
Binding of Natural Antibodies Generated after COVID-19 and Vaccination with Individual Peptides Corresponding to the SARS-CoV-2 S-Protein

 ,
,  ,
, 
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Oligopeptides
2.2. Immunization of Mice and Blood Collection
2.3. Screening of Antibodies That Recognize the S-Protein Oligopeptides
2.4. Structural Visualization of Oligopeptides
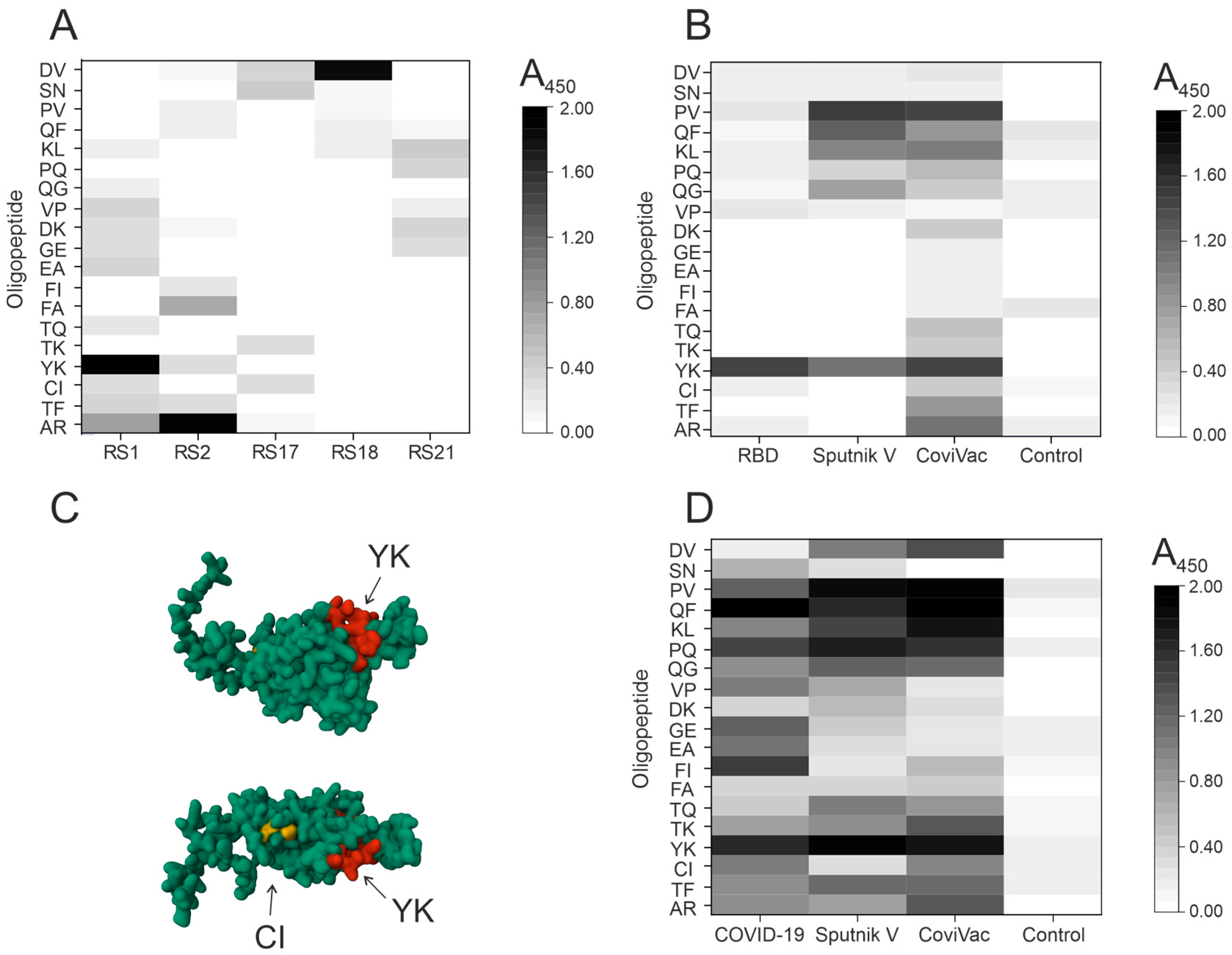
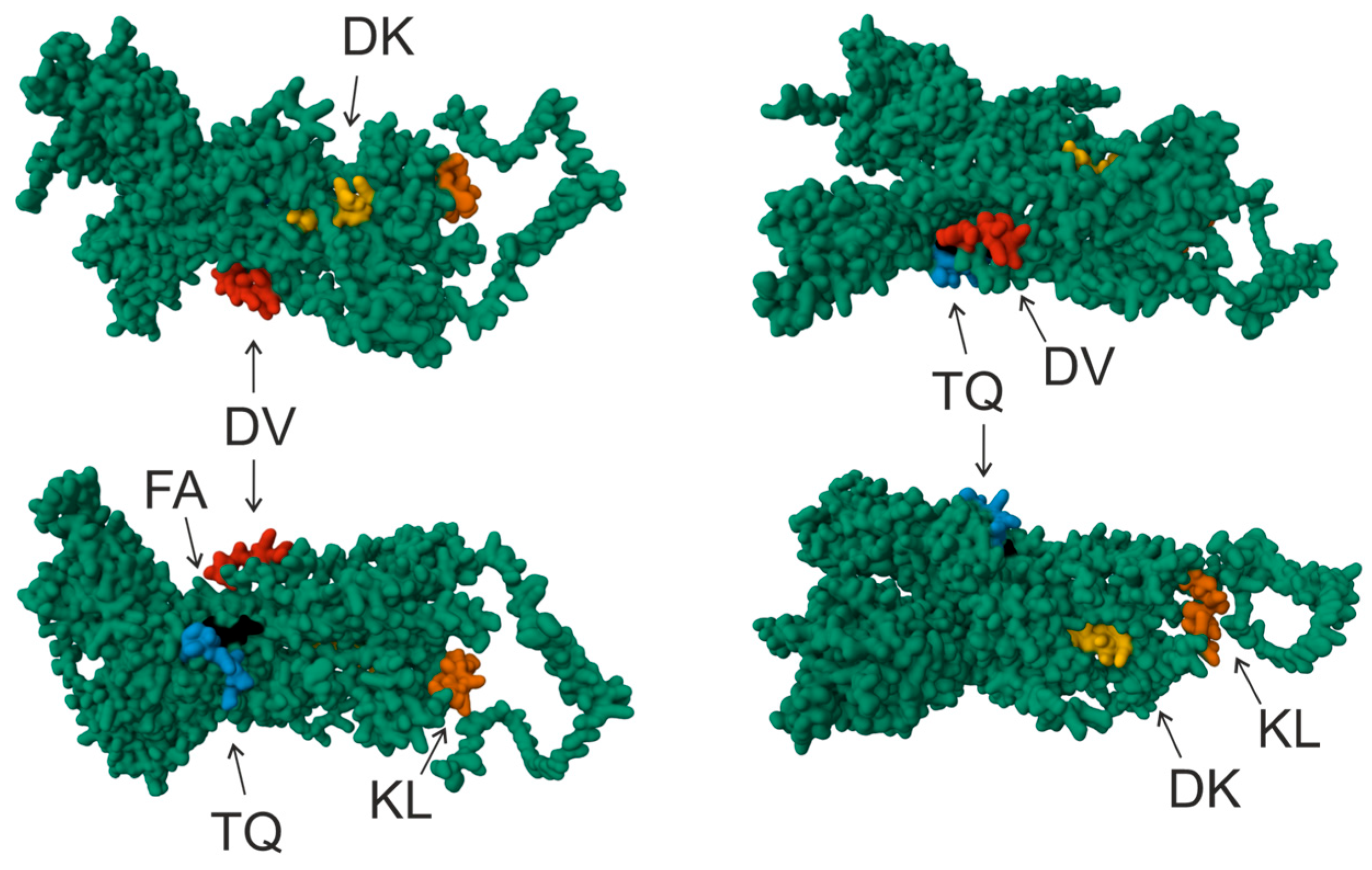
3. Results and Discussion
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zhao, F.; Zai, X.; Zhang, Z.; Xu, J.; Chen, W. Challenges and Developments in Universal Vaccine Design against SARS-CoV-2 Variants. npj Vaccines 2022, 7, 167. [Google Scholar] [CrossRef]
- Dörner, T.; Radbruch, A. Antibodies and B Cell Memory in Viral Immunity. Immunity 2007, 27, 384–392. [Google Scholar] [CrossRef]
- Poh, C.M.; Carissimo, G.; Wang, B.; Amrun, S.N.; Lee, C.Y.-P.; Chee, R.S.-L.; Fong, S.-W.; Yeo, N.K.-W.; Lee, W.-H.; Torres-Ruesta, A.; et al. Two Linear Epitopes on the SARS-CoV-2 Spike Protein That Elicit Neutralising Antibodies in COVID-19 Patients. Nat. Commun. 2020, 11, 2806. [Google Scholar] [CrossRef]
- Mishra, N.; Huang, X.; Joshi, S.; Guo, C.; Ng, J.; Thakkar, R.; Wu, Y.; Dong, X.; Li, Q.; Pinapati, R.S.; et al. Immunoreactive Peptide Maps of SARS-CoV-2. Commun. Biol. 2021, 4, 225. [Google Scholar] [CrossRef]
- Fleri, W.; Paul, S.; Dhanda, S.K.; Mahajan, S.; Xu, X.; Peters, B.; Sette, A. The Immune Epitope Database and Analysis Resource in Epitope Discovery and Synthetic Vaccine Design. Front. Immunol. 2017, 8, 250318. [Google Scholar] [CrossRef]
- Bahai, A.; Asgari, E.; Mofrad, M.R.K.; Kloetgen, A.; McHardy, A.C. EpitopeVec: Linear Epitope Prediction Using Deep Protein Sequence Embeddings. Bioinformatics 2021, 37, 4517–4525. [Google Scholar] [CrossRef]
- Jespersen, M.C.; Peters, B.; Nielsen, M.; Marcatili, P. BepiPred-2.0: Improving Sequence-Based B-Cell Epitope Prediction Using Conformational Epitopes. Nucleic Acids Res. 2017, 45, W24–W29. [Google Scholar] [CrossRef]
- Saha, S.; Raghava, G.P.S. Prediction of Continuous B-cell Epitopes in an Antigen Using Recurrent Neural Network. Proteins Struct. Funct. Bioinform. 2006, 65, 40–48. [Google Scholar] [CrossRef]
- Abbott, W.M.; Damschroder, M.M.; Lowe, D.C. Current Approaches to Fine Mapping of Antigen–Antibody Interactions. Immunology 2014, 142, 526–535. [Google Scholar] [CrossRef] [PubMed]
- Grover, R.K.; Zhu, X.; Nieusma, T.; Jones, T.; Boero, I.; MacLeod, A.S.; Mark, A.; Niessen, S.; Kim, H.J.; Kong, L.; et al. A Structurally Distinct Human Mycoplasma Protein That Generically Blocks Antigen-Antibody Union. Science 2014, 343, 656–661. [Google Scholar] [CrossRef] [PubMed]
- Blech, M.; Peter, D.; Fischer, P.; Bauer, M.M.T.; Hafner, M.; Zeeb, M.; Nar, H. One Target—Two Different Binding Modes: Structural Insights into Gevokizumab and Canakinumab Interactions to Interleukin-1β. J. Mol. Biol. 2013, 425, 94–111. [Google Scholar] [CrossRef] [PubMed]
- Fibriansah, G.; Ibarra, K.D.; Ng, T.-S.; Smith, S.A.; Tan, J.L.; Lim, X.-N.; Ooi, J.S.G.; Kostyuchenko, V.A.; Wang, J.; de Silva, A.M.; et al. Cryo-EM Structure of an Antibody That Neutralizes Dengue Virus Type 2 by Locking E Protein Dimers. Science 2015, 349, 88–91. [Google Scholar] [CrossRef] [PubMed]
- Moraes, I. Erratum to: The Next Generation in Membrane Protein Structure Determination. In The Next Generation in Membrane Protein Structure Determination; Springer: Berlin/Heidelberg, Germany, 2017; p. E1. [Google Scholar]
- Buus, S.; Rockberg, J.; Forsström, B.; Nilsson, P.; Uhlen, M.; Schafer-Nielsen, C. High-Resolution Mapping of Linear Antibody Epitopes Using Ultrahigh-Density Peptide Microarrays. Mol. Cell. Proteom. 2012, 11, 1790–1800. [Google Scholar] [CrossRef] [PubMed]
- Kramer, A.; Keitel, T.; Winkler, K.; Stöcklein, W.; Höhne, W.; Schneider-Mergener, J. Molecular Basis for the Binding Promiscuity of an Anti-P24 (HIV-1) Monoclonal Antibody. Cell 1997, 91, 799–809. [Google Scholar] [CrossRef] [PubMed]
- Petersen, G.; Song, D.; Hügle-Dörr, B.; Oldenburg, I.; Bautz, E.K.F. Mapping of Linear Epitopes Recognized by Monoclonal Antibodies with Gene-Fragment Phage Display Libraries. Mol. Gen. Genet. MGG 1995, 249, 425–431. [Google Scholar] [CrossRef] [PubMed]
- Rentero Rebollo, I.; Sabisz, M.; Baeriswyl, V.; Heinis, C. Identification of Target-Binding Peptide Motifs by High-Throughput Sequencing of Phage-Selected Peptides. Nucleic Acids Res. 2014, 42, e169. [Google Scholar] [CrossRef] [PubMed]
- Paull, M.L.; Johnston, T.; Ibsen, K.N.; Bozekowski, J.D.; Daugherty, P.S. A General Approach for Predicting Protein Epitopes Targeted by Antibody Repertoires Using Whole Proteomes. PLoS ONE 2019, 14, e0217668. [Google Scholar] [CrossRef] [PubMed]
- Belyavtsev, A.N.; Melnikova, M.V.; Shevchenko, N.G.; Sapronov, G.V.; Vakhrenev, R.G.; Shastina, N.S.; Kolesanova, E.F.; Nikolaeva, L.I. Synthesis and Analysis of the Properties of the Immunogenic Fragment of the Hepatitis C Virus NS4A Polypeptide. Bioorg. Chem. 2021, 47, 341–347. [Google Scholar] [CrossRef]
- Chibiskova, O.V.; Kotel’nikova, O.V.; Nesmeyanov, V.A.; Alliluev, A.P.; Vol’pina, O.M.; Koroev, D.O.; Filatova, M.P. Mechanisms of Defense Formation against Meningococcal Infection in Mice Immunized with Synthetic Peptides. Bull. Exp. Biol. Med. 2007, 143, 720–722. [Google Scholar] [CrossRef]
- Lan, J.; Ge, J.; Yu, J.; Shan, S.; Zhou, H.; Fan, S.; Zhang, Q.; Shi, X.; Wang, Q.; Zhang, L.; et al. Structure of the SARS-CoV-2 Spike Receptor-Binding Domain Bound to the ACE2 Receptor. Nature 2020, 581, 215–220. [Google Scholar] [CrossRef]
- Logunov, D.Y.; Livermore, D.M.; Ornelles, D.A.; Bayer, W.; Marques, E.; Czerkinsky, C.; Dolzhikova, I.V.; Ertl, H.C. COVID-19 Vaccination and HIV-1 Acquisition. Lancet 2022, 399, e34–e35. [Google Scholar] [CrossRef]
- Balakrishnan, V.S. The Arrival of Sputnik V. Lancet Infect. Dis. 2020, 20, 1128. [Google Scholar] [CrossRef] [PubMed]
- Kozlovskaya, L.I.; Piniaeva, A.N.; Ignatyev, G.M.; Gordeychuk, I.V.; Volok, V.P.; Rogova, Y.V.; Shishova, A.A.; Kovpak, A.A.; Ivin, Y.Y.; Antonova, L.P.; et al. Long-Term Humoral Immunogenicity, Safety and Protective Efficacy of Inactivated Vaccine against COVID-19 (CoviVac) in Preclinical Studies. Emerg. Microbes Infect. 2021, 10, 1790–1806. [Google Scholar] [CrossRef]
- Farrera-Soler, L.; Daguer, J.-P.; Barluenga, S.; Vadas, O.; Cohen, P.; Pagano, S.; Yerly, S.; Kaiser, L.; Vuilleumier, N.; Winssinger, N. Identification of Immunodominant Linear Epitopes from SARS-CoV-2 Patient Plasma. PLoS ONE 2020, 15, e0238089. [Google Scholar] [CrossRef]
- Bateman, A.; Martin, M.-J.; Orchard, S.; Magrane, M.; Ahmad, S.; Alpi, E.; Bowler-Barnett, E.H.; Britto, R.; Bye-A-Jee, H.; Cukura, A.; et al. UniProt: The Universal Protein Knowledgebase in 2023. Nucleic Acids Res. 2023, 51, D523–D531. [Google Scholar] [CrossRef]
- Mirdita, M.; Schütze, K.; Moriwaki, Y.; Heo, L.; Ovchinnikov, S.; Steinegger, M. ColabFold: Making Protein Folding Accessible to All. Nat. Methods 2022, 19, 679–682. [Google Scholar] [CrossRef] [PubMed]
- Sehnal, D.; Bittrich, S.; Deshpande, M.; Svobodová, R.; Berka, K.; Bazgier, V.; Velankar, S.; Burley, S.K.; Koča, J.; Rose, A.S. Mol* Viewer: Modern Web App for 3D Visualization and Analysis of Large Biomolecular Structures. Nucleic Acids Res. 2021, 49, W431–W437. [Google Scholar] [CrossRef] [PubMed]
- Szymczak, L.C.; Kuo, H.-Y.; Mrksich, M. Peptide Arrays: Development and Application. Anal. Chem. 2018, 90, 266–282. [Google Scholar] [CrossRef]
- Verastegui, M.; Moro, P.; Guevara, A.; Rodriguez, T.; Miranda, E.; Gilman, R.H. Enzyme-Linked Immunoelectrotransfer Blot Test for Diagnosis of Human Hydatid Disease. J. Clin. Microbiol. 1992, 30, 1557–1561. [Google Scholar] [CrossRef]
- Blake, M.S.; Johnston, K.H.; Russell-Jones, G.J.; Gotschlich, E.C. A Rapid, Sensitive Method for Detection of Alkaline Phosphatase-Conjugated Anti-Antibody on Western Blots. Anal. Biochem. 1984, 136, 175–179. [Google Scholar] [CrossRef]
- Tai, C.; Li, H.; Zhang, J. BCEDB: A Linear B-Cell Epitopes Database for SARS-CoV-2. Database 2023, 2023, baad065. [Google Scholar] [CrossRef]
- Matveev, A.L.; Pyankov, O.V.; Khlusevich, Y.A.; Tyazhelkova, O.V.; Emelyanova, L.A.; Timofeeva, A.M.; Shipovalov, A.V.; Chechushkov, A.V.; Zaitseva, N.S.; Kudrov, G.A.; et al. Novel B-Cell Epitopes of Non-Neutralizing Antibodies in the Receptor-Binding Domain of the SARS-CoV-2 S-Protein with Different Effects on the Severity of COVID-19. Biochemistry 2023, 88, 1205–1214. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Y.; Wei, Z.; Yuan, Q.; Chen, S.; Yu, W.; Lu, Y.; Gao, J.; Yang, Y. Identifying B-Cell Epitopes Using AlphaFold2 Predicted Structures and Pretrained Language Model. Bioinformatics 2023, 39, btad187. [Google Scholar] [CrossRef] [PubMed]
- McCoy, A.J.; Sammito, M.D.; Read, R.J. Implications of AlphaFold 2 for Crystallographic Phasing by Molecular Replacement. Acta Crystallogr. Sect. D Struct. Biol. 2022, 78, 1–13. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Oligopeptide Designation | Amino Acid Residues in the SARS-CoV-2 S-Protein | Sequence of the Oligopeptide Amino Acid Residues |
|---|---|---|
| AR | 67–78 | AIHVSGTNGTKR |
| TF | 307–318 | TVEKGIYQTSNF |
| CI | 391–402 | CFTNVYADSFVI |
| YK | 451–462 | YLYRLFRKSNLK |
| TK | 547–558 | TGTGVLTESNKK |
| TQ | 553–564 | TESNKKFLPFQQ |
| FA | 559–570 | FLPFQQFGRDIA |
| FI | 655–666 | FVNNSYECDIPI |
| EA | 661–672 | ECDIPIGAGICA |
| GE | 769–780 | GIAVEQDKNTQE |
| DK | 775–786 | DKNTQEVFAQVK |
| VP | 781–792 | VFAQVKQIYKTP |
| QG | 787–798 | QIYKTPPIKDFG |
| PQ | 793–804 | PIKDFGGFNFSQ |
| KL | 811–822 | KPSKRSFIEDLL |
| QF | 895–906 | QIPFAMQMAYRF |
| PV | 1057–1068 | PHGVVFLHVTYV |
| SN | 1147–1158 | SFKEELDKYFKN |
| DV | 1153–1164 | DKYFKNHTSPDV |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Timofeeva, A.M.; Sedykh, S.E.; Litvinova, E.A.; Dolgushin, S.A.; Matveev, A.L.; Tikunova, N.V.; Nevinsky, G.A. Binding of Natural Antibodies Generated after COVID-19 and Vaccination with Individual Peptides Corresponding to the SARS-CoV-2 S-Protein. Vaccines 2024, 12, 426. https://doi.org/10.3390/vaccines12040426
Timofeeva AM, Sedykh SE, Litvinova EA, Dolgushin SA, Matveev AL, Tikunova NV, Nevinsky GA. Binding of Natural Antibodies Generated after COVID-19 and Vaccination with Individual Peptides Corresponding to the SARS-CoV-2 S-Protein. Vaccines. 2024; 12(4):426. https://doi.org/10.3390/vaccines12040426
Chicago/Turabian StyleTimofeeva, Anna M., Sergey E. Sedykh, Ekaterina A. Litvinova, Sergey A. Dolgushin, Andrey L. Matveev, Nina V. Tikunova, and Georgy A. Nevinsky. 2024. "Binding of Natural Antibodies Generated after COVID-19 and Vaccination with Individual Peptides Corresponding to the SARS-CoV-2 S-Protein" Vaccines 12, no. 4: 426. https://doi.org/10.3390/vaccines12040426
APA StyleTimofeeva, A. M., Sedykh, S. E., Litvinova, E. A., Dolgushin, S. A., Matveev, A. L., Tikunova, N. V., & Nevinsky, G. A. (2024). Binding of Natural Antibodies Generated after COVID-19 and Vaccination with Individual Peptides Corresponding to the SARS-CoV-2 S-Protein. Vaccines, 12(4), 426. https://doi.org/10.3390/vaccines12040426