

Measuring Variant-Specific Neutralizing Antibody Profiles after Bivalent SARS-CoV-2 Vaccinations Using a Multivariant Surrogate Virus Neutralization Microarray

 , , and
, , and 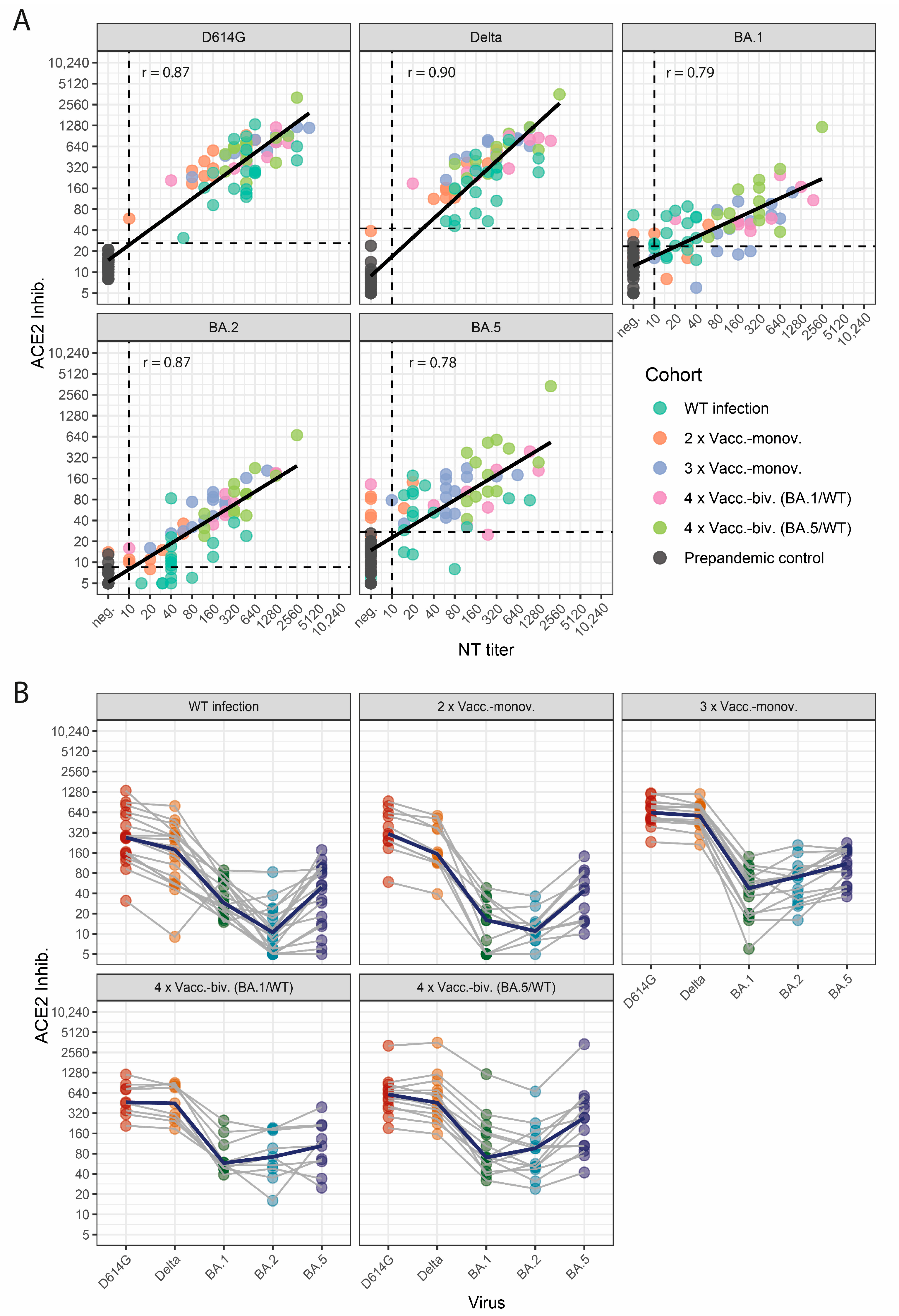{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Cohort
2.2. Live-Virus NTs
2.3. Multivariant Surrogate-Virus Neutralization Test
2.4. Statistical Analysis
3. Results
3.1. Correlation of the Multivariant sVNT with Variant-Specific Live-Virus NTs
3.2. Profiles of Neutralizing Activity after SARS-CoV-2 Wild-Type Infection and Vaccinations
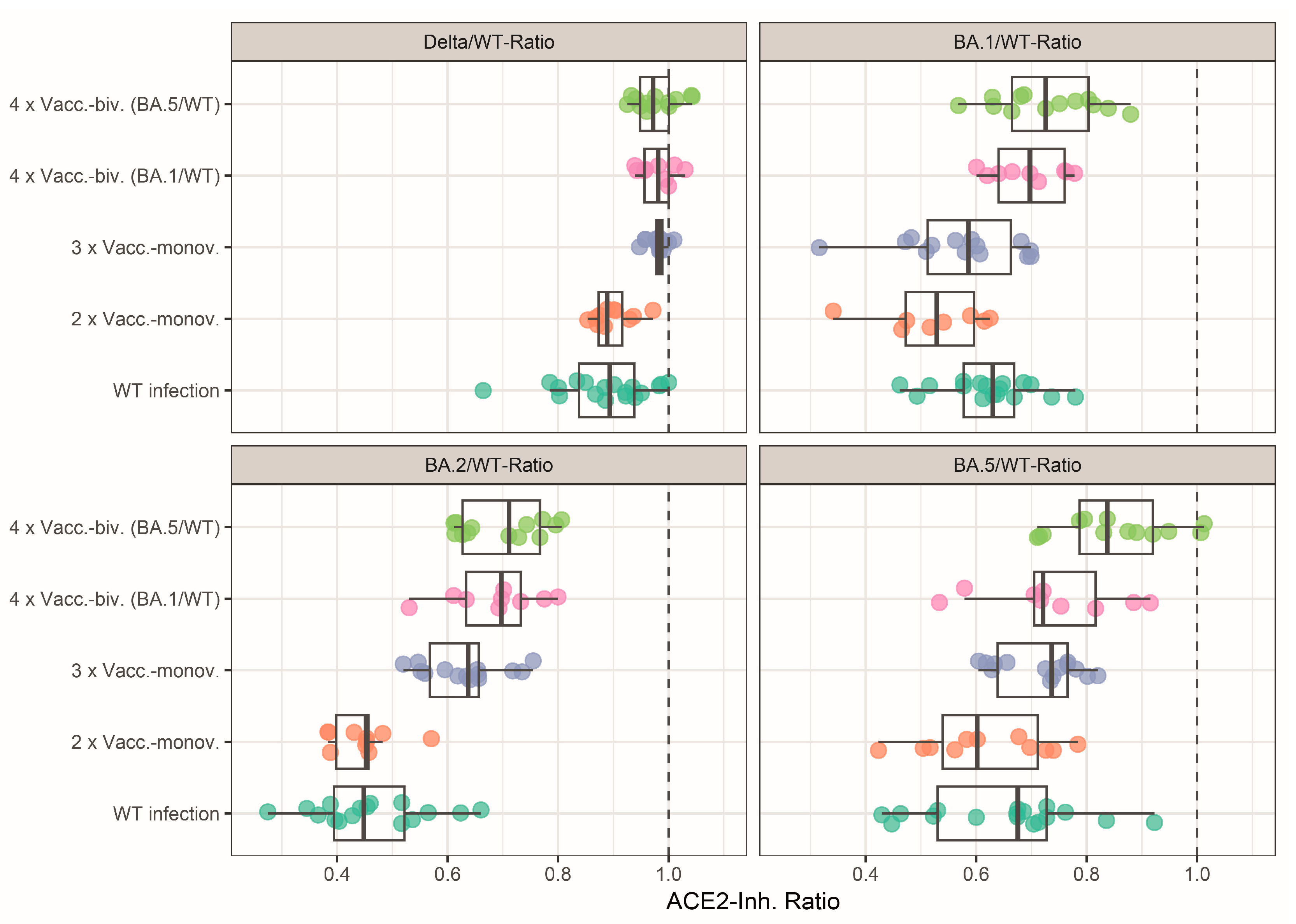
3.3. Effect of Mono- and Bivalent Booster Vaccinations on the Neutralizing Activity
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hachmann, N.P.; Miller, J.; Collier, A.Y.; Ventura, J.D.; Yu, J.; Rowe, M.; Bondzie, E.A.; Powers, O.; Surve, N.; Hall, K.; et al. Neutralization Escape by SARS-CoV-2 Omicron Subvariants BA.2.12.1, BA.4, and BA.5. N. Engl. J. Med. 2022, 387, 86–88. [Google Scholar] [CrossRef]
- Medits, I.; Springer, D.N.; Graninger, M.; Camp, J.V.; Höltl, E.; Aberle, S.W.; Traugott, M.T.; Hoepler, W.; Deutsch, J.; Lammel, O.; et al. Different Neutralization Profiles After Primary SARS-CoV-2 Omicron BA.1 and BA.2 Infections. Front. Immunol. 2022, 13, 946318. [Google Scholar] [CrossRef] [PubMed]
- Rössler, A.; Knabl, L.; von Laer, D.; Kimpel, J. Neutralization Profile after Recovery from SARS-CoV-2 Omicron Infection. N. Engl. J. Med. 2022, 386, 1764–1766. [Google Scholar] [CrossRef] [PubMed]
- Rössler, A.; Netzl, A.; Knabl, L.; Schäfer, H.; Wilks, S.H.; Bante, D.; Falkensammer, B.; Borena, W.; von Laer, D.; Smith, D.J.; et al. BA.2 and BA.5 omicron differ immunologically from both BA.1 omicron and pre-omicron variants. Nat. Commun. 2022, 13, 7701. [Google Scholar] [CrossRef] [PubMed]
- Rössler, A.; Netzl, A.; Knabl, L.; Bante, D.; Wilks, S.H.; Borena, W.; von Laer, D.; Smith, D.J.; Kimpel, J. Characterizing SARS-CoV-2 neutralization profiles after bivalent boosting using antigenic cartography. Nat. Commun. 2023, 14, 5224. [Google Scholar] [CrossRef] [PubMed]
- Springer, D.N.; Bauer, M.; Medits, I.; Camp, J.V.; Aberle, S.W.; Burtscher, C.; Höltl, E.; Weseslindtner, L.; Stiasny, K.; Aberle, J.H. Bivalent COVID-19 mRNA booster vaccination (BA.1 or BA.4/BA.5) increases neutralization of matched Omicron variants. npj Vaccines 2023, 8, 110. [Google Scholar] [CrossRef]
- Dejnirattisai, W.; Huo, J.; Zhou, D.; Zahradník, J.; Supasa, P.; Liu, C.; Duyvesteyn, H.M.E.; Ginn, H.M.; Mentzer, A.J.; Tuekprakhon, A.; et al. SARS-CoV-2 Omicron-B.1.1.529 leads to widespread escape from neutralizing antibody responses. Cell 2022, 185, 467–484.e415. [Google Scholar] [CrossRef] [PubMed]
- Springer, D.N.; Traugott, M.; Reuberger, E.; Kothbauer, K.B.; Borsodi, C.; Nägeli, M.; Oelschlägel, T.; Kelani, H.; Lammel, O.; Deutsch, J.; et al. A Multivariant Surrogate Neutralization Assay Identifies Variant-Specific Neutralizing Antibody Profiles in Primary SARS-CoV-2 Omicron Infection. Diagnostics 2023, 13, 2278. [Google Scholar] [CrossRef]
- Cao, Y.; Yisimayi, A.; Jian, F.; Song, W.; Xiao, T.; Wang, L.; Du, S.; Wang, J.; Li, Q.; Chen, X.; et al. BA.2.12.1, BA.4 and BA.5 escape antibodies elicited by Omicron infection. Nature 2022, 608, 593–602. [Google Scholar] [CrossRef]
- Zaballa, M.E.; Perez-Saez, J.; de Mestral, C.; Pullen, N.; Lamour, J.; Turelli, P.; Raclot, C.; Baysson, H.; Pennacchio, F.; Villers, J.; et al. Seroprevalence of anti-SARS-CoV-2 antibodies and cross-variant neutralization capacity after the Omicron BA.2 wave in Geneva, Switzerland: A population-based study. Lancet Reg. Health Eur. 2023, 24, 100547. [Google Scholar] [CrossRef]
- Santos da Silva, E.; Servais, J.Y.; Kohnen, M.; Arendt, V.; Staub, T.; CON-VINCE Consortium; CoVaLux Consortium; Krüger, R.; Fagherazzi, G.; Wilmes, P.; et al. Validation of a SARS-CoV-2 Surrogate Neutralization Test Detecting Neutralizing Antibodies against the Major Variants of Concern. Int. J. Mol. Sci. 2023, 24, 14965. [Google Scholar] [CrossRef]
- Collier, A.Y.; Miller, J.; Hachmann, N.P.; McMahan, K.; Liu, J.; Bondzie, E.A.; Gallup, L.; Rowe, M.; Schonberg, E.; Thai, S.; et al. Immunogenicity of BA.5 Bivalent mRNA Vaccine Boosters. N. Engl. J. Med. 2023, 388, 565–567. [Google Scholar] [CrossRef] [PubMed]
- Chalkias, S.; Harper, C.; Vrbicky, K.; Walsh, S.R.; Essink, B.; Brosz, A.; McGhee, N.; Tomassini, J.E.; Chen, X.; Chang, Y.; et al. A Bivalent Omicron-Containing Booster Vaccine against COVID-19. N. Engl. J. Med. 2022, 387, 1279–1291. [Google Scholar] [CrossRef] [PubMed]
- Altman, D.G.; Bland, J.M. Measurement in Medicine: The Analysis of Method Comparison Studies. J. R. Stat. Soc. Ser. D Stat. 1983, 32, 307–317. [Google Scholar] [CrossRef]
- Gruell, H.; Vanshylla, K.; Tober-Lau, P.; Hillus, D.; Schommers, P.; Lehmann, C.; Kurth, F.; Sander, L.E.; Klein, F. mRNA booster immunization elicits potent neutralizing serum activity against the SARS-CoV-2 Omicron variant. Nat. Med. 2022, 28, 477–480. [Google Scholar] [CrossRef] [PubMed]
- Quandt, J.; Muik, A.; Salisch, N.; Lui, B.G.; Lutz, S.; Krüger, K.; Wallisch, A.K.; Adams-Quack, P.; Bacher, M.; Finlayson, A.; et al. Omicron BA.1 breakthrough infection drives cross-variant neutralization and memory B cell formation against conserved epitopes. Sci. Immunol. 2022, 7, eabq2427. [Google Scholar] [CrossRef]
- Link-Gelles, R.; Levy, M.E.; Gaglani, M.; Irving, S.A.; Stockwell, M.; Dascomb, K.; DeSilva, M.B.; Reese, S.E.; Liao, I.C.; Ong, T.C.; et al. Effectiveness of 2, 3, and 4 COVID-19 mRNA Vaccine Doses Among Immunocompetent Adults During Periods when SARS-CoV-2 Omicron BA.1 and BA.2/BA.2.12.1 Sublineages Predominated—VISION Network, 10 States, December 2021–June 2022. Morb. Mortal. Wkly. Rep. 2022, 71, 931–939. [Google Scholar] [CrossRef]
- Regev-Yochay, G.; Gonen, T.; Gilboa, M.; Mandelboim, M.; Indenbaum, V.; Amit, S.; Meltzer, L.; Asraf, K.; Cohen, C.; Fluss, R.; et al. Efficacy of a Fourth Dose of COVID-19 mRNA Vaccine against Omicron. N. Engl. J. Med. 2022, 386, 1377–1380. [Google Scholar] [CrossRef]
- Chong, Y.; Goto, T.; Watanabe, H.; Tani, N.; Yonekawa, A.; Ikematsu, H.; Shimono, N.; Tanaka, Y.; Akashi, K. Achievement of sufficient antibody response after a fourth dose of wild-type SARS-CoV-2 mRNA vaccine in nursing home residents. Immun. Inflamm. Dis. 2023, 11, e962. [Google Scholar] [CrossRef]
- Xie, X.; Zou, J.; Kurhade, C.; Liu, M.; Ren, P.; Pei-Yong, S. Neutralization of SARS-CoV-2 Omicron sublineages by 4 doses of the original mRNA vaccine. Cell Rep. 2022, 41, 111729. [Google Scholar] [CrossRef]
- Krammer, F.; Ellebedy, A.H. Variant-adapted COVID-19 booster vaccines. Science 2023, 382, 157–159. [Google Scholar] [CrossRef] [PubMed]
- Carreño, J.M.; Singh, G.; Simon, V.; Krammer, F. Bivalent COVID-19 booster vaccines and the absence of BA.5-specific antibodies. Lancet. Microbe 2023, 8, E569. [Google Scholar] [CrossRef]
- Wang, W.; Lusvarghi, S.; Subramanian, R.; Epsi, N.J.; Wang, R.; Goguet, E.; Fries, A.C.; Echegaray, F.; Vassell, R.; Coggins, S.A.; et al. Antigenic cartography of well-characterized human sera shows SARS-CoV-2 neutralization differences based on infection and vaccination history. Cell Host Microbe 2022, 30, 1745–1758.e7. [Google Scholar] [CrossRef] [PubMed]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Springer, D.N.; Höltl, E.; Prüger, K.; Puchhammer-Stöckl, E.; Aberle, J.H.; Stiasny, K.; Weseslindtner, L. Measuring Variant-Specific Neutralizing Antibody Profiles after Bivalent SARS-CoV-2 Vaccinations Using a Multivariant Surrogate Virus Neutralization Microarray. Vaccines 2024, 12, 94. https://doi.org/10.3390/vaccines12010094
Springer DN, Höltl E, Prüger K, Puchhammer-Stöckl E, Aberle JH, Stiasny K, Weseslindtner L. Measuring Variant-Specific Neutralizing Antibody Profiles after Bivalent SARS-CoV-2 Vaccinations Using a Multivariant Surrogate Virus Neutralization Microarray. Vaccines. 2024; 12(1):94. https://doi.org/10.3390/vaccines12010094
Chicago/Turabian StyleSpringer, David Niklas, Eva Höltl, Katja Prüger, Elisabeth Puchhammer-Stöckl, Judith Helene Aberle, Karin Stiasny, and Lukas Weseslindtner. 2024. "Measuring Variant-Specific Neutralizing Antibody Profiles after Bivalent SARS-CoV-2 Vaccinations Using a Multivariant Surrogate Virus Neutralization Microarray" Vaccines 12, no. 1: 94. https://doi.org/10.3390/vaccines12010094
APA StyleSpringer, D. N., Höltl, E., Prüger, K., Puchhammer-Stöckl, E., Aberle, J. H., Stiasny, K., & Weseslindtner, L. (2024). Measuring Variant-Specific Neutralizing Antibody Profiles after Bivalent SARS-CoV-2 Vaccinations Using a Multivariant Surrogate Virus Neutralization Microarray. Vaccines, 12(1), 94. https://doi.org/10.3390/vaccines12010094