic and microbicidal abilities. Nat. Immunol. 2006, 7, 1116–1124. [Google Scholar] [CrossRef]
- Wu, L.; Yang, Y.; Gao, A.; Li, J.; Ye, J. Teleost fish IgM+ plasma-like cells possess IgM-secreting, phagocytic, and antigen-presenting capacities. Front. Immunol. 2022, 13, 1016974. [Google Scholar] [CrossRef]
- van der Wal, Y.A.; Jenberie, S.; Nordli, H.; Greiner-Tollersrud, L.; Kool, J.; Jensen, I.; Jørgensen, J.B. The importance of the Atlantic salmon peritoneal cavity B cell response: Local IgM secreting cells are predominant upon Piscirickettsia salmonis infection. Dev. Comp. Immunol. 2021, 123, 104125. [Google Scholar] [CrossRef]
- Dumetz, F.; Duchaud, E.; Claverol, S.; Orieux, N.; Papillon, S.; Lapaillerie, D.; Le Henaff, M. Analysis of the Flavobacterium psychrophilum outer-membrane subproteome and identification of new antigenic targets for vaccine by immunomics. Microbiology 2008, 154, 1793–1801. [Google Scholar] [CrossRef]
- Jan, A.T. Outer membrane vesicles (OMVs) of gram-negative bacteria: A perspective update. Front. Microbiol. 2017, 8, 1053. [Google Scholar] [CrossRef]
- Ellis, T.N.; Kuehn, M.J. Virulence and immunomodulatory roles of bacterial outer membrane vesicles. Microbiol. Mol. Biol. Rev. 2010, 74, 81–94. [Google Scholar] [CrossRef] [PubMed]
- Koeppen, K.; Hampton, T.H.; Jarek, M.; Scharfe, M.; Gerber, S.A.; Mielcarz, D.W.; Demers, E.G.; Dolben, E.L.; Hammond, J.H.; Hogan, D.A. A novel mechanism of host-pathogen interaction through sRNA in bacterial outer membrane vesicles. PLoS Pathog. 2016, 12, e1005672. [Google Scholar] [CrossRef] [PubMed]
- Furuyama, N.; Sircili, M.P. Outer membrane vesicles (OMVs) produced by gram-negative bacteria: Structure, functions, biogenesis, and vaccine application. BioMed Res. Int. 2021, 2021, 1490732. [Google Scholar] [CrossRef] [PubMed]
- Schwechheimer, C.; Kuehn, M.J. Outer-membrane vesicles from Gram-negative bacteria: Biogenesis and functions. Nat. Rev. Microbiol. 2015, 13, 605–619. [Google Scholar] [CrossRef] [PubMed]
- Tandberg, J.; Oliver, C.; Lagos, L.; Gaarder, M.; Yáñez, A.J.; Ropstad, E.; Winther-Larsen, H.C. Membrane vesicles from Piscirickettsia salmonis induce protective immunity and reduce development of salmonid rickettsial septicemia in an adult zebrafish model. Fish Shellfish Immunol. 2017, 67, 189–198. [Google Scholar] [CrossRef] [PubMed]
- Oliver, C.; Coronado, J.L.; Martínez, D.; Kashulin-Bekkelund, A.; Lagos, L.X.; Ciani, E.; Sanhueza-Oyarzún, C.; Mancilla-Nova, A.; Enríquez, R.; Winther-Larsen, H.C. Outer membrane vesicles from Piscirickettsia salmonis induce the expression of inflammatory genes and production of IgM in Atlantic salmon Salmo salar. Fish Shellfish Immunol. 2023, 139, 108887. [Google Scholar] [CrossRef] [PubMed]
- Hong, G.-E.; Kim, D.-G.; Park, E.-M.; Nam, B.-H.; Kim, Y.-O.; Kong, I.-S. Identification of Vibrio anguillarum outer membrane vesicles related to immunostimulation in the Japanese flounder, Paralichthys olivaceus. Biosci. Biotechnol. Biochem. 2009, 73, 437–439. [Google Scholar] [CrossRef]
- Gerritzen, M.J.; Martens, D.E.; Wijffels, R.H.; van der Pol, L.; Stork, M. Bioengineering bacterial outer membrane vesicles as vaccine platform. Biotechnol. Adv. 2017, 35, 565–574. [Google Scholar] [CrossRef]
- Kashyap, D.; Panda, M.; Baral, B.; Varshney, N.; Bhandari, V.; Parmar, H.S.; Prasad, A.; Jha, H.C. Outer membrane vesicles: An emerging vaccine platform. Vaccines 2022, 10, 1578. [Google Scholar] [CrossRef]
- Romstad, A.B.; Reitan, L.J.; Midtlyng, P.; Gravningen, K.; Evensen, Ø. Development of an antibody ELISA for potency testing of furunculosis (Aeromonas salmonicida subsp salmonicida) vaccines in Atlantic salmon (Salmo salar L.). Biologicals 2012, 40, 67–71. [Google Scholar] [CrossRef]
- Liu, S.; Wang, D.; Cao, Y.; Lu, T.; Liu, H.; Li, S. Effects of propolis on the immune enhancement of the formalin-inactivated Aeromonas salmonicida vaccine. Aquac. Res. 2020, 51, 4759–4770. [Google Scholar] [CrossRef]
- Wang, H.-R.; Hu, Y.-H.; Zhang, W.-W.; Sun, L. Construction of an attenuated Pseudomonas fluorescens strain and evaluation of its potential as a cross-protective vaccine. Vaccine 2009, 27, 4047–4055. [Google Scholar] [CrossRef] [PubMed]
- Raida, M.K.; Nylén, J.; Holten-Andersen, L.; Buchmann, K. Association between plasma antibody response and protection in rainbow trout Oncorhynchus mykiss immersion vaccinated against Yersinia ruckeri. PLoS ONE 2011, 6, e18832. [Google Scholar] [CrossRef] [PubMed]
- Villumsen, K.R.; Dalsgaard, I.; Holten-Andersen, L.; Raida, M.K. Correction: Potential Role of Specific Antibodies as Important Vaccine Induced Protective Mechanism against Aeromonas salmonicida in Rainbow Trout. PLoS ONE 2012, 7, e46733. [Google Scholar] [CrossRef]










Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ghasemieshkaftaki, M.; Cao, T.; Hossain, A.; Vasquez, I.; Santander, J. Haemato-Immunological Response of Immunized Atlantic Salmon (Salmo salar) to Moritella viscosa Challenge and Antigens. Vaccines 2024, 12, 70. https://doi.org/10.3390/vaccines12010070
Ghasemieshkaftaki M, Cao T, Hossain A, Vasquez I, Santander J. Haemato-Immunological Response of Immunized Atlantic Salmon (Salmo salar) to Moritella viscosa Challenge and Antigens. Vaccines. 2024; 12(1):70. https://doi.org/10.3390/vaccines12010070
Chicago/Turabian StyleGhasemieshkaftaki, Maryam, Trung Cao, Ahmed Hossain, Ignacio Vasquez, and Javier Santander. 2024. "Haemato-Immunological Response of Immunized Atlantic Salmon (Salmo salar) to Moritella viscosa Challenge and Antigens" Vaccines 12, no. 1: 70. https://doi.org/10.3390/vaccines12010070
APA StyleGhasemieshkaftaki, M., Cao, T., Hossain, A., Vasquez, I., & Santander, J. (2024). Haemato-Immunological Response of Immunized Atlantic Salmon (Salmo salar) to Moritella viscosa Challenge and Antigens. Vaccines, 12(1), 70. https://doi.org/10.3390/vaccines12010070