
Development of a Candidate TMV Epitope Display Vaccine against SARS-CoV-2

Abstract
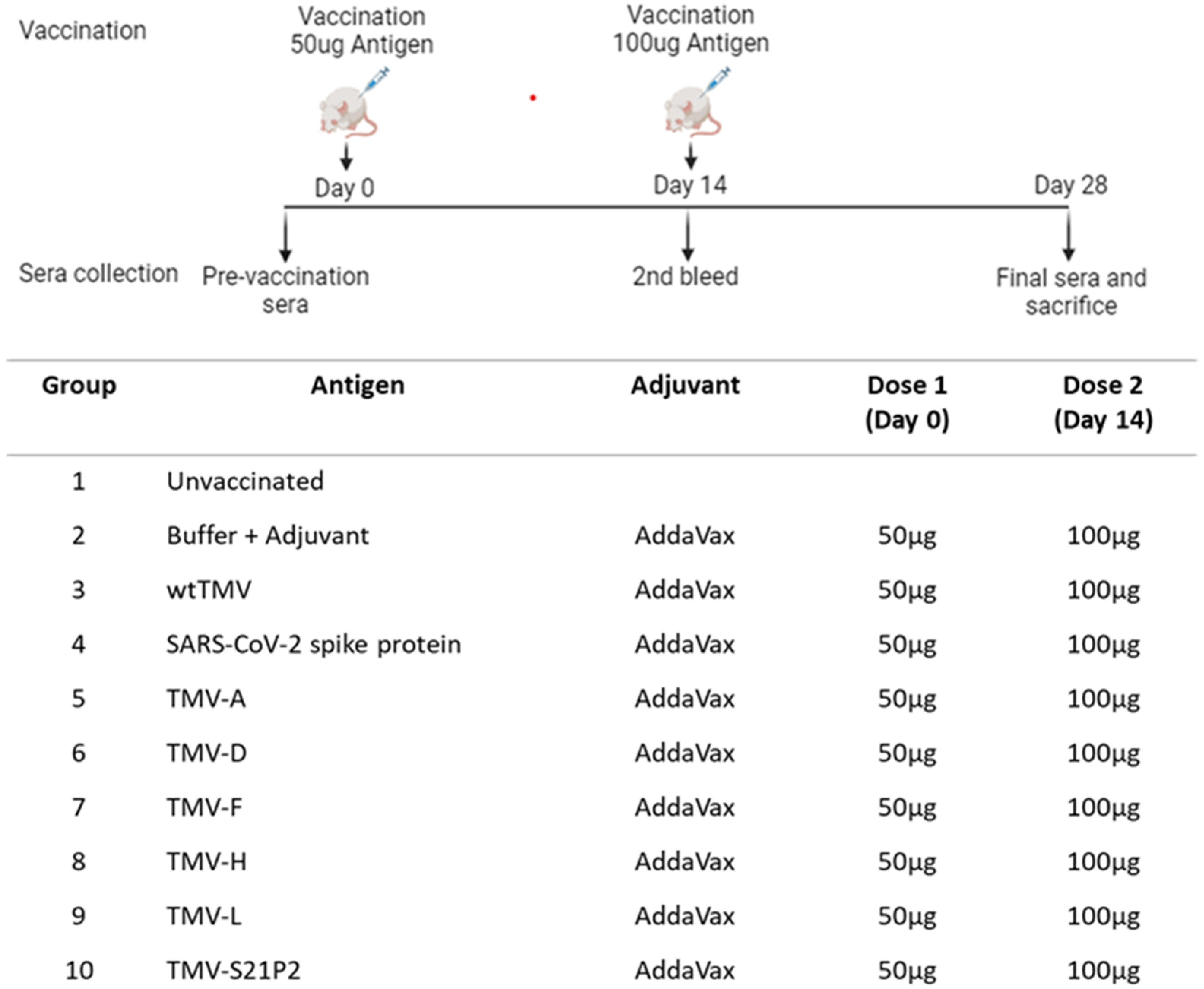
:1. Introduction
2. Methods
2.1. Peptide Design
2.2. TMV Expression and Virus Purification
2.3. DLS Measurements on Viral Samples
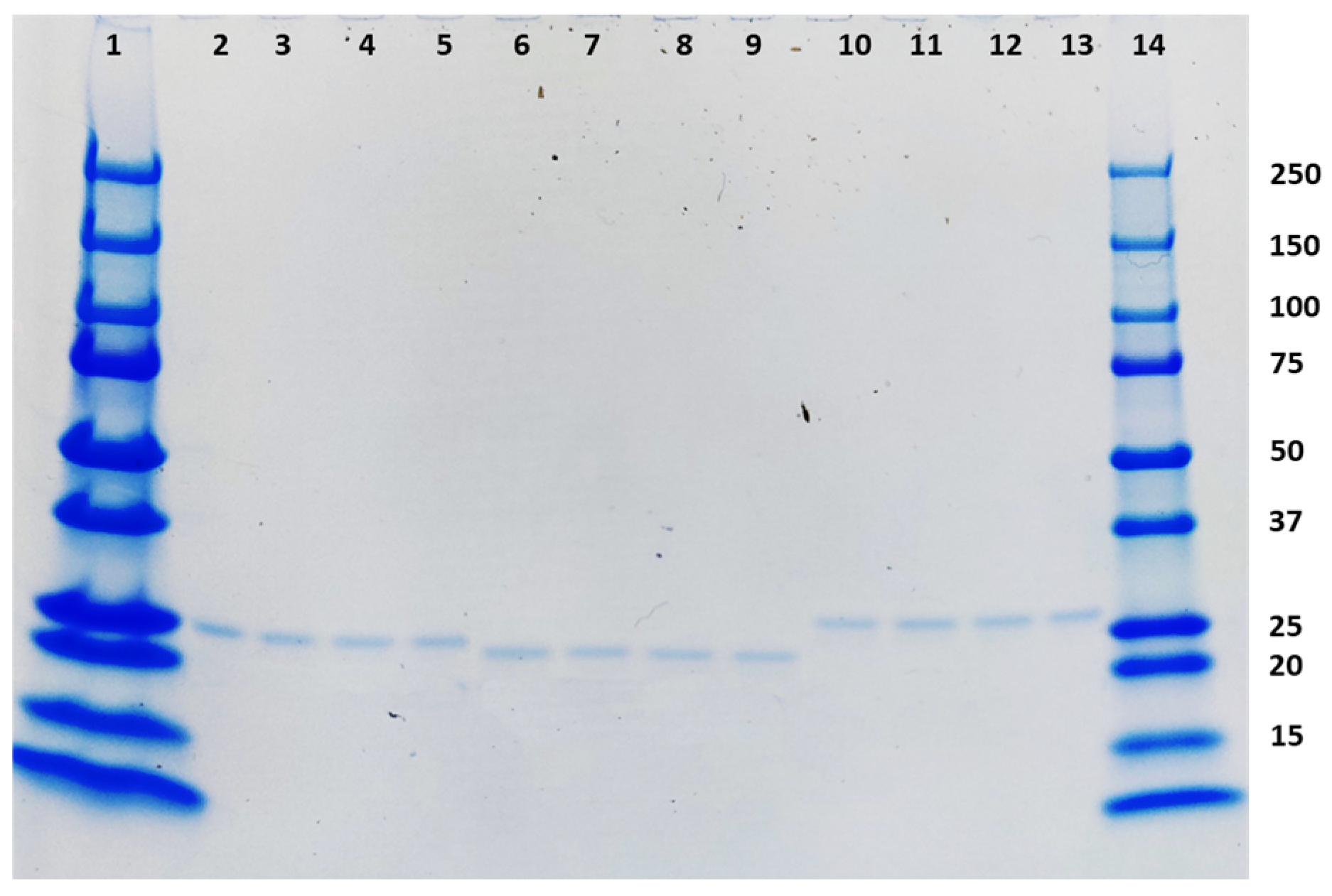
2.4. SDS-PAGE, Western Blot, and ELISA
2.5. Antigenicity Study
2.6. IFN-γ Detection with ELISA
2.7. Neutralization
2.8. Thermal Stability
3. Results
3.1. TMV Expression and Virus Purification
3.2. Antigenicity Study
3.2.1. Vaccine Immunogenicity
3.2.2. IFN-γ Detection
3.2.3. Neutralization
3.2.4. Thermal Stability
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Woo, P.C.Y.; De Groot, R.J.; Haagmans, B.; Lau, S.K.P.; Neuman, B.W.; Perlman, S.; Sola, I.; Van Der Hoek, L.; Wong, A.C.P.; Yeh, S.H. ICTV Virus Taxonomy Profile: Coronaviridae 2023. J. Gen. Virol. 2023, 104, 001843. [Google Scholar] [CrossRef]
- Philip, V. Coronavirus biology and replication: Implications for SARS-CoV-2. Nat. Rev. Microbiol. 2021, 19, 155–170. [Google Scholar] [CrossRef] [PubMed]
- Petrosillo, N.; Viceconte, G.; Ergonul, O.; Ippolito, G.; Petersen, E. COVID-19, SARS and MERS: Are they closely related? Clin. Microbiol. Infect. 2020, 26, 729–734. [Google Scholar] [CrossRef]
- Johns Hopkins University. COVID-19 Dashboard by the Center for Systems Science and Engineering (CSSE) at Johns Hopkins University (JHU). 2023. Available online: https://coronavirus.jhu.edu/map.html (accessed on 16 April 2024).
- Aleem, A.; Akbar Samad, A.B.; Vaqar, S. Emerging Variants of SARS-CoV-2 and Novel Therapeutics against Coronavirus (COVID-19); StatPearls Publishing: Treasure Island, FL, USA, 2023. [Google Scholar]
- Yi, C.; Sun, X.; Ling, Z.; Sun, B. Jigsaw puzzle of SARS-CoV-2 RBD evolution and immune escape. Cell. Mol. Immunol. 2022, 19, 848–851. [Google Scholar] [CrossRef] [PubMed]
- Gregory, A.E.; Titball, R.; Williamson, D.; Morici, L.A. Vaccine delivery using nanoparticles. Front. Cell. Infect. Microbiol. 2013, 3, 13. [Google Scholar] [CrossRef] [PubMed]
- Mohammad, S.; Hayat, G. Nanovaccine: A novel approach in immunization. J. Cell. Physiol. 2019, 234, 12530–12536. [Google Scholar] [CrossRef] [PubMed]
- McCormick, A.A. Tobacco derived cancer vaccines for non-Hodgkin’s lymphoma: Perspectives and progress. Hum. Vaccin. 2011, 7, 305–312. [Google Scholar] [CrossRef] [PubMed]
- Langowski, M.D.; Khan, F.A.; Bitzer, A.A.; Genito, C.J.; Schrader, A.J.; Martin, M.L.; Soto, K.; Zou, X.; Hadiwidjojo, S.; Beck, Z.; et al. Optimization of a Plasmodium falciparum circumsporozoite protein repeat vaccine using the tobacco mosaic virus platform. Proc. Natl. Acad. Sci. USA 2020, 117, 3114–3122. [Google Scholar] [CrossRef] [PubMed]
- Turpen, T.H.; Reinl, S.J.; Charoenvit, Y.; Hoffman, S.L.; Fallarme, V.; Grill, L.K. Malarial epitopes expressed on the surface of recombinant tobacco mosaic virus. Biotechnology 1995, 13, 53–57. [Google Scholar] [CrossRef] [PubMed]
- Steele, J.F.C.; Peyret, H.; Saunders, K.; Castells-graells, R.; Marsian, J.; Meshcheriakova, Y.; Lomonossoff, G.P. Synthetic plant virology for nanobiotechnology and nanomedicine. Wiley Interdiscip. Rev. Nanomed. Nanobiotechnol. 2017, 9, e1447. [Google Scholar] [CrossRef] [PubMed]
- McCormick, A.A.; Corbo, T.A.; Wykoff-Clary, S.; Palmer, K.E.; Pogue, G.P. Chemical conjugate TMV—Peptide bivalent fusion vaccines improve cellular immunity and tumor protection. Bioconjug. Chem. 2006, 17, 1330–1338. [Google Scholar] [CrossRef] [PubMed]
- Rybicki, E.P. Plant-made vaccines for humans and animals. Plant Biotechnol. J. 2010, 8, 620–637. [Google Scholar] [CrossRef] [PubMed]
- Pillet, S.; Aubin, É.; Trépanier, S.; Bussière, D.; Dargis, M.; Poulin, J.F.; Yassine-Diab, B.; Ward, B.J.; Landry, N. A plant-derived quadrivalent virus like particle influenza vaccine induces cross-reactive antibody and T cell response in healthy adults. Clin. Immunol. 2016, 168, 72–87. [Google Scholar] [CrossRef] [PubMed]
- Pillet, S.; Couillard, J.; Trépanier, S.; Poulin, J.F.; Yassine-Diab, B.; Guy, B.; Ward, B.J.; Landry, N. Immunogenicity and safety of a quadrivalent plant-derived virus like particle influenza vaccine candidate—Two randomized Phase II clinical trials in 18 to 49 and 50 years old adults. PLoS ONE 2019, 14, e0216533. [Google Scholar] [CrossRef] [PubMed]
- Evtushenko, E.A.; Ryabchevskaya, E.M.; Nikitin, N.A.; Atabekov, J.G.; Karpova, O.V. Plant virus particles with various shapes as potential adjuvants. Sci. Rep. 2020, 10, 10365. [Google Scholar] [CrossRef] [PubMed]
- Smith, M.L.; Lindbo, J.A.; Dillard-Telm, S.; Brosio, P.M.; Lasnik, A.B.; McCormick, A.A.; Nguyen, L.V.; Palmer, K.E. Modified tobacco mosaic virus particles as scaffolds for display of protein antigens for vaccine applications. Virology 2006, 348, 475–488. [Google Scholar] [CrossRef] [PubMed]
- Werner, S.; Marillonnet, S.; Hause, G.; Klimyuk, V.; Gleba, Y. Immunoabsorbent nanoparticles based on a tobamovirus displaying protein A. Proc. Natl. Acad. Sci. USA 2006, 103, 17678–17683. [Google Scholar] [CrossRef] [PubMed]
- Kemnade, J.O.; Seethammagari, M.; Collinson-Pautz, M.; Kaur, H.; Spencer, D.M.; McCormick, A.A. Tobacco mosaic virus efficiently targets DC uptake, activation and antigen-specific T cell responses in vivo. Vaccine 2014, 32, 4228–4233. [Google Scholar] [CrossRef] [PubMed]
- McCormick, A.A.; Palmer, K.E. Genetically engineered Tobacco mosaic virus as nanoparticle vaccines. Expert Rev. Vaccines 2008, 7, 33–41. [Google Scholar] [CrossRef] [PubMed]
- McCormick, A.A.; Corbo, T.A.; Wykoff-Clary, S.; Nguyen, L.V.; Smith, M.L.; Palmer, K.E.; Pogue, G.P. TMV-peptide fusion vaccines induce cell-mediated immune responses and tumor protection in two murine models. Vaccine 2006, 24, 6414–6423. [Google Scholar] [CrossRef] [PubMed]
- Wu, L. Expression of foot-and-mouth disease virus epitopes in tobacco by a tobacco mosaic virus-based vector. Vaccine 2003, 21, 4390–4398. [Google Scholar] [CrossRef] [PubMed]
- Chandler, G.L. Influenza Hemagglutinin Expression in Nicotiana tabacum and Nicotiana benthamiana. Ph.D. Thesis, Baylor University, Waco, TX, USA, 2007. [Google Scholar]
- Chakraborti, S.; Prabakaran, P.; Xiao, X.; Dimitrov, D.S. The SARS coronavirus S glycoprotein receptor binding domain: Fine mapping and functional characterization. Virol. J. 2005, 2, 73. [Google Scholar] [CrossRef] [PubMed]
- Poh, C.M.; Carissimo, G.; Wang, B.; Amrun, S.N.; Lee, C.Y.P.; Chee, R.S.L.; Fong, S.W.; Yeo, N.K.W.; Lee, W.H.; Torres-Ruesta, A.; et al. Two linear epitopes on the SARS-CoV-2 spike protein that elicit neutralising antibodies in COVID-19 patients. Nat. Commun. 2020, 11, 2806. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, S.F.; Quadeer, A.A. Preliminary Identification of Potential Vaccine Targets for the COVID-19 Coronavirus (SARS-CoV-2) Based on SARS-CoV Immunological Studies. Viruses 2020, 12, 254. [Google Scholar] [CrossRef]
- Maghsood, F.; Shokri, M.R.; Jeddi-Tehrani, M.; Rahvar, M.T.; Ghaderi, A.; Salimi, V.; Kardar, G.A.; Zarnani, A.H.; Amiri, M.M.; Shokri, F. Identification of immunodominant epitopes on nucleocapsid and spike proteins of the SARS-CoV-2 in Iranian COVID-19 patients. Pathog. Dis. 2022, 80, ftac001. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Li, J.; Du, L.; Yan, X.; Hu, G.; Zhou, Y.; Jiang, S. Identification and characterization of novel neutralizing epitopes in the receptor-binding domain of SARS-CoV spike protein: Revealing the critical antigenic determinants in inactivated SARS-CoV vaccine. Vaccine 2006, 24, 5498–5508. [Google Scholar] [CrossRef] [PubMed]
- Tarigan, S.; Dharmayanti, N.L.P.I.; Sugiartanti, D.; Putri, R.; Andriani; Nuradji, H.; Robinson, M.; Wiendayanthi, N.; Djufri, F. Characterization of two linear epitopes SARS CoV-2 spike protein formulated in tandem repeat. PLoS ONE 2023, 18, e0280627. [Google Scholar] [CrossRef] [PubMed]
- Lindbo, J.A. TRBO: A High-Efficiency Tobacco Mosaic Virus. Breakthr. Technol. 2007, 145, 1232–1240. [Google Scholar] [CrossRef] [PubMed]
- Henderson, E.A.; Tam, C.C.; Cheng, L.W.; Ngono, A.E.; Nguyen, A.V.; Shresta, S.; McGee, M.; Padgett, H.; Grill, L.K.; Martchenko Shilman, M. Investigation of the immunogenicity of Zika glycan loop. Virol. J. 2020, 17, 43. [Google Scholar] [CrossRef]
- Chapman, S. Tobamovirus isolation and RNA extraction. In Plant Virology Protocols; Foster, G., Taylor, S., Eds.; Humana Press Inc.: Totowa, NJ, USA, 1998; pp. 123–130. [Google Scholar]
- Bruckman, M.A.; Steinmetz, N.F. Chemical Modification of the Inner and Outer Surfaces of Tobacco Mosaic Virus (TMV). Virus Hybrids Nanomater. Methods Protoc. 2014, 1108, 173–185. [Google Scholar] [CrossRef] [PubMed]
- Love, A.J.; Makarov, V.V.; Sinitsyna, O.V.; Shaw, J.; Yaminsky, I.V.; Kalinina, N.O.; Taliansky, M.E. A genetically modified tobacco mosaic virus that can produce gold nanoparticles from a metal salt precursor. Front. Plant Sci. 2015, 6, 984. [Google Scholar] [CrossRef] [PubMed]
- McComb, R.C.; Ho, C.; Bradley, K.A.; Grill, L.K.; Martchenko, M. Presentation of peptides from Bacillus anthracis protective antigen on Tobacco Mosaic Virus as an epitope targeted anthrax vaccine. Vaccine 2015, 33, 6745–6751. [Google Scholar] [CrossRef]
- Ferrara, F.; Temperton, N. Pseudotype neutralization assays: From laboratory bench to data analysis. Methods Protoc. 2018, 1, 8. [Google Scholar] [CrossRef] [PubMed]
- Johnson, B.A.; Xie, X.; Bailey, A.L.; Kalveram, B.; Lokugamage, K.G.; Muruato, A.; Zou, J.; Zhang, X.; Juelich, T.; Smith, J.K.; et al. Loss of furin cleavage site attenuates SARS-CoV-2 pathogenesis. Nature 2021, 591, 293–299. [Google Scholar] [CrossRef] [PubMed]
- Bobrovnik, S.A. Determination of antibody affinity by ELISA. Theory. J. Biochem. Biophys. Methods 2003, 57, 213–236. [Google Scholar] [CrossRef] [PubMed]
- Rosa, S.S.; Prazeres, D.M.F.; Azevedo, A.M.; Marques, M.P.C. mRNA vaccines manufacturing: Challenges and bottlenecks. Vaccine 2021, 39, 2190–2200. [Google Scholar] [CrossRef] [PubMed]
- Bendahmane, M.; Koo, M.; Karrer, E.; Beachy, R.N. Display of epitopes on the surface of tobacco mosaic virus: Impact of charge and isoelectric point of the epitope on virus-host interactions. J. Mol. Biol. 1999, 290, 9–20. [Google Scholar] [CrossRef] [PubMed]
- Steinmetz, N.F.; Evans, D.J. Utilisation of plant viruses in bionanotechnology. Org. Biomol. Chem. 2007, 5, 2891–2902. [Google Scholar] [CrossRef] [PubMed]
- Lebel, M.-È.; Chartrand, K.; Leclerc, D.; Lamarre, A. Plant Viruses as Nanoparticle-Based Vaccines and Adjuvants. Vaccines 2015, 3, 620–637. [Google Scholar] [CrossRef] [PubMed]
- Kalkeri, R.; Cai, Z.; Lin, S.; Farmer, J.; Kuzmichev, Y.V.; Koide, F. SARS-CoV-2 spike pseudoviruses: A useful tool to study virus entry and address emerging neutralization escape phenotypes. Microorganisms 2021, 9, 1744. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Zhang, X.E. Construction and applications of SARS-CoV-2 pseudoviruses: A mini review. Int. J. Biol. Sci. 2021, 17, 1574–1580. [Google Scholar] [CrossRef] [PubMed]
- Royal, J.M.; Simpson, C.A.; McCormick, A.A.; Phillips, A.; Hume, S.; Morton, J.; Shepherd, J.; Oh, Y.; Swope, K.; Debeauchamp, J.L.; et al. Development of a SARS-CoV-2 vaccine candidate using plant-based manufacturing and a tobacco mosaic virus-like nano-particle. Vaccines 2021, 9, 1347. [Google Scholar] [CrossRef] [PubMed]
- Aggarwal, A.; Mehta, S.; Gupta, D.; Sheikh, S.; Pallagatti, S.; Singh, R.; Singla, I. Safety & effectiveness of COVID-19 vaccines: A narrative review. Indian J. Med. Res. 2022, 155, 91–104. [Google Scholar]
- Rahman, M.M.; Masum, M.H.U.; Wajed, S.; Talukder, A. A comprehensive review on COVID-19 vaccines: Development, effectiveness, adverse effects, distribution and challenges. VirusDisease 2022, 33, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Tsai, T.I.; Khalili, J.S.; Gilchrist, M.; Waight, A.B.; Cohen, D.; Zhuo, S.; Zhang, Y.; Ding, M.; Zhu, H.; Mak, A.N.S.; et al. ACE2-Fc fusion protein overcomes viral escape by potently neutralizing SARS-CoV-2 variants of concern. Antivir. Res. 2022, 199, 105271. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.; Irmler, M.; Keshavan, S.; Introna, M.; Beckers, J.; Palmberg, L.; Johanson, G.; Ganguly, K.; Upadhyay, S. Differential Effect of SARS-CoV-2 Spike Glycoprotein 1 on Human Bronchial and Alveolar Lung Mucosa Models: Implications for Pathogenicity. Viruses 2021, 13, 2537. [Google Scholar] [CrossRef] [PubMed]
- Banik, S.; Mansour, A.A.; Suresh, R.V.; Wykoff-Clary, S.; Malik, M.; McCormick, A.A.; Bakshi, C.S. Development of a Multivalent Subunit Vaccine against Tularemia Using Tobacco Mosaic Virus (TMV) Based Delivery System. PLoS ONE 2015, 10, e0130858. [Google Scholar] [CrossRef] [PubMed]
- Shivprasad, S.; Pogue, G.P.; Lewandowski, D.J.; Hidalgo, J.; Donson, J.; Grill, L.K.; Dawson, W.O. Heterologous sequences greatly affect foreign gene expression in tobacco mosaic virus-based vectors. Virology 1999, 255, 312–323. [Google Scholar] [CrossRef] [PubMed]
- Leav, B.A.; Yoshida, M.; Rogers, K.; Cohen, S.; Godiwala, N.; Blumberg, R.S.; Ward, H. An early intestinal mucosal source of gamma interferon is associated with resistance to and control of Cryptospondium parvum infection in mice. Infect. Immun. 2005, 73, 8425–8428. [Google Scholar] [CrossRef] [PubMed]
- Caccamo, N.; Guggino, G.; Joosten, S.A.; Gelsomino, G.; Di Carlo, P.; Titone, L.; Galati, D.; Bocchino, M.; Matarese, A.; Salerno, A.; et al. Multifunctional CD4+ T cells correlate with active Mycobacterium tuberculosis infection. Eur. J. Immunol. 2010, 40, 2211–2220. [Google Scholar] [CrossRef] [PubMed]
- Sedegah, M.; Porter, C.; Goguet, E.; Ganeshan, H.; Belmonte, M.; Huang, J.; Belmonte, A.; Inoue, S.; Acheampong, N.; Malloy, A.M.W.; et al. Cellular interferon-gamma and interleukin-2 responses to SARS-CoV-2 structural proteins are broader and higher in those vaccinated after SARS-CoV-2 infection compared to vaccinees without prior SARS-CoV-2 infection. PLoS ONE 2022, 17, e0276241. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Yuen, T.T.T.; Dou, Y.; Hu, J.; Li, R.; Zeng, Z.; Lin, X.; Gong, H.; Chan, C.H.C.; Yoon, C.; et al. Vaccine-induced protection against SARS-CoV-2 requires IFN-γ-driven cellular immune response. Nat. Commun. 2023, 14, 3440. [Google Scholar] [CrossRef] [PubMed]
- Sukhikh, G.T.; Priputnevich, T.V.; Ogarkova, D.A.; Pochtovyi, A.A.; Kustova, D.D.; Zlobin, V.I.; Logunov, D.Y.; Gushchin, V.A.; Gintsburg, A.L. Sputnik Light and Sputnik V Vaccination Is Effective at Protecting Medical Personnel from COVID-19 during the Period of Delta Variant Dominance. Vaccines 2022, 10, 1804. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Chen, X. CpG 1018 Is an Effective Adjuvant for Influenza Nucleoprotein. Vaccines 2023, 11, 649. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Position | Epitope Sequence | Mass (kDa) | |
|---|---|---|---|---|
| Epitope | Epitope + TMV CP | |||
| A | 300–330 | KCTLKSFTVEKGIYQTSNFRVQPTESIVRFP | 6.29 | 24.19 |
| B | 365–395 | YSVLYNSASFSTFKCYGVSPTKLNDLCFTNV | 4.16 | 22.06 |
| C | 420–440 | DYNYKLPDDFTGCVIAWNSNN | 3.05 | 20.95 |
| D | 435–480 | AWNSNNLDSKVGGNYNYLYRLFRKSNLKPFERDISTEIYQAGSTPC | 6.02 | 23.92 |
| E | 420–500 | DYNYKLPDDFTGCVIAWNSNNLDSKVGGNYNYLYRLFRKSNLKPFERDISTEIYQAGSTPCNGVEGFNCYFPLQSYGFQPT | 10.01 | 27.91 |
| F | 440–500 | NLDSKVGGNYNYLYRLFRKSNLKPFERDISTEIYQAGSTPCNGVEGFNCYFPLQSYGFQPT | 7.69 | 25.59 |
| G | 420–540 | DYNYKLPDDFTGCVIAWNSNNLDSKVGGNYNYLYRLFRKSNLKPFERDISTEIYQAGSTPCNGVEGFNCYFPLQSYGFQPTNGVGYQPYRVVVLSFELLHAPATVCGPKKSTNLVKNKCVN | 14.33 | 32.23 |
| H | 481–540 | NGVEGFNCYFPLQSYGFQPTNGVGYQPYRVVVLSFELLHAPATVCGPKKSTNLVKNKCVN | 7.28 | 25.18 |
| I | 475–500 | AGSTPCNGVEGFNCYFPLQSYGFQPT | 3.47 | 21.37 |
| J | 520–540 | APATVCGPKKSTNLVKNKCVN | 2.86 | 20.76 |
| K | 660–680 | YECDIPIGAGICASYQTQTNS | 2.92 | 20.82 |
| L | 660–710 | YECDIPIGAGICASYQTQTNSPRRARSVASQSIIAYTMSLGAENSVAYSNN | 6.12 | 24.02 |
| M | 990–1035 | EVQIDRLITGRLQSLQTYVTQQLIRAAEIRASANLAATKMSECVLG | 5.75 | 23.65 |
| N | 931–970 | IGKIQDSLSSTASALGKLQDVVNQNAQALNTLVKQLSSNF | 4.18 | 22.08 |
| S21P2 | 709–727 | PSKPSKRSFIEDLLFNKV | 2.8 | 20.7 |
| S14P5 | 552–570 | TESNKKFLPFQQFGRDIA | 2.8 | 20.72 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Phiri, K.; Grill, L. Development of a Candidate TMV Epitope Display Vaccine against SARS-CoV-2. Vaccines 2024, 12, 448. https://doi.org/10.3390/vaccines12050448
Phiri K, Grill L. Development of a Candidate TMV Epitope Display Vaccine against SARS-CoV-2. Vaccines. 2024; 12(5):448. https://doi.org/10.3390/vaccines12050448
Chicago/Turabian StylePhiri, Kelvin, and Larry Grill. 2024. "Development of a Candidate TMV Epitope Display Vaccine against SARS-CoV-2" Vaccines 12, no. 5: 448. https://doi.org/10.3390/vaccines12050448
APA StylePhiri, K., & Grill, L. (2024). Development of a Candidate TMV Epitope Display Vaccine against SARS-CoV-2. Vaccines, 12(5), 448. https://doi.org/10.3390/vaccines12050448