Anti-Leishmanial Vaccines: Assumptions, Approaches, and Annulments

 , , ,
, , ,
Abstract
:1. Introduction
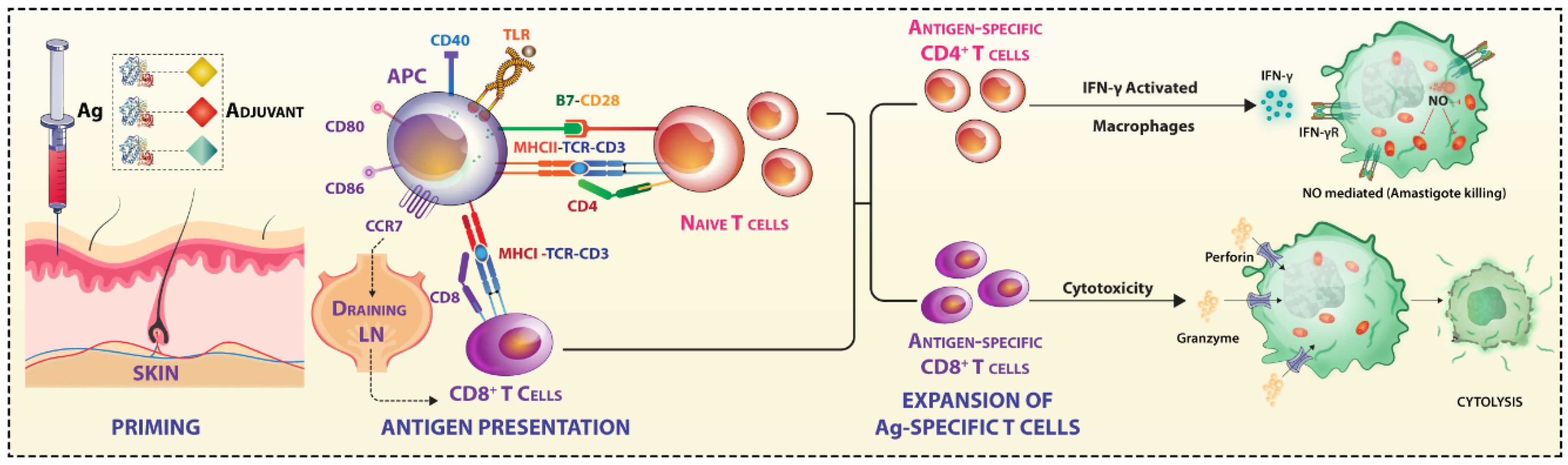
2. Factors in Antigenic Priming That Affect Vaccination Efficacy
2.1. Selection of an Infective-Stage Specific Vaccine Candidate
2.2. Determinant Selection as a Function of Relative Abundance of Leishmanial Proteins
2.3. Intracellular Compartmentalization Contributes to the Processing of an Antigen
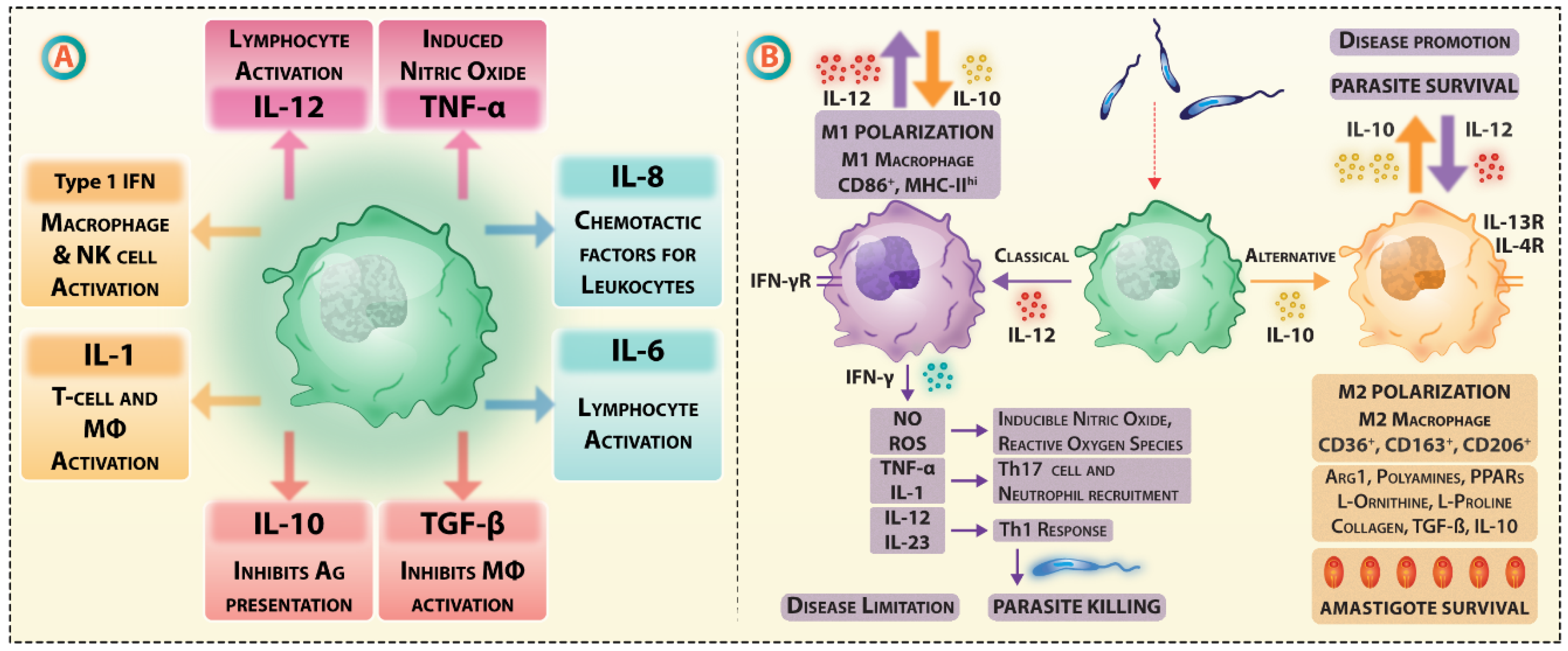
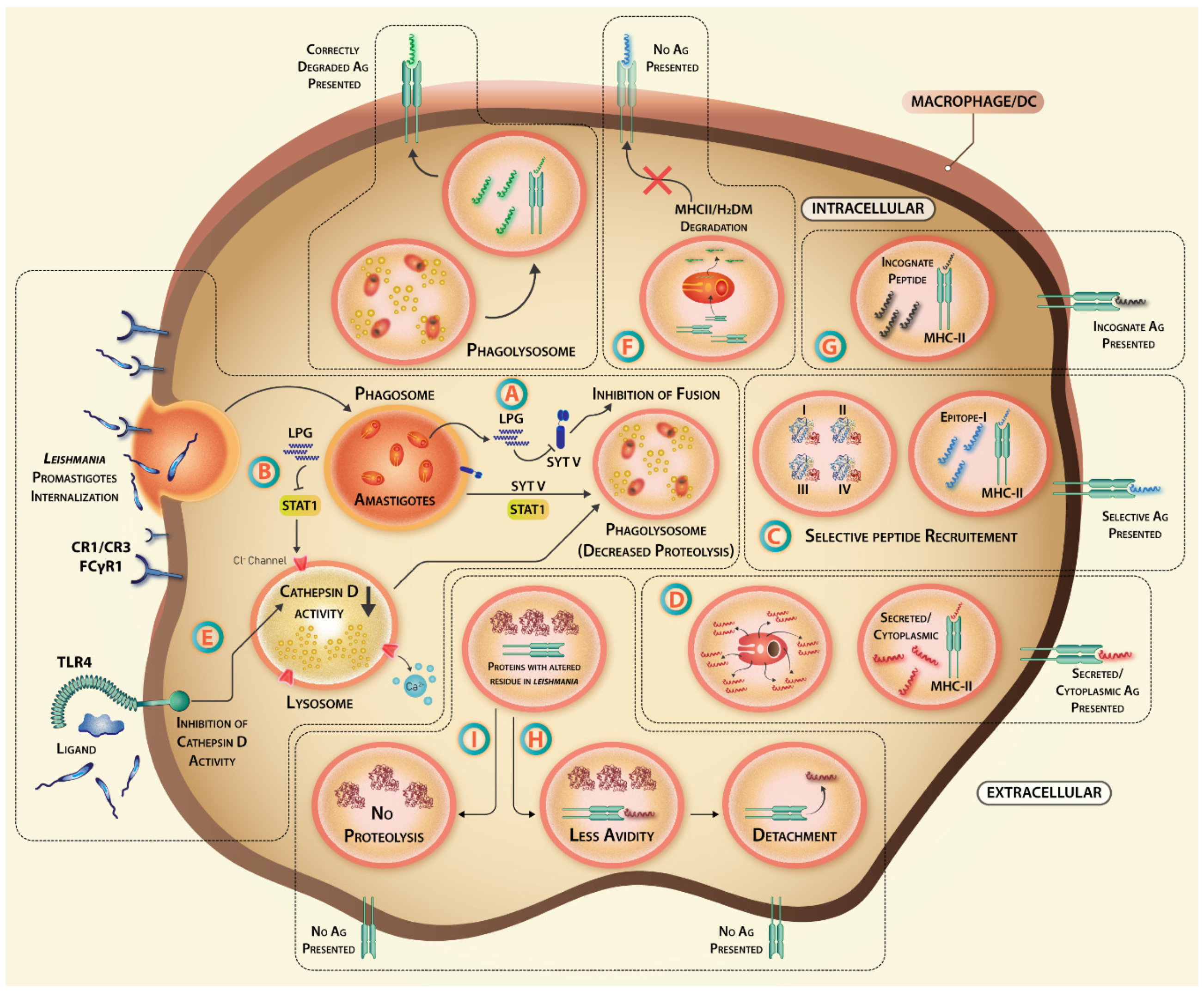
3. Leishmania Targeting of Antigen Processing and Presentation Affects Vaccination Efficacy
3.1. Receptor-Mediated Internalization of Leishmania Parasites
3.2. The Hijacking of Lysosomal Fusion Machinery inside Macrophages
3.3. Altered Activity of Proteases
3.4. Degradation of Antigen Presentation Machinery
3.5. Immunodominance and Epitope Crypticity: Importance in Anti-Leishmanial Vaccination
3.6. Loading of Antigen to MHC Molecule
3.7. Peptide-MHC-II Affinity: A Major Determinant of Immunodominant Epitope Selection
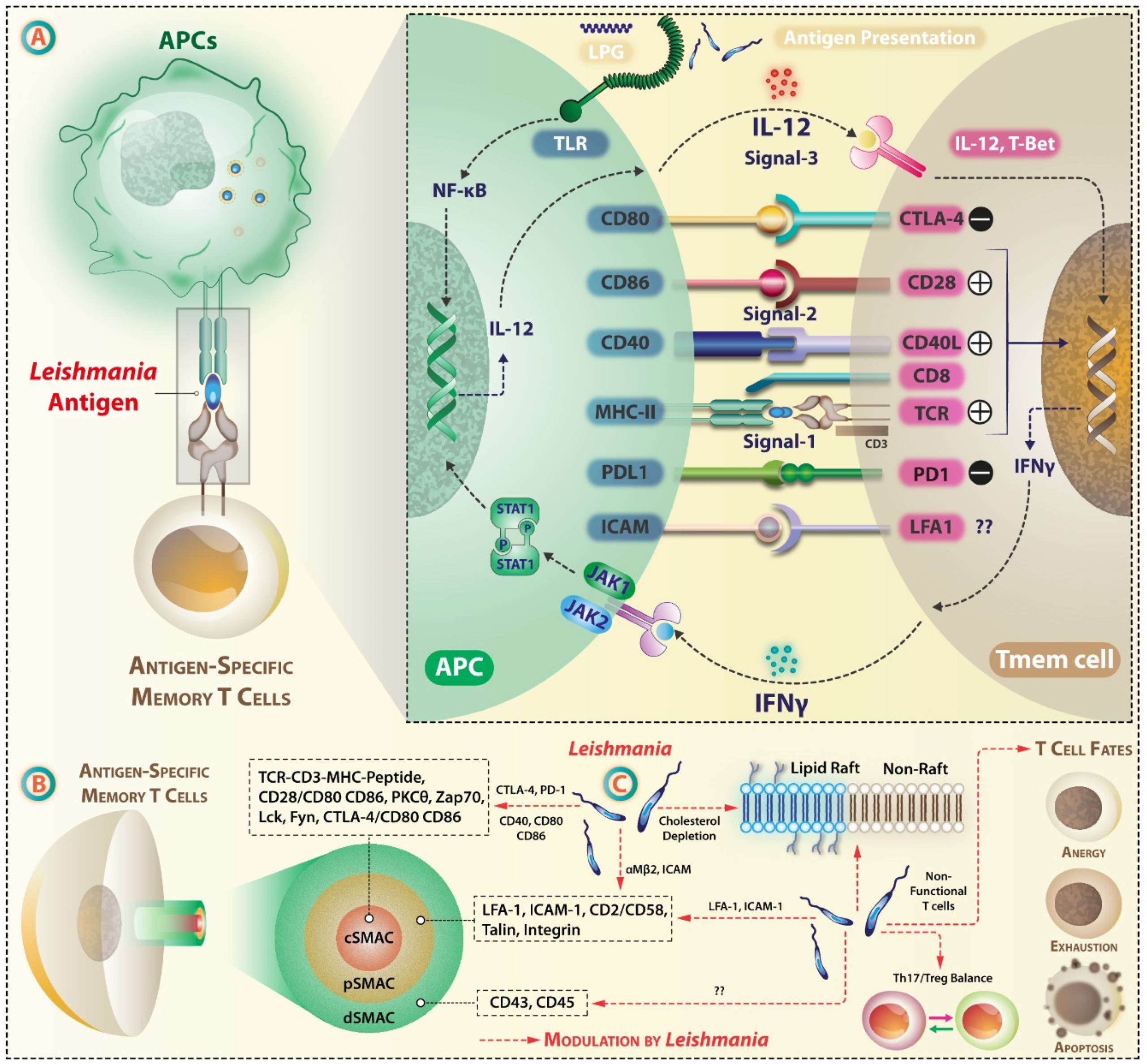
4. Leishmania Targets Antigen Presentation by APCs and Immune Synapse (IS)
5. Various Fates of T Cells during Infection
Plasticity of T Cells
6. Anergy, Exhaustion, and Apoptosis
Leishmania-Infected Macrophages Respond Differently to Antigen-Specific T Cells
7. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Garçon, N.; Stern, P.L.; Cunningham, A.L. Understanding Modern Vaccines: Perspectives in Vaccinology; Elsevier: Amsterdam, The Netherlands, 2011. [Google Scholar]
- Cambronero, M.R.; Prado-Cohrs, D.; Sanroma, M.L. Conceptos Inmunológicos Básicos Aplicados a La Vacunología. Vacunas 2017, 18, 49–58. [Google Scholar] [CrossRef]
- Esparza, J. A New Scientific Paradigm May Be Needed to Finally Develop an HIV Vaccine. Front. Immunol. 2015, 6, 124. [Google Scholar] [CrossRef] [PubMed]
- Cobey, S. Pathogen Evolution and the Immunological Niche. Ann. N. Y. Acad. Sci. 2014, 1320, 1. [Google Scholar] [CrossRef] [PubMed]
- Coler, R.N.; Reed, S.G. Second-Generation Vaccines against Leishmaniasis. Trends Parasitol. 2005, 21, 244–249. [Google Scholar] [CrossRef]
- Fernandes, A.P.; Coelho, E.A.F.; Machado-Coelho, G.L.L.; Junior, G.G.; Gazzinelli, R.T. Making an Anti-Amastigote Vaccine for Visceral Leishmaniasis: Rational, Update and Perspectives. Curr. Opin. Microbiol. 2012, 15, 476–485. [Google Scholar] [CrossRef]
- Kedzierski, L. Leishmaniasis Vaccine: Where Are We Today? J. Glob. Infect. Dis. 2010, 2, 177–185. [Google Scholar] [CrossRef]
- Burza, S.; Croft, S.L.; Boelaert, M. Leishmaniasis. Lancet 2018, 392, 951–970. [Google Scholar] [CrossRef]
- Titus, R.G. The Immunomodulatory Factors of Bloodfeeding Arthropod Saliva. Parasite Immunol. 2000, 22, 319–331. [Google Scholar]
- Zahedifard, F.; Gholami, E.; Taheri, T.; Taslimi, Y.; Doustdari, F.; Seyed, N.; Torkashvand, F.; Meneses, C.; Papadopoulou, B.; Kamhawi, S. Enhanced Protective Efficacy of Nonpathogenic Recombinant Leishmania Tarentolae Expressing Cysteine Proteinases Combined with a Sand Fly Salivary Antigen. PLoS Negl. Trop. Dis. 2014, 8, e2751. [Google Scholar] [CrossRef]
- Sacks, D.; Noben-Trauth, N. The Immunology of Susceptibility and Resistance to Leishmania Major in Mice. Nat. Rev. Immunol. 2002, 2, 845–858. [Google Scholar] [CrossRef]
- Zhang, W.-W.; Charest, H.; Ghedin, E.; Matlashewski, G. Identification and Overexpression of the A2 Amastigote-Specific Protein in Leishmania Donovani. Mol. Biochem. Parasitol. 1996, 78, 79–90. [Google Scholar] [CrossRef]
- Flinn, H.M.; Rangarajan, D.; Smith, D.F. Expression of a Hydrophilic Surface Protein in Infective Stages of Leishmania Major. Mol. Biochem. Parasitol. 1994, 65, 259–270. [Google Scholar] [CrossRef]
- Alce, T.M.; Gokool, S.; McGhie, D.; Stäger, S.; Smith, D.F. Expression of Hydrophilic Surface Proteins in Infective Stages of Leishmania Donovani. Mol. Biochem. Parasitol. 1999, 1, 191–196. [Google Scholar] [CrossRef]
- Dey, R.; Dagur, P.K.; Selvapandiyan, A.; McCoy, J.P.; Salotra, P.; Duncan, R.; Nakhasi, H.L. Live Attenuated Leishmania Donovani P27 Gene Knockout Parasites Are Nonpathogenic and Elicit Long-Term Protective Immunity in BALB/c Mice. J. Immunol. 2013, 190, 2138–2149. [Google Scholar] [CrossRef]
- Selvapandiyan, A.; Debrabant, A.; Duncan, R.; Muller, J.; Salotra, P.; Sreenivas, G.; Salisbury, J.L.; Nakhasi, H.L. Centrin Gene Disruption Impairs Stage-Specific Basal Body Duplication and Cell Cycle Progression in Leishmania. J. Biol. Chem. 2004, 279, 25703–25710. [Google Scholar] [CrossRef]
- Saljoughian, N.; Taheri, T.; Rafati, S. Live Vaccination Tactics: Possible Approaches for Controlling Visceral Leishmaniasis. Front. Immunol. 2014, 5, 134. [Google Scholar] [CrossRef] [Green Version]
- Bruhn, K.W.; Birnbaum, R.; Haskell, J.; Vanchinathan, V.; Greger, S.; Narayan, R.; Chang, P.L.; Tran, T.A.; Hickerson, S.M.; Beverley, S.M.; et al. Killed but Metabolically Active Leishmania Infantum as a Novel Whole-Cell Vaccine for Visceral Leishmaniasis. Clin. Vaccine Immunol. 2012, 19, 490–498. [Google Scholar] [CrossRef]
- Musa, A.M.; Khalil, E.A.G.; Mahgoub, F.A.E.; Elgawi, S.H.H.; Modabber, F.; Elkadaru, A.E.M.Y.; Aboud, M.H.; Noazin, S.; Ghalib, H.W.; El-Hassan, A.M. Immunochemotherapy of Persistent Post-Kala-Azar Dermal Leishmaniasis: A Novel Approach to Treatment. Trans. R. Soc. Trop. Med. Hyg. 2008, 102, 58–63. [Google Scholar] [CrossRef]
- Palatnik-de-Sousa, C.B.; Moreno, M.; Paraguai-de-Souza, E.; Borojevic, R. The FML Vaccine (Fucose-Manose Ligand) Protects Hamsters from Experimental Kala-Azar. Cienc. Cult. 1994, 64, 290–296. [Google Scholar]
- Santos, W.R.; Paraguai de Souza, E.; Palatnik, M.; Palatnik de Sousa, C.B.; de Souza, E.P.; Palatnik, M.; de Sousa, C.B.P. Vaccination of Swiss Albino Mice against Experimental Visceral Leishmaniasis with the FML Antigen of Leishmaniadonovani. Vaccine 1999, 17, 2554–2561. [Google Scholar] [CrossRef]
- Nogueira, F.S.; Moreira, M.A.B.; Borja-Cabrera, G.P.; Santos, F.N.; Menz, I.; Parra, L.E.; Xu, Z.; Chu, H.J.; Palatnik-de-Sousa, C.B.; Luvizotto, M.C.R. Leishmune® Vaccine Blocks the Transmission of Canine Visceral Leishmaniasis: Absence of Leishmania Parasites in Blood, Skin and Lymph Nodes of Vaccinated Exposed Dogs. Vaccine 2005, 23, 4805–4810. [Google Scholar] [CrossRef] [PubMed]
- Saraiva, E.M.; Barbosa, A.D.F.; Santos, F.N.; Borja-Cabrera, G.P.; Nico, D.; Souza, L.O.P.; Mendes-Aguiar, C.D.O.; De Souza, E.P.; Fampa, P.; Parra, L.E.; et al. The FML-Vaccine (Leishmune®) against Canine Visceral Leishmaniasis: A Transmission Blocking Vaccine. Vaccine 2006, 24, 2423–2431. [Google Scholar] [CrossRef] [PubMed]
- Afrin, F.; Ali, N. Adjuvanticity and Protective Immunity Elicited by Leishmania Donovani Antigens Encapsulated in Positively Charged Liposomes. Infect. Immun. 1997, 65, 2371–2377. [Google Scholar]
- Bhowmick, S.; Ravindran, R.; Ali, N. Leishmanial Antigens in Liposomes Promote Protective Immunity and Provide Immunotherapy against Visceral Leishmaniasis via Polarized Th1 Response. Vaccine 2007, 25, 6544–6556. [Google Scholar] [CrossRef] [PubMed]
- Mazumder, S.; Ravindran, R.; Banerjee, A.; Ali, N. Non-Coding PDNA Bearing Immunostimulatory Sequences Co-Entrapped with Leishmanial Antigens in Cationic Liposomes Elicits Almost Complete Protection against Experimental Visceral Leishmaniasis in BALB/c Mice. Vaccine 2007, 25, 8771–8781. [Google Scholar] [CrossRef] [PubMed]
- Ravindran, R.; Maji, M.; Ali, N. Vaccination with Liposomal Leishmanial Antigens Adjuvanted with Monophosphoryl Lipid-Trehalose Dicorynomycolate (MPL-TDM) Confers Long-Term Protection against Visceral Leishmaniasis through a Human Administrable Route. Mol. Pharm. 2012, 9, 59–70. [Google Scholar] [CrossRef] [PubMed]
- De Souza, E.P.; Bernardo, R.R.; Palatnik, M.; de Sousa, C.B.P. Vaccination of Balb/c Mice against Experimental Visceral Leishmaniasis with the GP36 Glycoprotein Antigen of Leishmania Donovani. Vaccine 2001, 19, 3104–3115. [Google Scholar] [CrossRef]
- Bhowmick, S.; Ali, N. Identification of Novel Leishmania Donovani Antigens That Help Define Correlates of Vaccine-Mediated Protection in Visceral Leishmaniasis. PLoS ONE 2009, 4, e5820. [Google Scholar] [CrossRef] [PubMed]
- Jaffe, C.L.; Rachamim, N.; Sarfstein, R. Characterization of Two Proteins from Leishmania Donovani and Their Use for Vaccination against Visceral Leishmaniasis. J. Immunol. 1990, 144, 699–706. [Google Scholar] [PubMed]
- Tewary, P.; Mehta, J.; Sukumaran, B.; Madhubala, R. Vaccination with Leishmania Soluble Antigen and Immunostimulatory Oligodeoxynucleotides Induces Specific Immunity and Protection against Leishmania Donovani Infection. FEMS Immunol. Med. Microbiol. 2004, 42, 241–248. [Google Scholar] [CrossRef]
- Iborra, S.; Carrión, J.; Anderson, C.; Alonso, C.; Sacks, D.; Soto, M. Vaccination with the Leishmania Infantum Acidic Ribosomal P0 Protein plus CpG Oligodeoxynucleotides Induces Protection against Cutaneous Leishmaniasis in C57BL/6 Mice but Does Not Prevent Progressive Disease in BALB/c Mice. Infect. Immun. 2005, 73, 5842–5852. [Google Scholar] [CrossRef] [PubMed]
- Garg, R.; Dube, A. Animal Models for Vaccine Studies for Visceral Leishmaniasis. Indian J. Med. Res. 2006, 123, 439–454. [Google Scholar] [PubMed]
- Kumari, S.; Samant, M.; Misra, P.; Khare, P.; Sisodia, B.; Shasany, A.K.; Dube, A. Th1-Stimulatory Polyproteins of Soluble Leishmania Donovani Promastigotes Ranging from 89.9 to 97.1 KDa Offers Long-Lasting Protection against Experimental Visceral Leishmaniasis. Vaccine 2008, 26, 5700–5711. [Google Scholar] [CrossRef] [PubMed]
- Kumari, S.; Samant, M.; Khare, P.; Sundar, S.; Sinha, S.; Dube, A. Induction of Th1-Type Cellular Responses in Cured/Exposed Leishmania-Infected Patients and Hamsters against Polyproteins of Soluble Leishmania Donovani Promastigotes Ranging from 89.9 to 97.1 KDa. Vaccine 2008, 26, 4813–4818. [Google Scholar] [CrossRef]
- Karanja, R.; Ingonga, J.; Mwangi, M.; Mwala, D.; Lugalia, R.; Magambo, J.; Tonui, W. Immunization with a Combination of Leishmania Major Lipophosphoglycan (LPG) and Phlebotomus Duboscqi Salivary Gland Lysates (SGLs) Abrogates Protective Effect of LPG against L. Major in BALB/c Mice. Afr. J. Health Sci. 2011, 18, 1–5. [Google Scholar]
- Tonui, W.K.; Mbati, P.A.; Anjili, C.O.; Orago, A.S.; Turco, S.J.; Githure, J.I.; Keoch, D.K. Transmission Blocking Vaccine Studies in Leishmaniasis: II Effect of Immunisation Using Leshmania Major Derived 63 Kilodalton on the Course of L. Major Infection in BALB/c Mice. East Afr. Med. J. 2001, 78, 90–92. [Google Scholar] [CrossRef]
- Bhowmick, S.; Ravindran, R.; Ali, N. Gp63 in Stable Cationic Liposomes Confers Sustained Vaccine Immunity to Susceptible BALB/c Mice Infected with Leishmania Donovani. Infect. Immun. 2008, 76, 1003–1015. [Google Scholar] [CrossRef]
- Handman, E.; Button, L.L.; McMaster, R.W. Leishmania Major: Production of Recombinant Gp63, Its Antigenicity and Immunogenicity in Mice. Exp. Parasitol. 1990, 70, 427–435. [Google Scholar] [CrossRef]
- Abdelhak, S.; Louzir, H.; Timm, J.; Blel, L.; Benlasfar, Z.; Lagranderie, M.; Gheorghiu, M.; Dellagi, K.; Gicquel, B. Recombinant BCG Expressing the Leishmania Surface Antigen Gp63 Induces Protective Immunity against Leishmania Major Infection in BALB/c Mice. Microbiology 1995, 141, 1585–1592. [Google Scholar] [CrossRef]
- Badiee, A.; Jaafari, M.R.; Khamesipour, A.; Samiei, A.; Soroush, D.; Kheiri, M.T.; Barkhordari, F.; McMaster, W.R.; Mahboudi, F. Enhancement of Immune Response and Protection in BALB/c Mice Immunized with Liposomal Recombinant Major Surface Glycoprotein of Leishmania (Rgp63): The Role of Bilayer Composition. Colloids Surf. B Biointerfaces 2009, 74, 37–44. [Google Scholar] [CrossRef]
- Mazumder, S.; Maji, M.; Das, A.; Ali, N. Potency, Efficacy and Durability of DNA/DNA, DNA/Protein and Protein/Protein Based Vaccination Using Gp63 against Leishmania Donovani in BALB/c Mice. PLoS ONE 2011, 6, e14644. [Google Scholar] [CrossRef] [PubMed]
- Chenik, M.; Louzir, H.; Ksontini, H.; Dilou, A.; Abdmouleh, I.; Dellagi, K. Vaccination with the Divergent Portion of the Protein Histone H2B of Leishmania Protects Susceptible BALB/c Mice against a Virulent Challenge with Leishmania Major. Vaccine 2006, 24, 2521–2529. [Google Scholar] [CrossRef] [PubMed]
- Meddeb-Garnaoui, A.; Toumi, A.; Ghelis, H.; Mahjoub, M.; Louzir, H.; Chenik, M. Cellular and Humoral Responses Induced by Leishmania Histone H2B and Its Divergent and Conserved Parts in Cutaneous and Visceral Leishmaniasis Patients, Respectively. Vaccine 2010, 28, 1881–1886. [Google Scholar] [CrossRef] [PubMed]
- Dole, V.S.; Raj, V.S.; Ghosh, A.; Madhubala, R.; Myler, P.J.; Stuart, K.D. Immunization with Recombinant LD1 Antigens Protects against Experimental Leishmaniasis. Vaccine 2000, 19, 423–430. [Google Scholar] [CrossRef]
- Tewary, P.; Sukumaran, B.; Saxena, S.; Madhubala, R. Immunostimulatory Oligodeoxynucleotides Are Potent Enhancers of Protective Immunity in Mice Immunized with Recombinant ORFF Leishmanial Antigen. Vaccine 2004, 22, 3053–3060. [Google Scholar] [CrossRef]
- Sharma, A.; Madhubala, R. Ubiquitin Conjugation of Open Reading Frame F DNA Vaccine Leads to Enhanced Cell-Mediated Immune Response and Induces Protection against Both Antimony-Susceptible and-Resistant Strains of Leishmania Donovani. J. Immunol. 2009, 183, 7719–7731. [Google Scholar] [CrossRef]
- Kushawaha, P.K.; Gupta, R.; Sundar, S.; Sahasrabuddhe, A.A.; Dube, A. Elongation Factor-2, a Th1 Stimulatory Protein of Leishmania Donovani, Generates Strong IFN-γ and IL-12 Response in Cured Leishmania -Infected Patients/Hamsters and Protects Hamsters against Leishmania Challenge. J. Immunol. 2011, 187, 6417–6427. [Google Scholar] [CrossRef]
- Bhardwaj, S.; Vasishta, R.K.; Arora, S.K. Vaccination with a Novel Recombinant Leishmania Antigen plus MPL Provides Partial Protection against L. Donovani Challenge in Experimental Model of Visceral Leishmaniasis. Exp. Parasitol. 2009, 121, 29–37. [Google Scholar] [CrossRef]
- Stäger, S.; Smith, D.F.; Kaye, P.M. Immunization with a Recombinant Stage-Regulated Surface Protein from Leishmania Donovani Induces Protection against Visceral Leishmaniasis. J. Immunol. 2000, 165, 7064–7071. [Google Scholar] [CrossRef]
- Nagill, R.; Kaur, S. Enhanced Efficacy and Immunogenicity of 78 KDa Antigen Formulated in Various Adjuvants against Murine Visceral Leishmaniasis. Vaccine 2010, 28, 4002–4012. [Google Scholar] [CrossRef]
- Streit, J.A.; Recker, T.J.; Donelson, J.E.; Wilson, M.E. BCG Expressing LCR1 of Leishmania Chagasi Induces Protective Immunity in Susceptible Mice. Exp. Parasitol. 2000, 94, 33–41. [Google Scholar] [CrossRef] [PubMed]
- Serezani, C.H.C.; Franco, A.R.; Wajc, M.; Yokoyama-Yasunaka, J.K.U.; Wunderlich, G.; Borges, M.M.; Uliana, S.R.B. Evaluation of the Murine Immune Response to Leishmania Meta 1 Antigen Delivered as Recombinant Protein or DNA Vaccine. Vaccine 2002, 20, 3755–3763. [Google Scholar] [CrossRef]
- Gomez-Rodriguez, J.; Wohlfert, E.A.; Handon, R.; Meylan, F.; Wu, J.Z.; Anderson, S.M.; Kirby, M.R.; Belkaid, Y.; Schwartzberg, P.L. Itk-Mediated Integration of T Cell Receptor and Cytokine Signaling Regulates the Balance between Th17 and Regulatory T Cells. J. Exp. Med. 2014, 211, 529–543. [Google Scholar] [CrossRef] [PubMed]
- Morris, R.V.; Shoemaker, C.B.; David, J.R.; Lanzaro, G.C.; Titus, R.G. Sandfly Maxadilan Exacerbates Infection with Leishmania Major and Vaccinating against It Protects against L. Major Infection. J. Immunol. 2001, 167, 5226–5230. [Google Scholar] [CrossRef] [PubMed]
- Gomes, R.; Teixeira, C.; Teixeira, M.J.; Oliveira, F.; Menezes, M.J.; Silva, C.; De Oliveira, C.I.; Miranda, J.C.; Elnaiem, D.E.; Kamhawi, S.; et al. Immunity to a Salivary Protein of a Sand Fly Vector Protects against the Fatal Outcome of Visceral Leishmaniasis in a Hamster Model. Proc. Natl. Acad. Sci. USA 2008, 105, 7845–7850. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, A.; Zhang, W.W.; Matlashewski, G. Immunization with A2 Protein Results in a Mixed Th1/Th2 and a Humoral Response Which Protects Mice against Leishmania Donovani Infections. Vaccine 2001, 20, 59–66. [Google Scholar] [CrossRef]
- Henriquez, F.L.; Campbell, S.A.; Roberts, C.W.; Mullen, A.B.; Burchmore, R.; Carter, K.C. Vaccination with Recombinant Leishmania Donovani Gamma-Glutamylcysteine Synthetase Fusion Protein Protects Against L. Donovani Infection. J. Parasitol. 2010, 96, 929–936. [Google Scholar] [CrossRef]
- Carcelen, J.; Iniesta, V.; Fernandez-Cotrina, J.; Serrano, F.; Parejo, J.C.; Corraliza, I.; Gallardo-Soler, A.; Marañón, F.; Soto, M.; Alonso, C.; et al. The Chimerical Multi-Component Q Protein from Leishmania in the Absence of Adjuvant Protects Dogs against an Experimental Leishmania Infantum Infection. Vaccine 2009, 27, 5964–5973. [Google Scholar] [CrossRef]
- Zadeh-Vakili, A.; Taheri, T.; Taslimi, Y.; Doustdari, F.; Salmanian, A.H.; Rafati, S. Immunization with the Hybrid Protein Vaccine, Consisting of Leishmania Major Cysteine Proteinases Type I (CPB) and Type II (CPA), Partially Protects against Leishmaniasis. Vaccine 2004, 22, 1930–1940. [Google Scholar] [CrossRef]
- Kronenberg, K.; Brosch, S.; Butsch, F.; Tada, Y.; Shibagaki, N.; Udey, M.C.; Von Stebut, E. Vaccination with TAT-Antigen Fusion Protein Induces Protective, CD8+ T Cell-Mediated Immunity against Leishmania Major. J. Investig. Dermatol. 2010, 130, 2602–2610. [Google Scholar] [CrossRef]
- Skeiky, Y.A.W.; Coler, R.N.; Brannon, M.; Stromberg, E.; Greeson, K.; Crane, R.T.; Campos-Neto, A.; Reed, S.G. Protective Efficacy of a Tandemly Linked, Multi-Subunit Recombinant Leishmanial Vaccine (Leish-111f) Formulated in MPL® Adjuvant. Vaccine 2002, 20, 3292–3303. [Google Scholar] [CrossRef]
- Coler, R.N.; Goto, Y.; Bogatzki, L.; Raman, V.; Reed, S.G. Leish-111f, a Recombinant Polyprotein Vaccine That Protects against Visceral Leishmaniasis by Elicitation of CD4+ T Cells. Infect. Immun. 2007, 75, 4648–4654. [Google Scholar] [CrossRef] [PubMed]
- Chakravarty, J.; Kumar, S.; Trivedi, S.; Rai, V.K.; Singh, A.; Ashman, J.A.; Laughlin, E.M.; Coler, R.N.; Kahn, S.J.; Beckmann, A.M.; et al. A Clinical Trial to Evaluate the Safety and Immunogenicity of the LEISH-F1+MPL-SE Vaccine for Use in the Prevention of Visceral Leishmaniasis. Vaccine 2011, 29, 3531–3537. [Google Scholar] [CrossRef] [PubMed]
- Titus, R.G.; Gueiros-Filho, F.J.; De Freitas, L.A.R.; Beverley, S.M. Development of a Safe Live Leishmania Vaccine Line by Gene Replacement. Proc. Natl. Acad. Sci. USA 1995, 92, 10267–10271. [Google Scholar] [CrossRef] [PubMed]
- Veras, P.S.T.; Brodskyn, C.I.; Balestieri, F.M.P.; De Freitas, L.A.R.; Ramos, A.P.S.; Queiroz, A.R.P.; Barral, A.; Beverley, S.M.; Barral-Netto, M. A Dhfr-Ts- Leishmania Major Knockout Mutant Cross-Protects against Leishmania Amazonensis. Mem. Inst. Oswaldo Cruz 1999, 94, 491–496. [Google Scholar] [CrossRef] [PubMed]
- Selvapandiyan, A.; Dey, R.; Nylen, S.; Duncan, R.; Sacks, D.; Nakhasi, H.L. Intracellular Replication-Deficient Leishmania Donovani Induces Long Lasting Protective Immunity against Visceral Leishmaniasis. J. Immunol. 2009, 183, 1813–1820. [Google Scholar] [CrossRef]
- Papadopoulou, B.; Roy, G.; Breton, M.; Kündig, C.; Dumas, C.; Fillion, I.; Singh, A.K.; Olivier, M.; Ouellette, M. Reduced Infectivity of a Leishmania Donovani Biopterin Transporter Genetic Mutant and Its Use as an Attenuated Strain for Vaccination. Infect. Immun. 2002, 70, 62–68. [Google Scholar] [CrossRef]
- Silvestre, R.; Cordeiro-Da-Silva, A.; Santarém, N.; Vergnes, B.; Sereno, D.; Ouaissi, A. SIR2-Deficient Leishmania Infantum Induces a Defined IFN-γ/IL-10 Pattern That Correlates with Protection. J. Immunol. 2007, 179, 3161–3170. [Google Scholar] [CrossRef]
- Dey, R.; Meneses, C.; Salotra, P.; Kamhawi, S.; Nakhasi, H.L.; Duncan, R. Characterization of a Leishmania Stage-Specific Mitochondrial Membrane Protein That Enhances the Activity of Cytochrome c Oxidase and Its Role in Virulence. Mol. Microbiol. 2010, 77, 399–414. [Google Scholar] [CrossRef]
- Carrión, J.; Folgueira, C.; Soto, M.; Fresno, M.; Requena, J.M. Leishmania Infantum HSP70-II Null Mutant as Candidate Vaccine against Leishmaniasis: A Preliminary Evaluation. Parasites Vectors 2011, 4, 150. [Google Scholar] [CrossRef]
- Solana, J.C.; Ramírez, L.; Corvo, L.; de Oliveira, C.I.; Barral-Netto, M.; Requena, J.M.; Iborra, S.; Soto, M. Vaccination with a Leishmania Infantum HSP70-II Null Mutant Confers Long-Term Protective Immunity against Leishmania Major Infection in Two Mice Models. PLoS Negl. Trop. Dis. 2017, 11, e0005644. [Google Scholar] [CrossRef] [PubMed]
- Kébaïer, C.; Uzonna, J.E.; Beverley, S.M.; Scott, P. Immunization with Persistent Attenuated Δlpg2 Leishmania Major Parasites Requires Adjuvant to Provide Protective Immunity in C57BL/6 Mice. Infect. Immun. 2006, 74, 777–780. [Google Scholar] [CrossRef] [PubMed]
- Uzonna, J.E.; Späth, G.F.; Beverley, S.M.; Scott, P. Vaccination with Phosphoglycan-Deficient Leishmania Major Protects Highly Susceptible Mice from Virulent Challenge without Inducing a Strong Th1 Response. J. Immunol. 2004, 172, 3793–3797. [Google Scholar] [CrossRef] [PubMed]
- Breton, M.; Tremblay, M.J.; Ouellette, M.; Papadopoulou, B. Live Nonpathogenic Parasitic Vector as a Candidate Vaccine against Visceral Leishmaniasis. Infect. Immun. 2005, 73, 6372–6382. [Google Scholar] [CrossRef] [Green Version]
- Mizbani, A.; Taheri, T.; Zahedifard, F.; Taslimi, Y.; Azizi, H.; Azadmanesh, K.; Papadopoulou, B.; Rafati, S. Recombinant Leishmania Tarentolae Expressing the A2 Virulence Gene as a Novel Candidate Vaccine against Visceral Leishmaniasis. Vaccine 2009, 28, 53–62. [Google Scholar] [CrossRef]
- Yam, K.K.; Hugentobler, F.; Pouliot, P.; Stern, A.M.; Lalande, J.D.; Matlashewski, G.; Olivier, M.; Cousineau, B. Generation and Evaluation of A2-Expressing Lactococcus Lactis Live Vaccines against Leishmania Donovani in BALB/c Mice. J. Med. Microbiol. 2011, 60, 1248–1260. [Google Scholar] [CrossRef]
- Gurunathan, S.; Sacks, D.L.; Brown, D.R.; Reiner, S.L.; Charest, H.; Glaichenhaus, N.; Seder, R.A. Vaccination with DNA Encoding the Immunodominant LACK Parasite Antigen Confers Protective Immunity to Mice Infected with Leishmania Major. J. Exp. Med. 1997, 186, 1137–1147. [Google Scholar] [CrossRef]
- Gomes, D.C.d.O.; Pinto, E.F.; de Melo, L.D.B.; Lima, W.P.; Larraga, V.; Lopes, U.G.; Rossi-Bergmann, B.; de Oliveira Gomes, D.C.; Pinto, E.F.; de Melo, L.D.B.; et al. Intranasal Delivery of Naked DNA Encoding the LACK Antigen Leads to Protective Immunity against Visceral Leishmaniasis in Mice. Vaccine 2007, 25, 2168–2172. [Google Scholar] [CrossRef]
- De, D.O.G.; Da, B.S.C.S.; De, H.M.G.; Lopes, U.G.; Rossi-Bergmann, B.; De Oliveira Gomes, D.C.; Da Silva Costa Souza, B.L.; De Matos Guedes, H.L.; Lopes, U.G.; Rossi-Bergmann, B. Intranasal Immunization with LACK-DNA Promotes Protective Immunity in Hamsters Challenged with Leishmania Chagasi. Parasitology 2011, 138, 1892–1897. [Google Scholar] [CrossRef]
- Melby, P.C.; Yang, J.; Zhao, W.; Perez, L.E.; Cheng, J. Leishmania Donovani P36(LACK) DNA Vaccine Is Highly Immunogenic but Not Protective against Experimental Visceral Leishmaniasis. Infect. Immun. 2001, 69, 4719–4725. [Google Scholar] [CrossRef]
- Kaur, T.; Sobti, R.C.; Kaur, S. Cocktail of Gp63 and Hsp70 Induces Protection against Leishmania Donovani in BALB/c Mice. Parasite Immunol. 2011, 33, 95–103. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, A.; Labrecque, S.; Matlashewski, G. Protection against Leishmania Donovani Infection by DNA Vaccination: Increased DNA Vaccination Efficiency through Inhibiting the Cellular P53 Response. Vaccine 2001, 19, 3169–3178. [Google Scholar] [CrossRef]
- Resende, D.M.; Caetano, B.C.; Dutra, M.S.; Penido, M.L.O.; Abrantes, C.F.; Verly, R.M.; Resende, J.M.; Piló-Veloso, D.; Rezende, S.A.; Bruna-Romero, O.; et al. Epitope Mapping and Protective Immunity Elicited by Adenovirus Expressing the Leishmania Amastigote Specific A2 Antigen: Correlation with IFN-γ and Cytolytic Activity by CD8+ T Cells. Vaccine 2008, 26, 4585–4593. [Google Scholar] [CrossRef] [PubMed]
- Sukumaran, B.; Tewary, P.; Saxena, S.; Madhubala, R. Vaccination with DNA Encoding ORFF Antigen Confers Protective Immunity in Mice Infected with Leishmania Donovani. Vaccine 2003, 21, 1292–1299. [Google Scholar] [CrossRef]
- Tewary, P.; Saxena, S.; Madhubala, R. Co-administration of IL-12 DNA with rORFF antigen confers long-term protective immunity against experimental visceral leishmaniaisis. Vaccine 2006, 24, 2409–2416. [Google Scholar] [CrossRef]
- Basu, R.; Bhaumik, S.; Basu, J.M.; Naskar, K.; De, T.; Roy, S. Kinetoplastid Membrane Protein-11 DNA Vaccination Induces Complete Protection against Both Pentavalent Antimonial-Sensitive and -Resistant Strains of Leishmania Donovani That Correlates with Inducible Nitric Oxide Synthase Activity and IL-4 Generation: E. J. Immunol. 2005, 174, 7160–7171. [Google Scholar] [CrossRef]
- Bhaumik, S.; Basu, R.; Sen, S.; Naskar, K.; Roy, S. KMP-11 DNA Immunization Significantly Protects against L. Donovani Infection but Requires Exogenous IL-12 as an Adjuvant for Comparable Protection against L. Major. Vaccine 2009, 27, 1306–1316. [Google Scholar] [CrossRef]
- Carter, K.C.; Henriquez, F.L.; Campbell, S.A.; Roberts, C.W.; Nok, A.; Mullen, A.B.; McFarlane, E. DNA Vaccination against the Parasite Enzyme Gamma-Glutamylcysteine Synthetase Confers Protection against Leishmania Donovani Infection. Vaccine 2007, 25, 4502–4509. [Google Scholar] [CrossRef]
- Saldarriaga, O.A.; Travi, B.L.; Park, W.; Perez, L.E.; Melby, P.C. Immunogenicity of a Multicomponent DNA Vaccine against Visceral Leishmaniasis in Dogs. Vaccine 2006, 24, 1928–1940. [Google Scholar] [CrossRef]
- Samant, M.; Gupta, R.; Kumari, S.; Misra, P.; Khare, P.; Kushawaha, P.K.; Sahasrabuddhe, A.A.; Dube, A. Immunization with the DNA-Encoding N-Terminal Domain of Proteophosphoglycan of Leishmania Donovani Generates Th1-Type Immunoprotective Response against Experimental Visceral Leishmaniasis. J. Immunol. 2009, 183, 470–479. [Google Scholar] [CrossRef]
- Sachdeva, R.; Banerjea, A.C.; Malla, N.; Dubey, M.L. Immunogenicity and Efficacy of Single Antigen Gp63, Polytope and PolytopeHSP70 DNA Vaccines against Visceral Leishmaniasis in Experimental Mouse Model. PLoS ONE 2009, 4, e7880. [Google Scholar] [CrossRef] [PubMed]
- Aguilar-Be, I.; da Silva Zardo, R.; de Souza, E.P.; Borja-Cabrera, G.P.; Rosado-Vallado, M.; Mut-Martin, M.; del Rosario García-Miss, M.; de Sousa, C.B.P.; Dumonteil, E. Cross-Protective Efficacy of a Prophylactic Leishmania Donovani DNA Vaccine against Visceral and Cutaneous Murine Leishmaniasis. Infect. Immun. 2005, 73, 812–819. [Google Scholar] [CrossRef] [PubMed]
- Gamboa-León, R.; Paraguai de Souza, E.; Borja-Cabrera, G.P.; Santos, F.N.; Myashiro, L.M.; Pinheiro, R.O.; Dumonteil, E.; Palatnik-de-Sousa, C.B. Immunotherapy against Visceral Leishmaniasis with the Nucleoside Hydrolase-DNA Vaccine of Leishmania Donovani. Vaccine 2006, 24, 4863–4873. [Google Scholar] [CrossRef] [PubMed]
- Das, S.; Freier, A.; Boussoffara, T.; Das, S.; Oswald, D.; Losch, F.O.; Selka, M.; Sacerdoti-Sierra, N.; Schönian, G.; Wiesmüller, K.H.; et al. Modular Multiantigen T Cell Epitope-Enriched DNA Vaccine against Human Leishmaniasis. Sci. Transl. Med. 2014, 6, 234ra56. [Google Scholar] [CrossRef]
- Rodríguez-Cortés, A.; Ojeda, A.; López-Fuertes, L.; Timón, M.; Altet, L.; Solano-Gallego, L.; Sánchez-Robert, E.; Francino, O.; Alberola, J. Vaccination with Plasmid DNA Encoding KMPII, TRYP, LACK and GP63 Does Not Protect Dogs against Leishmania Infantum Experimental Challenge. Vaccine 2007, 25, 7962–7971. [Google Scholar] [CrossRef]
- Fragaki, K.; Suffia, I.; Ferrua, B.; Rousseau, D.; Le Fichoux, Y.; Kubar, J. Immunisation with DNA Encoding Leishmania Infantum Protein PapLe22 Decreases the Frequency of Parasitemic Episodes in Infected Hamsters. Vaccine 2001, 19, 1701–1709. [Google Scholar] [CrossRef]
- Guha, R.; Gupta, D.; Rastogi, R.; Vikram, R.; Krishnamurthy, G.; Bimal, S.; Roy, S.; Mukhopadhyay, A. Vaccination with Leishmania Hemoglobin Receptor-Encoding DNA Protects against Visceral Leishmaniasis. Sci. Transl. Med. 2013, 5, 202ra121. [Google Scholar] [CrossRef]
- Lakshmi, B.S.; Wang, R.; Madhubala, R. Leishmania Genome Analysis and High-Throughput Immunological Screening Identifies Tuzin as a Novel Vaccine Candidate against Visceral Leishmaniasis. Vaccine 2014, 32, 3816–3822. [Google Scholar] [CrossRef]
- Tewary, P.; Jain, M.; Sahani, M.H.; Saxena, S.; Madhubala, R. A Heterologous Prime-Boost Vaccination Regimen Using ORFF DNA and Recombinant ORFF Protein Confers Protective Immunity against Experimental Visceral Leishmaniasis. J. Infect. Dis. 2005, 191, 2130–2137. [Google Scholar] [CrossRef]
- Rafati, S.; Zahedifard, F.; Nazgouee, F. Prime-Boost Vaccination Using Cysteine Proteinases Type I and II of Leishmania Infantum Confers Protective Immunity in Murine Visceral Leishmaniasis. Vaccine 2006, 24, 2169–2175. [Google Scholar] [CrossRef]
- Masih, S.; Arora, S.K.; Vasishta, R.K. Efficacy of Leishmania Donovani Ribosomal P1 Gene as DNA Vaccine in Experimental Visceral Leishmaniasis. Exp. Parasitol. 2011, 129, 55–64. [Google Scholar] [CrossRef] [PubMed]
- Saljoughian, N.; Taheri, T.; Zahedifard, F.; Taslimi, Y.; Doustdari, F.; Bolhassani, A.; Doroud, D.; Azizi, H.; Heidari, K.; Vasei, M.; et al. Development of Novel Prime-Boost Strategies Based on a Tri-Gene Fusion Recombinant L. tarentolae Vaccine against Experimental Murine Visceral Leishmaniasis. PLoS Negl. Trop. Dis. 2013, 7, e2174. [Google Scholar] [CrossRef] [PubMed]
- Dondji, B.; Pérez-Jimenez, E.; Goldsmith-Pestana, K.; Esteban, M.; McMahon-Pratt, D. Heterologous Prime-Boost Vaccination with the LACK Antigen Protects against Murine Visceral Leishmaniasis. Infect. Immun. 2005, 73, 5286–5289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dondji, B.; Deak, E.; Goldsmith-Pestana, K.; Pererz-Jimenez, E.; Esteban, M.; Miyake, S.; Yamamura, T.; McMahon-Pratt, D. Intradermal NKT Cell Activation during DNA Priming in Heterologous Prime-Boost Vaccination Enhances T Cell Responses and Protection against Leishmania. Eur. J. Immunol. 2008, 38, 706–719. [Google Scholar] [CrossRef]
- Jayakumar, A.; Castilho, T.M.; Park, E.; Goldsmith-Pestana, K.; Blackwell, J.M.; McMahon-Pratt, D. TLR1/2 Activation during Heterologous Prime-Boost Vaccination (DNA-MVA) Enhances CD8+ T Cell Responses Providing Protection against Leishmania (Viannia). PLoS Negl. Trop. Dis. 2011, 5, e1204. [Google Scholar] [CrossRef]
- Ramos, I.; Alonso, A.; Peris, A.; Marcen, J.M.; Abengozar, M.A.; Alcolea, P.J.; Castillo, J.A.; Larraga, V. Antibiotic Resistance Free Plasmid DNA Expressing LACK Protein Leads towards a Protective Th1 Response against Leishmania Infantum Infection. Vaccine 2009, 27, 6695–6703. [Google Scholar] [CrossRef]
- Bayih, A.G.; Daifalla, N.S.; Gedamu, L. DNA-Protein Immunization Using Leishmania Peroxidoxin-1 Induces a Strong CD4+ T Cell Response and Partially Protects Mice from Cutaneous Leishmaniasis: Role of Fusion Murine Granulocyte-Macrophage Colony-Stimulating Factor DNA Adjuvant. PLoS Negl. Trop. Dis. 2014, 8, e3391. [Google Scholar] [CrossRef]
- Tsagozis, P.; Karagouni, E.; Dotsika, E. Dendritic Cells Pulsed with Peptides of Gp63 Induce Differential Protection against Experimental Cutaneous Leishmaniasis. Int. J. Immunopathol. Pharmacol. 2004, 17, 343–352. [Google Scholar] [CrossRef]
- Remer, K.A.; Apetrei, C.; Schwarz, T.; Linden, C.; Moll, H. Vaccination with Plasmacytoid Dendritic Cells Induces Protection against Infection with Leishmania Major in Mice. Eur. J. Immunol. 2007, 37, 2463–2473. [Google Scholar] [CrossRef]
- Ahuja, S.S.; Reddick, R.L.; Sato, N.; Montalbo, E.; Kostecki, V.; Zhao, W.; Dolan, M.J.; Melby, P.C.; Ahuja, S.K. Dendritic Cell (DC)-Based Anti-Infective Strategies: DCs Engineered to Secrete IL-12 Are a Potent Vaccine in a Murine Model of an Intracellular Infection. J. Immunol. 1999, 163, 3890–3897. [Google Scholar]
- Agallou, M.; Margaroni, M.; Karagouni, E. Cellular Vaccination with Bone Marrow-Derived Dendritic Cells Pulsed with a Peptide of Leishmania Infantum KMP-11 and CpG Oligonucleotides Induces Protection in a Murine Model of Visceral Leishmaniasis. Vaccine 2011, 29, 5053–5064. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, M.; Pal, C.; Ray, M.; Maitra, S.; Mandal, L.; Bandyopadhyay, S. Dendritic Cell-Based Immunotherapy Combined with Antimony-Based Chemotherapy Cures Established Murine Visceral Leishmaniasis. J. Immunol. 2003, 170, 5625–5629. [Google Scholar] [CrossRef] [PubMed]
- Basu, R.; Bhaumik, S.; Haldar, A.K.; Naskar, K.; De, T.; Dana, S.K.; Walden, P.; Roy, S. Hybrid Cell Vaccination Resolves Leishmania Donovani Infection by Eliciting a Strong CD8+ Cytotoxic T-Lymphocyte Response with Concomitant Suppression of Interleukin-10 (IL-10) but Not IL-4 or IL-13. Infect. Immun. 2007, 75, 5956–5966. [Google Scholar] [CrossRef] [PubMed]
- Juncker, A.S.; Larsen, M.V.; Weinhold, N.; Nielsen, M.; Brunak, S.; Lund, O. Systematic Characterisation of Cellular Localisation and Expression Profiles of Proteins Containing MHC Ligands. PLoS ONE 2009, 4, e7448. [Google Scholar] [CrossRef]
- Bassani-Sternberg, M.; Pletscher-Frankild, S.; Jensen, L.J.; Mann, M. Mass Spectrometry of Human Leukocyte Antigen Class I Peptidomes Reveals Strong Effects of Protein Abundance and Turnover on Antigen Presentation. Mol. Cell. Proteom. 2015, 14, 658–673. [Google Scholar] [CrossRef] [Green Version]
- Wolfram, M.; Ilg, T.; Mottram, J.C.; Overath, P. Antigen Presentation by Leishmania Mexicana-infected Macrophages: Activation of Helper T Cells Specific for Amastigote Cysteine Proteinases Requires Intracellular Killing of the Parasites. Eur. J. Immunol. 1995, 25, 1094–1100. [Google Scholar] [CrossRef]
- Wolfram, M.; Fuchs, M.; Wiese, M.; Stierhof, Y.; Overath, P. Antigen Presentation by Leishmania Mexicana-infected Macrophages: Activation of Helper T Cells by a Model Parasite Antigen Secreted into the Parasitophorous Vacuole or Expressed on the Amastigote Surface. Eur. J. Immunol. 1996, 26, 3153–3162. [Google Scholar] [CrossRef]
- Kima, P.E.; Soong, L.; Chicharro, C.; Ruddle, N.H.; McMahon-Pratt, D. Leishmania-infected Macrophages Sequester Endogenously Synthesized Parasite Antigens from Presentation to CD4+ T Cells. Eur. J. Immunol. 1996, 26, 3163–3169. [Google Scholar] [CrossRef]
- Prickett, S.; Gray, P.M.; Colpitts, S.L.; Scott, P.; Kaye, P.M.; Smith, D.F. In Vivo Recognition of Ovalbumin Expressed by Transgenic Leishmania Is Determined by Its Subcellular Localization. J. Immunol. 2006, 176, 4826–4833. [Google Scholar] [CrossRef]
- Ueno, N.; Wilson, M.E. Receptor-Mediated Phagocytosis of Leishmania: Implications for Intracellular Survival. Trends Parasitol. 2012, 28, 335–344. [Google Scholar] [CrossRef]
- Wang, T.T.; Maamary, J.; Tan, G.S.; Bournazos, S.; Davis, C.W.; Krammer, F.; Schlesinger, S.J.; Palese, P.; Ahmed, R.; Ravetch, J.V. Anti-HA Glycoforms Drive B Cell Affinity Selection and Determine Influenza Vaccine Efficacy. Cell 2015, 162, 160–169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stäger, S.; Alexander, J.; Kirby, A.C.; Botto, M.; Van Rooijen, N.; Smith, D.F.; Brombacher, F.; Kaye, P.M. Natural Antibodies and Complement Are Endogenous Adjuvants for Vaccine-Induced CD8+ T-Cell Responses. Nat. Med. 2003, 9, 1287–1292. [Google Scholar] [CrossRef] [PubMed]
- Dameshghi, S.; Zavaran-Hosseini, A.; Soudi, S.; Shirazi, F.J.; Nojehdehi, S.; Hashemi, S.M. Mesenchymal Stem Cells Alter Macrophage Immune Responses to Leishmania Major Infection in Both Susceptible and Resistance Mice. Immunol. Lett. 2016, 170, 15–26. [Google Scholar] [CrossRef] [PubMed]
- Verreck, F.A.W.; De Boer, T.; Langenberg, D.M.L.; Hoeve, M.A.; Kramer, M.; Vaisberg, E.; Kastelein, R.; Kolk, A.; De Waal-Malefyt, R.; Ottenhoff, T.H.M. Human IL-23-Producing Type 1 Macrophages Promote but IL-10-Producing Type 2 Macrophages Subvert Immunity to (Myco)Bacteria. Proc. Natl. Acad. Sci. USA 2004, 101, 4560–4565. [Google Scholar] [CrossRef]
- Duque, G.A.; Fukuda, M.; Turco, S.J.; Stäger, S.; Descoteaux, A. Leishmania Promastigotes Induce Cytokine Secretion in Macrophages through the Degradation of Synaptotagmin XI. J. Immunol. 2014, 193, 2363–2372. [Google Scholar] [CrossRef]
- Vinet, A.F.; Fukuda, M.; Turco, S.J.; Descoteaux, A. The Leishmania Donovani Lipophosphoglycan Excludes the Vesicular Proton-ATPase from Phagosomes by Impairing the Recruitment of Synaptotagmin V. PLoS Pathog. 2009, 5, e1000628. [Google Scholar] [CrossRef]
- Forget, G.; Gregory, D.J.; Olivier, M. Proteasome-Mediated Degradation of STAT1α Following Infection of Macrophages with Leishmania Donovani. J. Biol. Chem. 2005, 280, 30542–30549. [Google Scholar] [CrossRef]
- Späth, G.F.; Schlesinger, P.; Schreiber, R.; Beverley, S.M. A Novel Role for Stat1 in Phagosome Acidification and Natural Host Resistance to Intracellular Infection by Leishmania Major. PLoS Pathog. 2009, 5, e1000381. [Google Scholar] [CrossRef]
- Hsieh, C.-S.; deRoos, P.; Honey, K.; Beers, C.; Rudensky, A.Y. A Role for Cathepsin L and Cathepsin S in Peptide Generation for MHC Class II Presentation. J. Immunol. 2002, 168, 2618–2625. [Google Scholar] [CrossRef]
- Rossman, M.D.; Maida, B.T.; Douglas, S.D. Monocyte-Derived Macrophage and Alveolar Macrophage Fibronectin Production and Cathepsin D Activity. Cell. Immunol. 1990, 126, 268–277. [Google Scholar] [CrossRef]
- Kropf, P.; Freudenberg, M.A.; Modolell, M.; Price, H.P.; Herath, S.; Antoniazi, S.; Galanos, C.; Smith, D.F.; Müller, I. Toll-like Receptor 4 Contributes to Efficient Control of Infection with the Protozoan Parasite Leishmania Major. Infect. Immun. 2004, 72, 1920–1928. [Google Scholar] [CrossRef] [PubMed]
- Liuzzo, J.P.; Petanceska, S.S.; Moscatelli, D.; Devi, L.A. Inflammatory Mediators Regulate Cathepsin S in Macrophages and Microglia: A Role in Attenuating Heparan Sulfate Interactions. Mol. Med. 1999, 5, 320–333. [Google Scholar] [CrossRef] [PubMed]
- Blum, J.S.; Wearsch, P.A.; Cresswell, P. Pathways of Antigen Processing. Annu. Rev. Immunol. 2013, 31, 443–473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Antoine, J.C.; Lang, T.; Prina, E.; Courret, N.; Hellio, R. H-2M Molecules, like MHC Class II Molecules, Are Targeted to Parasitophorous Vacuoles of Leishmania-Infected Macrophages and Internalized by Amastigotes of L. Amazonensis and L. Mexicana. J. Cell Sci. 1999, 112, 2559–2570. [Google Scholar] [PubMed]
- Sercarz, E.E.; Lehmann, P.V.; Ametani, A.; Benichou, G.; Miller, A.; Moudgil, K. Dominance and Crypticity of T Cell Antigenic Determinants. Annu. Rev. Immunol. 1993, 11, 729–766. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; McCluskey, J. Immunodominance and Immunodomination: Critical Factors in Developing Effective CD8+ T-Cell–Based Cancer Vaccines. Adv. Cancer Res. 2006, 95, 203–247. [Google Scholar] [PubMed]
- Luciani, F.; Sanders, M.T.; Oveissi, S.; Pang, K.C.; Chen, W. Increasing Viral Dose Causes a Reversal in CD8 + T Cell Immunodominance during Primary Influenza Infection Due to Differences in Antigen Presentation, T Cell Avidity, and Precursor Numbers. J. Immunol. 2013, 190, 36–47. [Google Scholar] [CrossRef]
- Egui, A.; Ledesma, D.; Pérez-Antón, E.; Montoya, A.; Gómez, I.; Robledo, S.M.; Infante, J.J.; Vélez, I.D.; Lopez, M.C.; Thomas, M.C. Phenotypic and Functional Profiles of Antigen-Specific CD4+ and CD8+ T Cells Associated with Infection Control in Patients with Cutaneous Leishmaniasis. Front. Cell. Infect. Microbiol. 2018, 8, 393. [Google Scholar] [CrossRef]
- Budd, R.C.; Cerottini, J.C.; Horvath, C.; Bron, C.; Pedrazzini, T.; Howe, R.C.; MacDonald, H.R. Distinction of Virgin and Memory T Lymphocytes. Stable Acquisition of the Pgp-1 Glycoprotein Concomitant with Antigenic Stimulation. J. Immunol. 1987, 138, 3120–3129. [Google Scholar]
- Mackay, C.R.; Marston, W.L.; Dudler, L. Naive and Memory T Cells Show Distinct Pathways of Lymphocyte Recirculation. J. Exp. Med. 1990, 171, 801–817. [Google Scholar] [CrossRef]
- Nyambura, L.W.; Jarmalavicius, S.; Walden, P. Impact of Leishmania Donovani Infection on the HLA I Self Peptide Repertoire of Human Macrophages. PLoS ONE 2018, 13, e0200297. [Google Scholar] [CrossRef] [PubMed]
- Roy, K.; Naskar, K.; Ghosh, M.; Roy, S. Class II MHC/Peptide Interaction in Leishmania Donovani Infection: Implications in Vaccine Design. J. Immunol. 2014, 192, 5873–5880. [Google Scholar] [CrossRef] [PubMed]
- Weaver, J.M.; Lazarski, C.A.; Richards, K.A.; Chaves, F.A.; Jenks, S.A.; Menges, P.R.; Sant, A.J. Immunodominance of CD4 T Cells to Foreign Antigens Is Peptide Intrinsic and Independent of Molecular Context: Implications for Vaccine Design. J. Immunol. 2008, 181, 3039–3048. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Williams, K.P.; Chang, Y.-H.; Smith, J.A. Immunodominance: A Single Amino Acid Substitution within an Antigenic Site Alters Intramolecular Selection of T Cell Determinants. J. Immunol. 1993, 151, 1852–1858. [Google Scholar]
- Mazor, R.; Vassall, A.N.; Eberle, J.A.; Beers, R.; Weldon, J.E.; Venzon, D.J.; Tsang, K.Y.; Benhar, I.; Pastan, I. Identification and Elimination of an Immunodominant T-Cell Epitope in Recombinant Immunotoxins Based on Pseudomonas Exotoxin A. Proc. Natl. Acad. Sci. USA 2012, 109, E3597–E3603. [Google Scholar] [CrossRef] [PubMed]
- Mazor, R.; Eberle, J.A.; Hu, X.; Vassall, A.N.; Onda, M.; Beers, R.; Lee, E.C.; Kreitman, R.J.; Lee, B.; Baker, D. Recombinant Immunotoxin for Cancer Treatment with Low Immunogenicity by Identification and Silencing of Human T-Cell Epitopes. Proc. Natl. Acad. Sci. USA 2014, 111, 8571–8576. [Google Scholar] [CrossRef] [PubMed]
- Schneider, S.C.; Ohmen, J.; Fosdick, L.; Gladstone, B.; Guo, J.; Ametani, A.; Sercarz, E.E.; Deng, H. Cutting Edge: Introduction of an Endopeptidase Cleavage Motif into a Determinant Flanking Region of Hen Egg Lysozyme Results in Enhanced T Cell Determinant Display. J. Immunol. 2000, 165, 20–23. [Google Scholar] [CrossRef] [Green Version]
- Moss, D.L.; Park, H.-W.; Mettu, R.R.; Landry, S.J. Deimmunizing Substitutions in Pseudomonas Exotoxin Domain III Perturb Antigen Processing without Eliminating T-Cell Epitopes. J. Biol. Chem. 2019, 294, 4667–4681. [Google Scholar] [CrossRef]
- Kim, A.; Boronina, T.N.; Cole, R.N.; Darrah, E.; Sadegh-Nasseri, S. Distorted Immunodominance by Linker Sequences or Other Epitopes from a Second Protein Antigen during Antigen-Processing. Sci. Rep. 2017, 7, 46418. [Google Scholar] [CrossRef]
- Laffitte, M.-C.N.; Leprohon, P.; Papadopoulou, B.; Ouellette, M. Plasticity of the Leishmania Genome Leading to Gene Copy Number Variations and Drug Resistance. F1000Research 2016, 5, 2350. [Google Scholar] [CrossRef]
- Iantorno, S.A.; Durrant, C.; Khan, A.; Sanders, M.J.; Beverley, S.M.; Warren, W.C.; Berriman, M.; Sacks, D.L.; Cotton, J.A.; Grigg, M.E. Gene Expression in Leishmania Is Regulated Predominantly by Gene Dosage. MBio 2017, 8, e01393-17. [Google Scholar] [CrossRef]
- Monks, C.R.F.; Freiberg, B.A.; Kupfer, H.; Sciaky, N.; Kupfer, A. Three-Dimensional Segregation of Supramolecular Activation Clusters in T Cells. Nature 1998, 395, 82–86. [Google Scholar] [CrossRef] [PubMed]
- Grakoui, A.; Bromley, S.K.; Sumen, C.; Davis, M.M.; Shaw, A.S.; Allen, P.M.; Dustin, M.L. The Immunological Synapse: A Molecular Machine Controlling T Cell Activation. Science 1999, 285, 221–227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hosseini, B.H.; Louban, I.; Djandji, D.; Wabnitz, G.H.; Deeg, J.; Bulbuc, N.; Samstag, Y.; Gunzer, M.; Spatz, J.P.; Hämmerling, G.J. Immune Synapse Formation Determines Interaction Forces between T Cells and Antigen-Presenting Cells Measured by Atomic Force Microscopy. Proc. Natl. Acad. Sci. USA 2009, 106, 17852–17857. [Google Scholar] [CrossRef] [PubMed]
- Dustin, M.L.; Bromley, S.K.; Kan, Z.; Peterson, D.A.; Unanue, E.R. Antigen Receptor Engagement Delivers a Stop Signal to Migrating T Lymphocytes. Proc. Natl. Acad. Sci. USA 1997, 94, 3909–3913. [Google Scholar] [CrossRef]
- Park, E.J.; Peixoto, A.; Imai, Y.; Goodarzi, A.; Cheng, G.; Carman, C.V.; von Andrian, U.H.; Shimaoka, M. Distinct Roles for LFA-1 Affinity Regulation during T-Cell Adhesion, Diapedesis, and Interstitial Migration in Lymph Nodes. Blood 2010, 115, 1572–1581. [Google Scholar] [CrossRef]
- Zhang, W.; Trible, R.P.; Samelson, L.E. LAT Palmitoylation: Its Essential Role in Membrane Microdomain Targeting and Tyrosine Phosphorylation during T Cell Activation. Immunity 1998, 9, 239–246. [Google Scholar] [CrossRef] [Green Version]
- Bi, K.; Tanaka, Y.; Coudronniere, N.; Sugie, K.; Hong, S.; van Stipdonk, M.J.B.; Altman, A. Antigen-Induced Translocation of PKC-θ to Membrane Rafts Is Required for T Cell Activation. Nat. Immunol. 2001, 2, 556–563. [Google Scholar] [CrossRef]
- Molina, T.J.; Kishihara, K.; Siderovskid, D.P.; Van Ewijk, W.; Narendran, A.; Timms, E.; Wakeham, A.; Paige, C.J.; Hartmann, K.U.; Veillette, A.; et al. Profound Block in Thymocyte Development in Mice Lacking P56lck. Nature 1992, 357, 161–164. [Google Scholar] [CrossRef]
- Rub, A.; Dey, R.; Jadhav, M.; Kamat, R.; Chakkaramakkil, S.; Majumdar, S.; Mukhopadhyaya, R.; Saha, B. Cholesterol Depletion Associated with Leishmania Major Infection Alters Macrophage CD40 Signalosome Composition and Effector Function. Nat. Immunol. 2009, 10, 273–280. [Google Scholar] [CrossRef]
- Meier, C.L.; Svensson, M.; Kaye, P.M. Leishmania -Induced Inhibition of Macrophage Antigen Presentation Analyzed at the Single-Cell Level. J. Immunol. 2003, 171, 6706–6713. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, J.; Das, S.; Guha, R.; Ghosh, D.; Naskar, K.; Das, A.; Roy, S. Hyperlipidemia Offers Protection against Leishmania Donovani Infection: Role of Membrane Cholesterol. J. Lipid Res. 2012, 53, 2560–2572. [Google Scholar] [CrossRef] [PubMed]
- Albrecht, I.; Gatfield, J.; Mini, T.; Jeno, P.; Pieters, J. Essential Role for Cholesterol in the Delivery of Exogenous Antigens to the MHC Class I-Presentation Pathway. Int. Immunol. 2006, 18, 755–765. [Google Scholar] [CrossRef] [PubMed]
- Roy, K.; Mandloi, S.; Chakrabarti, S.; Roy, S. Cholesterol Corrects Altered Conformation of MHC-II Protein in Leishmania Donovani Infected Macrophages: Implication in Therapy. PLoS Negl. Trop. Dis. 2016, 10, e0004710. [Google Scholar] [CrossRef]
- Semini, G.; Paape, D.; Paterou, A.; Schroeder, J.; Barrios-Llerena, M.; Aebischer, T. Changes to Cholesterol Trafficking in Macrophages by Leishmania Parasites Infection. Microbiologyopen 2017, 6, e00469. [Google Scholar] [CrossRef]
- Banerjee, S.; Ghosh, J.; Sen, S.; Guha, R.; Dhar, R.; Ghosh, M.; Datta, S.; Raychaudhury, B.; Naskar, K.; Haldar, A.K.; et al. Designing Therapies against Experimental Visceral Leishmaniasis by Modulating the Membrane Fluidity of Antigen-Presenting Cells. Infect. Immun. 2009, 77, 2330–2342. [Google Scholar] [CrossRef] [Green Version]
- Reiner, N.E.; Ng, W.; McMaster, W.R. Parasite-Accessory Cell Interactions in Murine Leishmaniasis. II. Leishmania Donovani Suppresses Macrophage Expression of Class I and Class II Major Histocompatibility Complex Gene Products. J. Immunol. 1987, 138, 1926–1932. [Google Scholar]
- Muraille, E.; De Trez, C.; Pajak, B.; Torrentera, F.A.; De Baetselier, P.; Leo, O.; Carlier, Y. Amastigote Load and Cell Surface Phenotype of Infected Cells from Lesions and Lymph Nodes of Susceptible and Resistant Mice Infected with Leishmania Major. Infect. Immun. 2003, 71, 2704–2715. [Google Scholar] [CrossRef]
- Silverman, J.M.; Clos, J.; Horakova, E.; Wang, A.Y.; Wiesgigl, M.; Kelly, I.; Lynn, M.A.; McMaster, W.R.; Foster, L.J.; Levings, M.K.; et al. Leishmania Exosomes Modulate Innate and Adaptive Immune Responses through Effects on Monocytes and Dendritic Cells. J. Immunol. 2010, 185, 5011–5022. [Google Scholar] [CrossRef] [Green Version]
- Pinheiro, N.F.; Hermida, M.D.R.; Macedo, M.P.; Mengel, J.; Bafica, A.; dos-Santos, W.L.C. Leishmania Infection Impairs Β1-Integrin Function and Chemokine Receptor Expression in Mononuclear Phagocytes. Infect. Immun. 2006, 74, 3912–3921. [Google Scholar] [CrossRef]
- Rosenthal, L.A.; Sutterwala, F.S.; Kehrli, M.E.; Mosser, D.M. Leishmania Major-Human Macrophage Interactions: Cooperation between Mac-1 (CD11b/CD18) and Complement Receptor Type 1 (CD35) in Promastigote Adhesion. Infect. Immun. 1996, 64, 2206–2215. [Google Scholar] [PubMed]
- de Menezes, J.P.B.; Koushik, A.; Das, S.; Guven, C.; Siegel, A.; Laranjeira-Silva, M.F.; Losert, W.; Andrews, N.W. Leishmania Infection Inhibits Macrophage Motility by Altering F-Actin Dynamics and the Expression of Adhesion Complex Proteins. Cell. Microbiol. 2017, 19, e12668. [Google Scholar] [CrossRef] [PubMed]
- Bachmann, M.F.; Sebzda, E.; Kündig, T.M.; Shahinian, A.; Speiser, D.E.; Mak, T.W.; Ohashi, P.S. T Cell Responses Are Governed by Avidity and Co-Stimulatory Thresholds. Eur. J. Immunol. 1996, 26, 2017–2022. [Google Scholar] [CrossRef] [PubMed]
- Brown, J.A.; Titus, R.G.; Nabavi, N.; Glimcher, L.H. Blockade of CD86 Ameliorates Leishmania Major Infection by Down- Regulating the Th2 Response. J. Infect. Dis. 1996, 174, 1303–1308. [Google Scholar] [CrossRef]
- Li, J.G.; Du, Y.M.; Yan, Z.D.; Yan, J.; Zhuansun, Y.X.; Chen, R.; Zhang, W.; Feng, S.L.; Ran, P.X. CD80 and CD86 Knockdown in Dendritic Cells Regulates Th1/Th2 Cytokine Production in Asthmatic Mice. Exp. Ther. Med. 2016, 11, 878–884. [Google Scholar] [CrossRef]
- Brodskyn, C.I.; DeKrey, G.K.; Titus, R.G. Influence of Costimulatory Molecules on Immune Response to Leishmania Major by Human Cells in Vitro. Infect. Immun. 2001, 69, 665–672. [Google Scholar] [CrossRef]
- Freedman, A.S.; Freeman, G.J.; Rhynhart, K.; Nadler, L.M. Selective Induction of B7/BB-1 on Interferon-γ Stimulated Monocytes: A Potential Mechanism for Amplification of T Cell Activation through the CD28 Pathway. Cell. Immunol. 1991, 137, 429–437. [Google Scholar] [CrossRef]
- Kiener, P.A.; Moran-Davis, P.; Rankin, B.M.; Wahl, A.F.; Aruffo, A.; Hollenbaugh, D. Stimulation of CD40 with Purified Soluble Gp39 Induces Proinflammatory Responses in Human Monocytes. J. Immunol. 1995, 155, 4917–4925. [Google Scholar]
- Caux, C.; Massacrier, C.; Vanbervliet, B.; Dubois, B.; Van Kooten, C.; Durand, I.; Banchereau, J. Activation of Human Dendritic Cells through CD40 Cross-Linking. J. Exp. Med. 1994, 180, 1263–1272. [Google Scholar] [CrossRef]
- Saha, B.; Das, G.; Vohra, H.; Ganguly, N.K.; Mishra, G.C. Macrophage-T Cell Interaction in Experimental Visceral Leishmaniasis: Failure to Express Costimulatory Molecules on Leishmania-infected Macrophages and Its Implication in the Suppression of Cell-mediated Immunity. Eur. J. Immunol. 1995, 25, 2492–2498. [Google Scholar] [CrossRef]
- Walunas, T.L.; Lenschow, D.J.; Bakker, C.Y.; Linsley, P.S.; Freeman, G.J.; Green, J.M.; Thompson, C.B.; Bluestone, J.A. CTLA-4 Can Function as a Negative Regulator of T Cell Activation. Immunity 1994, 1, 405–413. [Google Scholar] [CrossRef]
- Perez, V.L.; Van Parijs, L.; Biuckians, A.; Zheng, X.X.; Strom, T.B.; Abbas, A.K. Induction of Peripheral T Cell Tolerance in Vivo Requires CTLA-4 Engagement. Immunity 1997, 6, 411–417. [Google Scholar] [CrossRef]
- Lindsten, T.; Lee, K.P.; Harris, E.S.; Petryniak, B.; Craighead, N.; Reynolds, P.J.; Lombard, D.B.; Freeman, G.J.; Nadler, L.M.; Gray, G.S. Characterization of CTLA-4 Structure and Expression on Human T Cells. J. Immunol. 1993, 151, 3489–3499. [Google Scholar] [PubMed]
- Kaye, P.M.; Murphy, M.L.; Engwerda, C.R.; Gorak, P.M.A. B7-2 Blockade Enhances T Cell Responses to Leishmania Donovani. J. Immunol. 1997, 159, 4460–4466. [Google Scholar]
- Murphy, M.L.; Cotterell, S.E.; Gorak, P.M.; Engwerda, C.R.; Kaye, P.M. Blockade of CTLA-4 Enhances Host Resistance to the Intracellular Pathogen, Leishmania Donovani. J. Immunol. 1998, 161, 4153–4160. [Google Scholar]
- Sheppard, K.A.; Fitz, L.J.; Lee, J.M.; Benander, C.; George, J.A.; Wooters, J.; Qiu, Y.; Jussif, J.M.; Carter, L.L.; Wood, C.R.; et al. PD-1 Inhibits T-Cell Receptor Induced Phosphorylation of the ZAP70/CD3ζ Signalosome and Downstream Signaling to PKCθ. FEBS Lett. 2004, 574, 37–41. [Google Scholar] [CrossRef]
- Roy, S.; Gupta, P.; Palit, S.; Basu, M.; Ukil, A.; Das, P.K. The Role of PD-1 in Regulation of Macrophage Apoptosis and Its Subversion by Leishmania Donovani. Clin. Transl. Immunol. 2017, 6, e137. [Google Scholar] [CrossRef]
- Abe, T.; Tanaka, Y.; Piao, J.; Tanimine, N.; Oue, N.; Hinoi, T.; Garcia, N.V.; Miyasaka, M.; Matozaki, T.; Yasui, W.; et al. Signal Regulatory Protein Alpha Blockade Potentiates Tumoricidal Effects of Macrophages on Gastroenterological Neoplastic Cells in Syngeneic Immunocompetent Mice. Ann. Gastroenterol. Surg. 2018, 2, 451–462. [Google Scholar] [CrossRef]
- Ontoria, E.; Hernández-Santana, Y.E.; González-García, A.C.; López, M.C.; Valladares, B.; Carmelo, E. Transcriptional Profiling of Immune-Related Genes in Leishmania Infantum-Infected Mice: Identification of Potential Biomarkers of Infection and Progression of Disease. Front. Cell. Infect. Microbiol. 2018, 8, 197. [Google Scholar] [CrossRef]
- Sanderson, N.S.R.; Puntel, M.; Kroeger, K.M.; Bondale, N.S.; Swerdlow, M.; Iranmanesh, N.; Yagita, H.; Ibrahim, A.; Castro, M.G.; Lowenstein, P.R. Cytotoxic Immunological Synapses Do Not Restrict the Action of Interferon-γ to Antigenic Target Cells. Proc. Natl. Acad. Sci. USA 2012, 109, 7835–7840. [Google Scholar] [CrossRef]
- Huse, M.; Lillemeier, B.F.; Kuhns, M.S.; Chen, D.S.; Davis, M.M. T Cells Use Two Directionally Distinct Pathways for Cytokine Secretion. Nat. Immunol. 2006, 7, 247–255. [Google Scholar] [CrossRef] [PubMed]
- Chemin, K.; Bohineust, A.; Dogniaux, S.; Tourret, M.; Guégan, S.; Miro, F.; Hivroz, C. Cytokine Secretion by CD4+ T Cells at the Immunological Synapse Requires Cdc42-Dependent Local Actin Remodeling but Not Microtubule Organizing Center Polarity. J. Immunol. 2012, 189, 2159–2168. [Google Scholar] [CrossRef] [PubMed]
- Fooksman, D.R.; Vardhana, S.; Vasiliver-Shamis, G.; Liese, J.; Blair, D.A.; Waite, J.; Sacristán, C.; Victora, G.D.; Zanin-Zhorov, A.; Dustin, M.L. Functional Anatomy of T Cell Activation and Synapse Formation. Annu. Rev. Immunol. 2010, 28, 79–105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Philipsen, L.; Engels, T.; Schilling, K.; Gurbiel, S.; Fischer, K.D.; Tedford, K.; Schraven, B.; Gunzer, M.; Reichardt, P. Multimolecular Analysis of Stable Immunological Synapses Reveals Sustained Recruitment and Sequential Assembly of Signaling Clusters. Mol. Cell. Proteom. 2013, 12, 2551–2567. [Google Scholar] [CrossRef] [Green Version]
- Malherbe, L.; Filippi, C.; Julia, V.; Foucras, G.; Moro, M.; Appel, H.; Wucherpfennig, K.; Guéry, J.C.; Glaichenhaus, N. Selective Activation and Expansion of High-Affinity CD4+ T Cells in Resistant Mice upon Infection with Leishmania Major. Immunity 2000, 13, 771–782. [Google Scholar] [CrossRef]
- Pentcheva-Hoang, T.; Egen, J.G.; Wojnoonski, K.; Allison, J.P. B7-1 and B7-2 Selectively Recruit CTLA-4 and CD28 to the Immunological Synapse. Immunity 2004, 21, 401–413. [Google Scholar] [CrossRef] [Green Version]
- Gannavaram, S.; Bhattacharya, P.; Ismail, N.; Kaul, A.; Singh, R.; Nakhasi, H.L. Modulation of Innate Immune Mechanisms to Enhance Leishmania Vaccine-Induced Immunity: Role of Coinhibitory Molecules. Front. Immunol. 2016, 7, 187. [Google Scholar] [CrossRef] [Green Version]
- Gomes, N.A.; Gattass, C.R.; Barreto-de-Souza, V.; Wilson, M.E.; DosReis, G.A. TGF-β Mediates CTLA-4 Suppression of Cellular Immunity in Murine Kalaazar. J. Immunol. 2000, 164, 2001–2008. [Google Scholar] [CrossRef]
- He, X.; Holmes, T.H.; Mahmood, K.; Kemble, G.W.; Dekker, C.L.; Arvin, A.M.; Greenberg, H.B. Phenotypic Changes in Influenza-Specific CD8 + T Cells after Immunization of Children and Adults with Influenza Vaccines. J. Infect. Dis. 2008, 197, 803–811. [Google Scholar] [CrossRef]
- Viganò, S.; Utzschneider, D.T.; Perreau, M.; Pantaleo, G.; Zehn, D.; Harari, A. Functional Avidity: A Measure to Predict the Efficacy of Effector T Cells? Clin. Dev. Immunol. 2012, 2012. [Google Scholar] [CrossRef]
- Betts, M.R.; Price, D.A.; Brenchley, J.M.; Loré, K.; Guenaga, F.J.; Smed-Sorensen, A.; Ambrozak, D.R.; Migueles, S.A.; Connors, M.; Roederer, M.; et al. The Functional Profile of Primary Human Antiviral CD8 + T Cell Effector Activity Is Dictated by Cognate Peptide Concentration. J. Immunol. 2004, 172, 6407–6417. [Google Scholar] [CrossRef] [PubMed]
- Langenkamp, A.; Casorati, G.; Garavaglia, C.; Dellabona, P.; Lanzavecchia, A.; Sallusto, F. T Cell Priming by Dendritic Cells: Thresholds for Proliferation, Differentiation and Death and Intraclonal Functional Diversification. Eur. J. Immunol. 2002, 32, 2046–2054. [Google Scholar] [CrossRef]
- Wooldridge, L.; Lissina, A.; Vernazza, J.; Gostick, E.; Laugel, B.; Hutchinson, S.L.; Mirza, F.; Dunbar, P.R.; Boulter, J.M.; Glick, M.; et al. Enhanced Immunogenicity of CTL Antigens through Mutation of the CD8 Binding MHC Class I Invariant Region. Eur. J. Immunol. 2007, 37, 1323–1333. [Google Scholar] [CrossRef] [PubMed]
- Azzam, H.S.; Grinberg, A.; Lui, K.; Shen, H.; Shores, E.W.; Love, P.E. CD5 Expression Is Developmentally Regulated by T Cell Receptor (TCR) Signals and TCR Avidity. J. Exp. Med. 1998, 188, 2301–2311. [Google Scholar] [CrossRef] [PubMed]
- Ebert, P.J.R.; Jiang, S.; Xie, J.; Li, Q.-J.; Davis, M.M. An Endogenous Positively Selecting Peptide Enhances Mature T Cell Responses and Becomes an Autoantigen in the Absence of MicroRNA MiR-181a. Nat. Immunol. 2009, 10, 1162. [Google Scholar] [CrossRef] [PubMed]
- Pircher, H.; Rohrer, U.H.; Moskophidis, D.; Zinkernagel, R.M.; Hengartner, H. Lower Receptor Avidity Required for Thymic Clonal Deletion than for Effector T-Cell Function. Nature 1991, 351, 482–485. [Google Scholar] [CrossRef]
- Liu, H.; Cao, A.T.; Feng, T.; Li, Q.; Zhang, W.; Yao, S.; Dann, S.M.; Elson, C.O.; Cong, Y. TGF-β Converts Th1 Cells into Th17 Cells through Stimulation of Runx1 Expression. Eur. J. Immunol. 2015, 45, 1010–1018. [Google Scholar] [CrossRef]
- Li, M.O.; Flavell, R.A. TGF-β: A Master of All T Cell Trades. Cell 2008, 134, 392–404. [Google Scholar] [CrossRef] [PubMed]
- Sakaguchi, S.; Yamaguchi, T.; Nomura, T.; Ono, M. Regulatory T Cells and Immune Tolerance. Cell 2008, 133, 775–787. [Google Scholar] [CrossRef] [Green Version]
- Ziegler, S.F.; Buckner, J.H. FOXP3 and the Regulation of Treg/Th17 Differentiation. Microbes Infect. 2009, 11, 594–598. [Google Scholar] [CrossRef]
- Xu, T.; Stewart, K.M.; Wang, X.; Liu, K.; Xie, M.; Ryu, J.K.; Li, K.; Ma, T.; Wang, H.; Ni, L. Metabolic Control of T H 17 and Induced T Reg Cell Balance by an Epigenetic Mechanism. Nature 2017, 548, 228. [Google Scholar] [CrossRef] [PubMed]
- Wherry, E.J. T Cell Exhaustion. Nat. Immunol. 2011, 12, 492–499. [Google Scholar] [CrossRef]
- Barber, D.L.; Wherry, E.J.; Masopust, D.; Zhu, B.; Allison, J.P.; Sharpe, A.H.; Freeman, G.J.; Ahmed, R. Restoring Function in Exhausted CD8 T Cells during Chronic Viral Infection. Nature 2006, 439, 682. [Google Scholar] [CrossRef] [PubMed]
- Blackburn, S.D.; Shin, H.; Haining, W.N.; Zou, T.; Workman, C.J.; Polley, A.; Betts, M.R.; Freeman, G.J.; Vignali, D.A.A.; Wherry, E.J. Coregulation of CD8+ T Cell Exhaustion by Multiple Inhibitory Receptors during Chronic Viral Infection. Nat. Immunol. 2009, 10, 29. [Google Scholar] [CrossRef] [PubMed]
- Mescher, M.F.; Curtsinger, J.M.; Agarwal, P.; Casey, K.A.; Gerner, M.; Hammerbeck, C.D.; Popescu, F.; Xiao, Z. Signals Required for Programming Effector and Memory Development by CD8+ T Cells. Immunol. Rev. 2006, 211, 81–92. [Google Scholar] [CrossRef] [PubMed]
- Tough, D.F.; Sprent, J. Turnover of Naive-and Memory-Phenotype T Cells. J. Exp. Med. 1994, 179, 1127–1135. [Google Scholar] [CrossRef]
- Munn, D.H.; Sharma, M.D.; Baban, B.; Harding, H.P.; Zhang, Y.; Ron, D.; Mellor, A.L. GCN2 Kinase in T Cells Mediates Proliferative Arrest and Anergy Induction in Response to Indoleamine 2, 3-Dioxygenase. Immunity 2005, 22, 633–642. [Google Scholar] [CrossRef] [PubMed]
- Sitkovsky, M.V. T Regulatory Cells: Hypoxia-Adenosinergic Suppression and Re-Direction of the Immune Response. Trends Immunol. 2009, 30, 102–108. [Google Scholar] [CrossRef]
- Mueller, D.L. Mechanisms Maintaining Peripheral Tolerance. Nat. Immunol. 2010, 11, 21. [Google Scholar] [CrossRef]
- Fallarino, F.; Grohmann, U.; Puccetti, P. Indoleamine 2,3-Dioxygenase: From Catalyst to Signaling Function. Eur. J. Immunol. 2012, 42, 1932–1937. [Google Scholar] [CrossRef]
- Deaglio, S.; Dwyer, K.M.; Gao, W.; Friedman, D.; Usheva, A.; Erat, A.; Chen, J.F.; Enjyoji, K.; Linden, J.; Oukka, M.; et al. Adenosine Generation Catalyzed by CD39 and CD73 Expressed on Regulatory T Cells Mediates Immune Suppression. J. Exp. Med. 2007, 204, 1257–1265. [Google Scholar] [CrossRef] [PubMed]
- Wing, K.; Onishi, Y.; Prieto-Martin, P.; Yamaguchi, T.; Miyara, M.; Fehervari, Z.; Nomura, T.; Sakaguchi, S. CTLA-4 Control over Foxp3+ Regulatory T Cell Function. Science 2008, 322, 271–275. [Google Scholar] [CrossRef] [PubMed]
- Makala, L.H.C.; Baban, B.; Lemos, H.; El-Awady, A.R.; Chandler, P.R.; Hou, D.Y.; Munn, D.H.; Mellor, A.L. Leishmania Major Attenuates Host Immunity by Stimulating Local Indoleamine 2,3-Dioxygenase Expression. J. Infect. Dis. 2011, 203, 715–725. [Google Scholar] [CrossRef] [PubMed]
- Figueiredo, A.B.; Serafim, T.D.; Marques-da-Silva, E.A.; Meyer-Fernandes, J.R.; Afonso, L.C.C. Leishmania Amazonensis Impairs DC Function by Inhibiting CD40 Expression via A 2B Adenosine Receptor Activation. Eur. J. Immunol. 2012, 42, 1203–1215. [Google Scholar] [CrossRef] [PubMed]
- Joshi, T.; Rodriguez, S.; Perovic, V.; Cockburn, I.A.; Stäger, S. B7-H1 Blockade Increases Survival of Dysfunctional CD8+ T Cells and Confers Protection against Leishmania Donovaniinfections. PLoS Pathog. 2009, 5, e1000431. [Google Scholar] [CrossRef]
- Pinheiro, R.O.; Pinto, E.F.; Benedito, A.B.; Lopes, U.G.; Rossi-Bergmann, B. The T-Cell Anergy Induced by Leishmania Amazonensis Antigens Is Related with Defective Antigen Presentation and Apoptosis. An. Acad. Bras. Cienc. 2004, 76, 519–527. [Google Scholar] [CrossRef]
- Krammer, P.H.; Arnold, R.; Lavrik, I.N. Life and Death in Peripheral T Cells. Nat. Rev. Immunol. 2007, 7, 532–542. [Google Scholar] [CrossRef]
- Woodland, D.L. Jump-Starting the Immune System: Prime-Boosting Comes of Age. Trends Immunol. 2004, 25, 98–104. [Google Scholar] [CrossRef]
- Masopust, D.; Ha, S.-J.; Vezys, V.; Ahmed, R. Stimulation History Dictates Memory CD8 T Cell Phenotype: Implications for Prime-Boost Vaccination. J. Immunol. 2006, 177, 831–839. [Google Scholar] [CrossRef] [Green Version]
- Wirth, T.C.; Xue, H.H.; Rai, D.; Sabel, J.T.; Bair, T.; Harty, J.T.; Badovinac, V.P. Repetitive Antigen Stimulation Induces Stepwise Transcriptome Diversification but Preserves a Core Signature of Memory CD8+ T Cell Differentiation. Immunity 2010, 33, 128–140. [Google Scholar] [CrossRef] [Green Version]
- Sallusto, F.; Lanzavecchia, A.; Araki, K.; Ahmed, R. From Vaccines to Memory and Back. Immunity 2010, 33, 451–463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Butler, N.S.; Nolz, J.C.; Harty, J.T. Immunologic Considerations for Generating Memory CD8 T Cells through Vaccination. Cell. Microbiol. 2011, 13, 925–933. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, V.; Cordeiro-da-Silva, A.; Laforge, M.; Ouaissi, A.; Akharid, K.; Silvestre, R.; Estaquier, J. Impairment of T Cell Function in Parasitic Infections. PLoS Negl. Trop. Dis. 2014, 8, e2567. [Google Scholar] [CrossRef] [PubMed]
- Haring, J.S.; Badovinac, V.P.; Harty, J.T. Inflaming the CD8+ T Cell Response. Immunity 2006, 25, 19–29. [Google Scholar] [CrossRef] [Green Version]
- Matte, C.; Descoteaux, A. Leishmania Donovani Amastigotes Impair Gamma Interferon-Induced STAT1α Nuclear Translocation by Blocking the Interaction between STAT1α and Importin-A5. Infect. Immun. 2010, 78, 3736–3743. [Google Scholar] [CrossRef]
- Bhardwaj, N.; Rosas, L.E.; Lafuse, W.P.; Satoskar, A.R. Leishmania Inhibits STAT1-Mediated IFN-γ Signaling in Macrophages: Increased Tyrosine Phosphorylation of Dominant Negative STAT1β by Leishmania Mexicana. Int. J. Parasitol. 2005, 35, 75–82. [Google Scholar] [CrossRef]
- Ray, M.; Gam, A.A.; Boykins, R.A.; Kenney, R.T. Inhibition of Interferon-γ Signaling by Leishmania Donovani. J. Infect. Dis. 2000, 181, 1121–1128. [Google Scholar] [CrossRef]
- Blanchette, J.; Racette, N.; Faure, R.; Siminovitch, K.A.; Olivier, M. Leishmania-Induced Increases in Activation of Macrophage SHP-1 Tyrosine Phosphatase Are Associated with Impaired IFN-γ-Triggered JAK2 Activation. Eur. J. Immunol. 1999, 29, 3737–3744. [Google Scholar] [CrossRef]
- Nandan, D.; Reiner, N.E. Attenuation of Gamma Interferon-Induced Tyrosine Phosphorylation in Mononuclear Phagocytes Infected with Leishmania Donovani: Selective Inhibition of Signaling through Janus Kinases and Stat1. Infect. Immun. 1995, 63, 4495–4500. [Google Scholar]
- Cecchinato, V.; Bernasconi, E.; Speck, R.F.; Proietti, M.; Sauermann, U.; D’Agostino, G.; Danelon, G.; Jost, T.R.; Grassi, F.; Raeli, L. Impairment of CCR6+ and CXCR3+ Th Cell Migration in HIV-1 Infection Is Rescued by Modulating Actin Polymerization. J. Immunol. 2017, 198, 184–195. [Google Scholar] [CrossRef]
- Salgado-Pabón, W.; Celli, S.; Arena, E.T.; Nothelfer, K.; Roux, P.; Sellge, G.; Frigimelica, E.; Bousso, P.; Sansonetti, P.J.; Phalipon, A. Shigella Impairs T Lymphocyte Dynamics in Vivo. Proc. Natl. Acad. Sci. USA 2013, 110, 4458–4463. [Google Scholar] [CrossRef] [PubMed]
- Steigerwald, M.; Moll, H. Leishmania Major Modulates Chemokine and Chemokine Receptor Expression by Dendritic Cells and Affects Their Migratory Capacity. Infect. Immun. 2005, 73, 2564–2567. [Google Scholar] [CrossRef] [PubMed]
- Hermida, M.D.R.; Doria, P.G.; Taguchi, A.M.P.; Mengel, J.O.; dos-Santos, W.L.C. Leishmania Amazonensis Infection Impairs Dendritic Cell Migration from the Inflammatory Site to the Draining Lymph Node. BMC Infect. Dis. 2014, 14, 450. [Google Scholar] [CrossRef] [PubMed]
- Marr, A.K.; MacIsaac, J.L.; Jiang, R.; Airo, A.M.; Kobor, M.S.; McMaster, W.R. Leishmania Donovani Infection Causes Distinct Epigenetic DNA Methylation Changes in Host Macrophages. PLoS Pathog. 2014, 10, e1004419. [Google Scholar] [CrossRef]
- Uzonna, J.E.; Wei, G.; Yurkowski, D.; Bretscher, P. Immune Elimination of Leishmania Major in Mice: Implications for Immune Memory, Vaccination, and Reactivation Disease. J. Immunol. 2001, 167, 6967–6974. [Google Scholar] [CrossRef]
- Zaph, C.; Uzonna, J.; Beverley, S.M.; Scott, P. Central Memory T Cells Mediate Long-Term Immunity to Leishmania Major in the Absence of Persistent Parasites. Nat. Med. 2004, 10, 1104. [Google Scholar] [CrossRef]
- Peters, N.C.; Kimblin, N.; Secundino, N.; Kamhawi, S.; Lawyer, P.; Sacks, D.L. Vector Transmission of Leishmania Abrogates Vaccine-Induced Protective Immunity. PLoS Pathog. 2009, 5, e1000484. [Google Scholar] [CrossRef]
- Gomes, R.; Oliveira, F.; Teixeira, C.; Meneses, C.; Gilmore, D.C.; Elnaiem, D.-E.; Kamhawi, S.; Valenzuela, J.G. Immunity to Sand Fly Salivary Protein LJM11 Modulates Host Response to Vector-Transmitted Leishmania Conferring Ulcer-Free Protection. J. Investig. Dermatol. 2012, 132, 2735–2743. [Google Scholar] [CrossRef] [Green Version]
- Glennie, N.D.; Yeramilli, V.A.; Beiting, D.P.; Volk, S.W.; Weaver, C.T.; Scott, P. Skin-Resident Memory CD4+ T Cells Enhance Protection against Leishmania Major Infection. J. Exp. Med. 2015, 212, 1405–1414. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| VACCINE CANDIDATE | STRAIN (CHALLENGE) | ROI | ADJUVANT | OUTCOME | DRAWBACK(S) | REF |
|---|---|---|---|---|---|---|
| 1ST GENERATION VACCINES | ||||||
| Leishmanization (LZ) | L. major | S.C | -- | Higher success rate in Uzbekistan & Iran | Loss of infectivity, safety & ethical issues | [17] |
| Killed but metabolically active | L. major & L. infantum chagasi | S.C | -- | Treating them with amotosalen and low dose of UV radiation | Shown promise in murine model but failed in human trials | [18] |
| Killed but metabolically active | L. major (Alum ppt. Autoclaved L. major) | -- | BCG | Shown promise in VL and PKDL patients | Chances of relapse | [19] |
| 2ND GENERATION VACCINES | ||||||
| FML (Fucose Mannose Ligand) | L. donovani | S.C, I.P | Saponin, Al(OH)3, QuilA IL-12, BCG | Protective efficacy in BALB/C, Swiss albino, Hamster & dogs | Not shown promise in human | [20,21] |
| Leishmune | L. chagasi | S.C | -- | No parasites were detected in blood, Skin and Lymph node | No trials in humans | [22] |
| Leishmune® | L. donovani L. chagasi | S.C | Saponin | Have transmission blocking property | Partial protection | [23] |
| LAG (L. donovani promastigote membrane antigens) | L. donovani | I.P | Liposome | Shown promise in mice and hamster | Failed in higher animal models | [24] |
| SLA (Soluble Leishmanial Antigen) | L. donovani | I.P, S.C | Liposome, MPL-TDM, non-coding pDNA+ISS | Prophylactic and therapeutic effect in mice model | Free or negatively charged liposome confers partial protection | [25,26,27] |
| gp36 | L. donovani | S.C | Saponin | Protective effect in murine model | Not reached human trials | [28] |
| Ld91, Ld72, Ld51, Ld31 | L. donovani | I.P | Liposome | Reduce parasite burden in visceral organ | Protective efficacy only in mice models | [29] |
| dp72 and gp70-2 | L. donovani | I.P | Corynebacterium parvum | Decrease in parasitemia in visceral organ | Gp70-2 were not promising in mice | [30] |
| SA (Soluble Antigen) | L. donovani | I.M | CpG-ODN | Decrease in parasite load | Ag alone show mixed response in mice models | [31] |
| Ribosomal P0 protein | L. major | S.C | CpG-ODN | Shown promise in C57BL/6 mice | Failure in BALB/C mice | [32] |
| F2 subfraction (97.4-68 kda) | L. donovani | I.D | BCG | Th1 mediated cellular response in cured VL patients and hamsters | Not tested in clinical studies | [33] |
| F2 sub-fraction (89.9-97.1 kDa) | L. donovani | I.D | BCG | 99% parasite inhibition were observed in hamsters | Specific characterization of antigen is needed for higher trials | [34,35] |
| LPG and Phlebotomus duboscqi salivary gland lysates (SGLs) | L. major | S.C | -- | LPG alone provide protection in BALB/C mice | LPG+SGLs failed to provide protection | [36] |
| LPG | Phlebotomus duboscqi sandflies | -- | -- | Block the transmission of Leishmania | -- | [37] |
| gp63 (leishmanolysin) | L. donovani | I.P | Cationic (DSPC) liposomes | Protective effect in BALB/C mice | Mixed responses | [38] |
| Recombinant.gp63 | L. major | -- | -- | Recognition of fusion protein | Fusion protein did not protect the mice | [39] |
| gp63 | L. major | I.V, S.C | -- | BCG expressing gp63 only provide Protection | Limited up to mice only | [40] |
| gp63 | L. major | S.C | CpG ODN + Liposome | Th1 type response | Adjuvant is needed for protection | [41] |
| gp63 | L. donovani | I.M, S.C | CpG ODN | Evoke cellular and humoral response | No trials on higher animal | [42] |
| H2B (Histone protein) | L. major | S.C | CpG | Th1 type response | Only amino-terminal region confer protection | [43] |
| H2B | L. major, L. infantum patients | -- | -- | Whole H2B protein induces Th1 profile in individuals | Not reached clinical trials | [44] |
| rORFF + BT1 | L. donovani | S.C | CFA, ICFA | Detection of high levels of anti-ORFF and anti-BT1 antibodies | Partial protection | [45] |
| rORFF | L. donovani | I.M | CpG ODN | Combination of CpG ODN with rORFF induces reduction in parasite load | Partial protection | [46] |
| ORFF | L. donovani (antimony sensitive & resistant) | I.M | -- | UBQ-ORFF induces cellular & humoral response | Ubiquitin is required for protective effect | [47] |
| Elongation factor 2 (LelF-2) | L. donovani | I.D | -- | Shown promise in Hamster | Not reach clinical trials | [48] |
| rF14 | L. donovani | I.M | MPL | Reduce in parasite burden in spleen and liver | Partial protection | [49] |
| HASPB1 | L. donovani | S.C | IL-12 | Induces DC to produce IL-12 | rHASPB1 immunization induces Th2 response | [50] |
| 78kDa | L. donovani | S.C | MPL-A, Liposome rIL-12, ALD, FCA | Enhanced DTH response, reduction in infection rate of peritoneal macrophages | Antigen alone induces IgG1 isotypes | [51] |
| LCR1 | L. chagasi | I.P, S.C | -- | Protective effect in BALB/C mice | BCG-LCR1 I.P administration did not show protection | [52] |
| Meta1 antigen | L. major | I.M, S.C | IFA | MCP-3/meta 1 fusion partially shift towards Th1 type | Rec. protein Meta1 antigen induces Th2 response | [53] |
| LirCyP1 | L. infantum | I.P | IL-12 | Promote differentiation of helper and memory T cell | Partial protection | [54] |
| Maxadilan (MAX) | L. major | S.C, I.P | CFA IFA | Vaccination against Max elicits Th1 type response | MAX enhance blood flow and inhibit the immune response of the host | [55] |
| LJM19 | L. infantum chagasi | I.D | -- | Induces low parasite load and high IFN-γ/TGF-β ratio | Not reached clinical trials | [56] |
| A2 protein | L. donovani | I.P | Propianibacterium acnes | Production of IFN-ϒ response | Mixed Responses | [57] |
| rLdγGCS | L. donovani | S.C | NIV | Induces humoral response | Mixed Responses | [58] |
| FUSION, HYBRID AND POLYPROTEIN VACCINES | ||||||
| Chimeric Q protein | L. infantum | S.C | -- | Clearance of parasites from majority of the dogs | Studies during natural infection condition are required | [59] |
| Amastigote cysteine proteases | L. major | S.C | -- | Hybrid CPA/B elicits protective response | Partial protection | [60] |
| TAT-antigen fusion protein | L. major | I.D | CpG ODN 1826 | CD8+ T cells rapidly proliferate, activate Leishmania-specific Tc1 cells | Protective effect is observed in C57BL/6 mice | [61] |
| rLeish-111f | L. major | S.C | rIL-12 MPL-SE | Protective efficacy in BALB/C mice | Mixed IgG2a/IgG1 Ab response | [62] |
| Leish-111f | L. infantum | S.C | MPL-SE | 99.6% reductions in parasite loads, completed phase 1 and 2 safety human trial | -- | [63] |
| LEISH-F1 | L. donovani | S.C | MPL-SE | Vaccine is safe, immunogenic in healthy subjects | Further trials are needed | [64] |
| LIVE, ATTENUATED (GENETICALLY) VACCINES | ||||||
| dhfr-ts− null mutant | L. major | I.V, S.C, I.M | -- | Unable to cause disease in susceptible and immune deficient (nu/nu) BALB/C mice | Need to be tested on animal higher models | [65] |
| dhfr-ts− L. major knock out mutant | L. amazonensis | S.C, I.V | -- | Cross- Protection was observed in both BALB/C and C57BL/6 mice | Partial degree of protection | [66] |
| LdCen1(−/−) L. donovani centrin null mutants | L. donovani L. braziliensis | I.M I.C | -- | No parasites were seen in spleen, liver in BALB/C, SCID mice and Hamster, Cross protection against L. braziliensis | Not reach clinical trials | [67] |
| (BT1−/−) Biopterin transporter null mutant | L. donovani | I.V | -- | Th1 mediated response were observed in BALB/C mice | Need to be tested on higher animal models | [68] |
| LiSIR2(+/−) L. infantum SIR2 single knockout | L. infantum | I.P | -- | Reversion of T cell anergy, increased IFN-γ/IL-10 ratio | Mixed response in BALB/C mice | [69] |
| Ldp27(−/−) | L. donovani L. infantum chagasi | I.C | -- | less COX (Cytochrome C oxidase complex) activity and ATP synthesis | Safety efficacy need to be tested on higher animal models | [70] |
| ΔHSP70-II | L. major | I.P, I.V, S.C | -- | Promise protection in BALB/C model, consider safe in SCID mice and hamster | Need to be tested on higher animal models | [71] |
| LiΔHSP70-II null mutant | L. major | I.V, S.C | -- | Promise protection in BALB/C and C57BL/6 mice | Limited only to mice models | [72] |
| Δlpg2 | L. major | S.C | CPG-ODN | Shown promise in C57BL/6 mice | Antigen alone is not sufficient to provide protection | [73] |
| lpg2− | L. major | Foot pad | -- | Suppress IL-4 and IL-10 responses | Very low IFN-ϒ response were observed | [74] |
| Leishmania tarentolae (Non-pathogenic) | L. donovani | I.P | -- | Activates DC, Induces IFN-ϒ, Th1 phenotype in BALB/C mice | Further testing is needed | [75] |
| A2-recombinant L. tarentolae | L. infantum | I.P, I.V | -- | Th1 response in BALB/C mice | I.V route elicits Th2 response | [76] |
| A2-expressing Lactococcus lactis | L. donovani | S.C | -- | Antigen-specific humoral response was observed | Only in BALB/C mice | [77] |
| 3RD GENERATION VACCINES | ||||||
| LACK | L. major | S.C | +rIL-12 -rIL-12 | Protective response in BALB/C mice | Need to be tested on higher animal models | [78] |
| pCIneo-LACK | L. chagasi | I.N Intranasal | -- | Reduce parasite burden in liver and spleen | Limited up to BALB/C mice | [79] |
| LACK-DNA | L. chagasi | I.N | -- | Cellular and humoral response were observed | Not reached clinical trials | [80] |
| p36 (LACK) DNA | L. donovani | I.D, S.C | IL-12 | Robust Th1 response, highly immunogenic | No protection | [81] |
| gp63 and Hsp70 | L. donovani | S.C | -- | Significantly reduce the parasite burden | Need to be tested on higher animal models | [82] |
| A2+E6 | L. donovani | I.M | -- | Induce cellular and humoral response, inhibit cellular p53 response | Limited to mice model | [83] |
| A2 (Expressed in adenovirus) | L. chagasi | S.C | -- | Reduced parasitism in spleen and liver, production of high level of IFN-ϒ | Low antibody response | [84] |
| ORFF | L. donovani | I.M | -- | Induces both cellular and humoral response | Limited to BALB/C mice | [85] |
| rORFF | L. donovani | I.M | IL-12 | Higher levels of IFN-γ were observed | Need to be tested on higher animal models | [86] |
| KMP-11 | L. donovani | I.M | -- | Reversal of T-cell anergy with functional IL-2 generation | Mixed response | [87] |
| KMP-11 | L. major | I.M, S.C | IL-12 | Robust IFN-ϒ production, polarized Th1 response | Require adjuvant for complete protection | [88] |
| ϒ GCS | L. donovani | I.M | -- | Elevated humoral and cell mediated response | Limited to mice model only | [89] |
| H2A, H2B, H3, H4, LACK | L. donovani | I.M, I.D | IL-12 GM-CSF | Induce DTH response, reduction in parasite burden | Unable to induce parasite-specific antibody response | [90] |
| N-terminal domain of proteophosphoglycan (PPG) | L. donovani | I.M | -- | Induce IFN-ϒ, TNF-α, IL-12 and downregulate TGF-β, IL-4, IL-10 | Need further trials ahead of hamster model | [91] |
| Single antigen Gp63, polytope and polytope HSP70 | L. donovani | I.M, I.P | -- | Strong Th1 response | Limited to mice only | [92] |
| NH36 | L. chagasi L. mexicana | I.M, S.C | -- | Reduction in parasite load, Th1 mediated response, cross-protection | No differences were observed after infection with L. mexicana | [93] |
| NH36 | L. chagasi | I.M, I.P | -- | Reduction in parasite burden in liver | Clinical trials not done | [94] |
| KMP11, TSA, elongation factor P74, CPA and CPB, HASPB, A2 LEISHDNAVAX | L. donovani | I.D | -- | Induce CD4+ and CD8+ T cell responses in genetically diverse human populations | Require clinical trials | [95] |
| KMPII, TRYP, LACK and GP63 | L. infantum | I.D | -- | Vaccine was safe and well tolerated for dogs | No protection in dogs | [96] |
| papLe22 | L. infantum | I.M | -- | Downregulate Th2 type response in hamsters | Low antibody response | [97] |
| HbR | L. donovani | I.M | -- | Complete protection in both BALB/C and Hamsters | Clinical trials not done | [98] |
| Tuzin | L. donovani | I.M | -- | higher levels of IFN-γ and IL-12 production | Limited only to BALB/C mice | [99] |
| HETEROLOGOUS PRIME-BOOST (HPB) VACCINES | ||||||
| ORFF | L. donovani | I.M | Alum | 75%–80% reduction in parasite load, enhanced production of IgG2a and IFN-γ in BALB/C mice | Need to be tested on higher animal models | [100] |
| Cysteine proteinases type I and II | L. infantum | S.C | CpG ODN and Montanide 720 | Antigen-specific immune response was observed | Need to be tested on higher animal model | [101] |
| rLdP1 | L. donovani | I.M | FCA | 75.68% decrease in splenic parasite burden in hamsters, | Challenge by other Leishmania species needed for evaluation | [102] |
| A2-CPA-CPB (CTE) recombinant L. tarentolae | L. infantum | Foot pad | cationic solid lipid nanoparticle (cSLN) | High NO production, and low parasite burden | Mixed responses | [103] |
| LACK-WR or LACK-MVA | L. infantum | I.D, I.P | -- | High levels of protection in the draining lymph node | Low protection observed in Spleen and Liver | [104] |
| AlphaGalCer + DNAp36+ VVp36 | L. major | I.D, I.P | αGalCer | 20-fold reductions in parasite burdens | Limited to mice only | [105] |
| DNA/MVA TRYP | L. Viannia panamensis | I.D | α-GalCer, LPS, CpG, Pam3CSK4, MALP-2 | TLR1/2 activation, antigen specific CD8 cells | Need to be tested on higher animal model | [106] |
| pORT-LACK/MVA-LACK | L. infantum | S.C | -- | increase in expression of Th1 type cytokines in PBMC and target organs | Limited to canine model only | [107] |
| LdPxn-1 (L. donovani Peroxidoxin-1) | L. major | I.M S.C | m-GMCSF | Induction of multipotent CD4+ cells | Mixed responses | [108] |
| DENDRITIC CELL(DC)-BASED VACCINES | ||||||
| DCs pulsed with peptide L1 of gp63 (SPL) & pulsing with peptide L2 of gp63 | L. major | I.V | -- | Reduce lesion and parasite load, Th1 phenotype in BALB/C mice with L1 peptide | Partial protection with L1 peptide, whereas L2 peptide switch towards Th2 profile | [109] |
| Plasmacytoid DCs | L. major | I.V | CpG-ODN | Induce T cell-mediated protection | IFN-α production could not be detected | [110] |
| SLDA-pulsed DCs | L. donovani | I.V | IL-12 | Live parasites were not detected in the liver of mice, 1-3 log lower parasite burdens | Granuloma formation were found in SLDA-pulsed, non-transduced DCs BALB/C mice | [111] |
| BM-DCs pulsed with KMP-11 (12–31aa) peptide | L. infantum | I.V | CpG-ODN | Induced the differentiation of peptide-specific Th17 cells | Mixed responses | [112] |
| SLDA-pulsed syngeneic BM-DCs | L. donovani | I.V, I.M | -- | Complete clearance of parasites from spleen and liver | Need to be tested on higher animal models | [113] |
| Hybrid cell vaccine (HCV) | L. donovani | I.V | -- | Strong antigen-specific CTL generation | Expression of IL-4 and IL-13 get elevated at both transcription and translational levels | [114] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zutshi, S.; Kumar, S.; Chauhan, P.; Bansode, Y.; Nair, A.; Roy, S.; Sarkar, A.; Saha, B. Anti-Leishmanial Vaccines: Assumptions, Approaches, and Annulments. Vaccines 2019, 7, 156. https://doi.org/10.3390/vaccines7040156
Zutshi S, Kumar S, Chauhan P, Bansode Y, Nair A, Roy S, Sarkar A, Saha B. Anti-Leishmanial Vaccines: Assumptions, Approaches, and Annulments. Vaccines. 2019; 7(4):156. https://doi.org/10.3390/vaccines7040156
Chicago/Turabian StyleZutshi, Shubhranshu, Sunil Kumar, Prashant Chauhan, Yashwant Bansode, Arathi Nair, Somenath Roy, Arup Sarkar, and Bhaskar Saha. 2019. "Anti-Leishmanial Vaccines: Assumptions, Approaches, and Annulments" Vaccines 7, no. 4: 156. https://doi.org/10.3390/vaccines7040156
APA StyleZutshi, S., Kumar, S., Chauhan, P., Bansode, Y., Nair, A., Roy, S., Sarkar, A., & Saha, B. (2019). Anti-Leishmanial Vaccines: Assumptions, Approaches, and Annulments. Vaccines, 7(4), 156. https://doi.org/10.3390/vaccines7040156