Immunogenicity of HIV-1 Vaccines Expressing Chimeric Envelope Glycoproteins on the Surface of Pr55 Gag Virus-Like Particles

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Antibodies, Plasmids, Cell Lines, Media, and Reagents
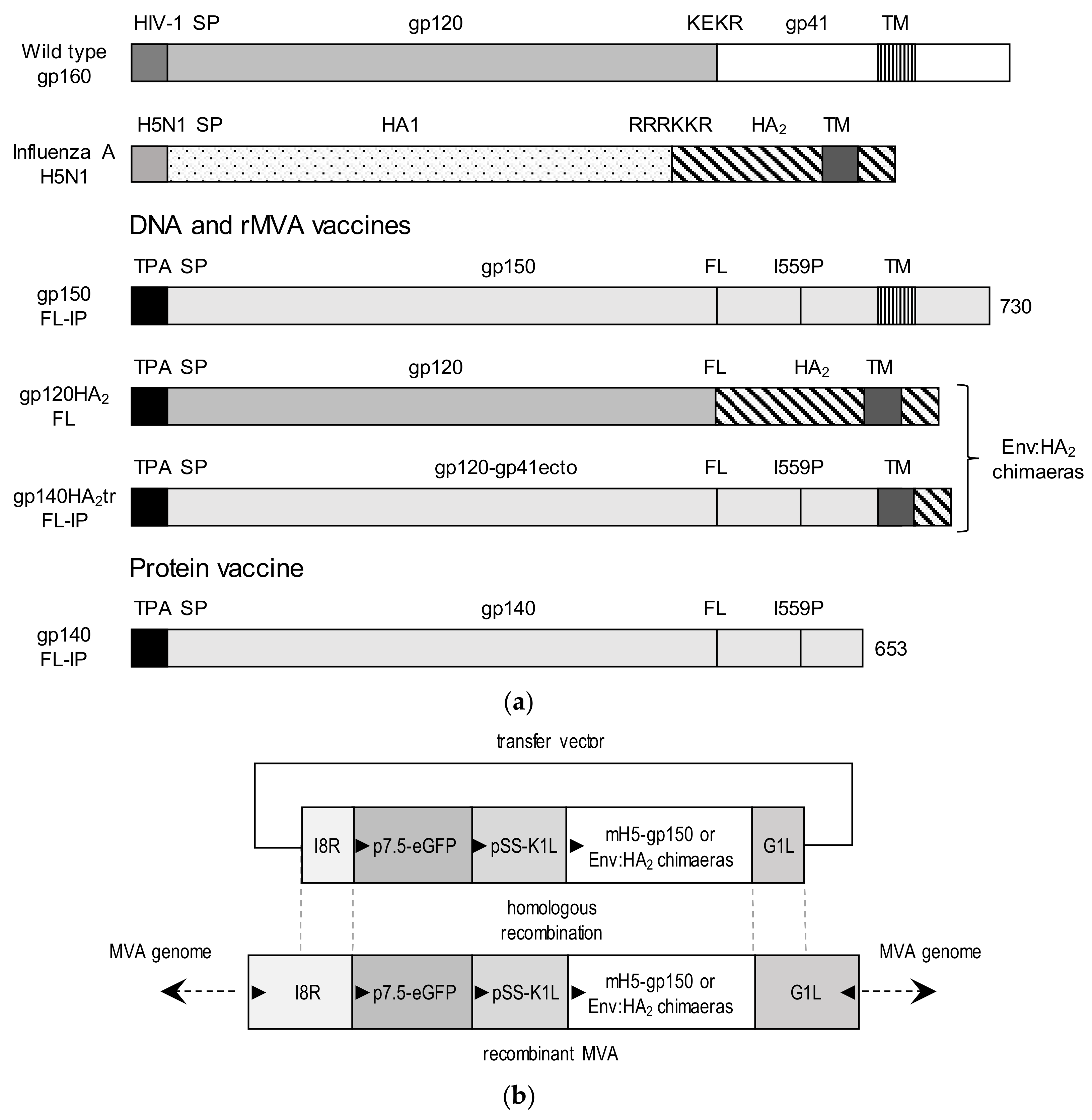
2.2. Design and Construction of DNA and MVA Vaccines Expressing Chimeric Env
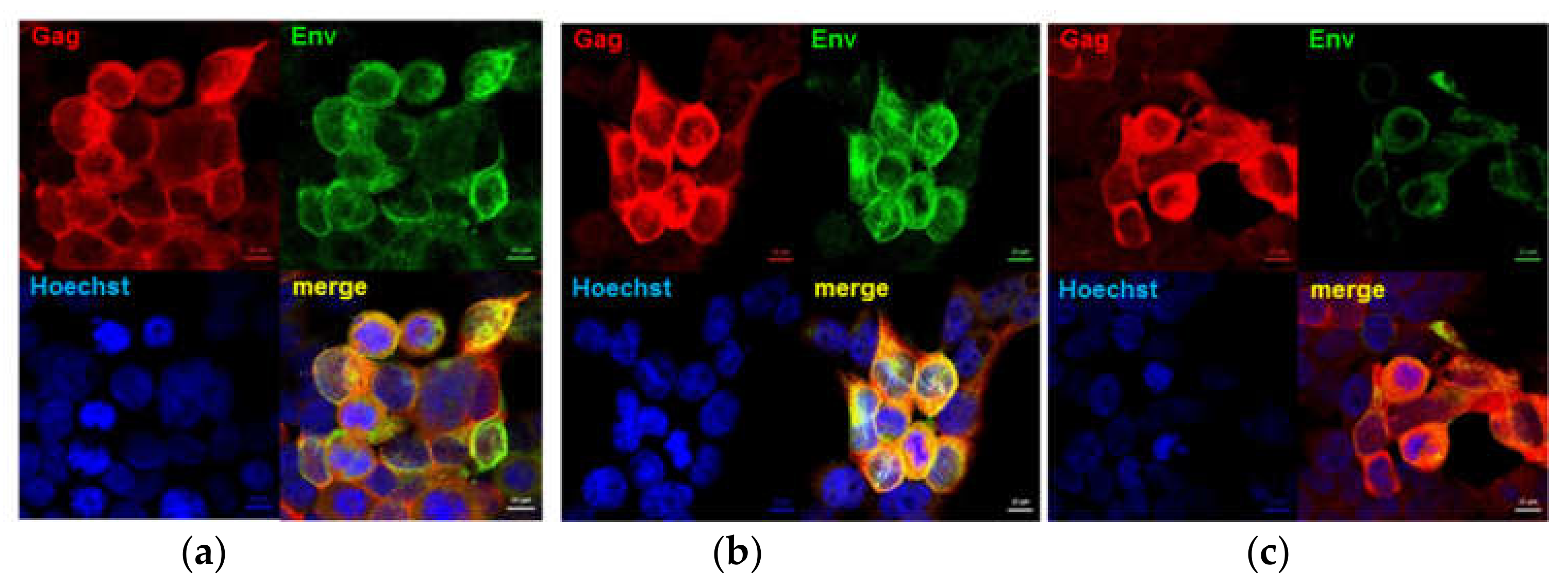
2.3. Confirmation of Gag and Chimeric Env Expression by DNA and MVA Vaccines
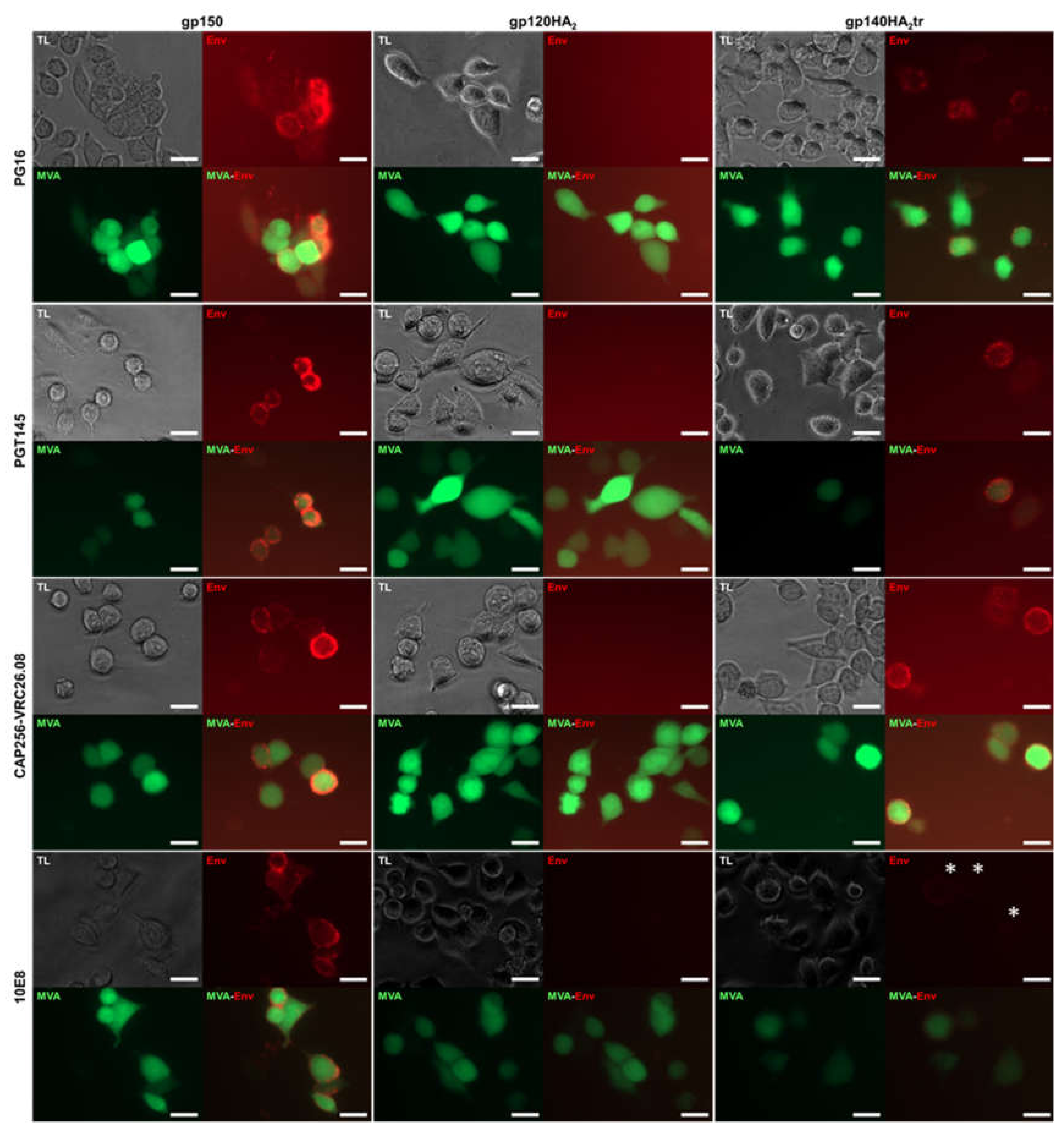
2.4. Live Cell Staining with Human, Monoclonal Antibodies to HIV-1 Env
2.5. VLP Isolation and Characterization
2.6. Rabbit Immunizations
2.7. Anti-CAP256 SU gp140 and Scaffolded-CAP256 SU V1V2-loop Enzyme-Linked Immunosorbent Assay (ELISA)
2.8. HIV Neutralization Assays
2.9. Statistical Analysis
3. Results
3.1. Design of Chimeric Env Proteins
3.2. Expression of Chimeric HIV Envelope Proteins
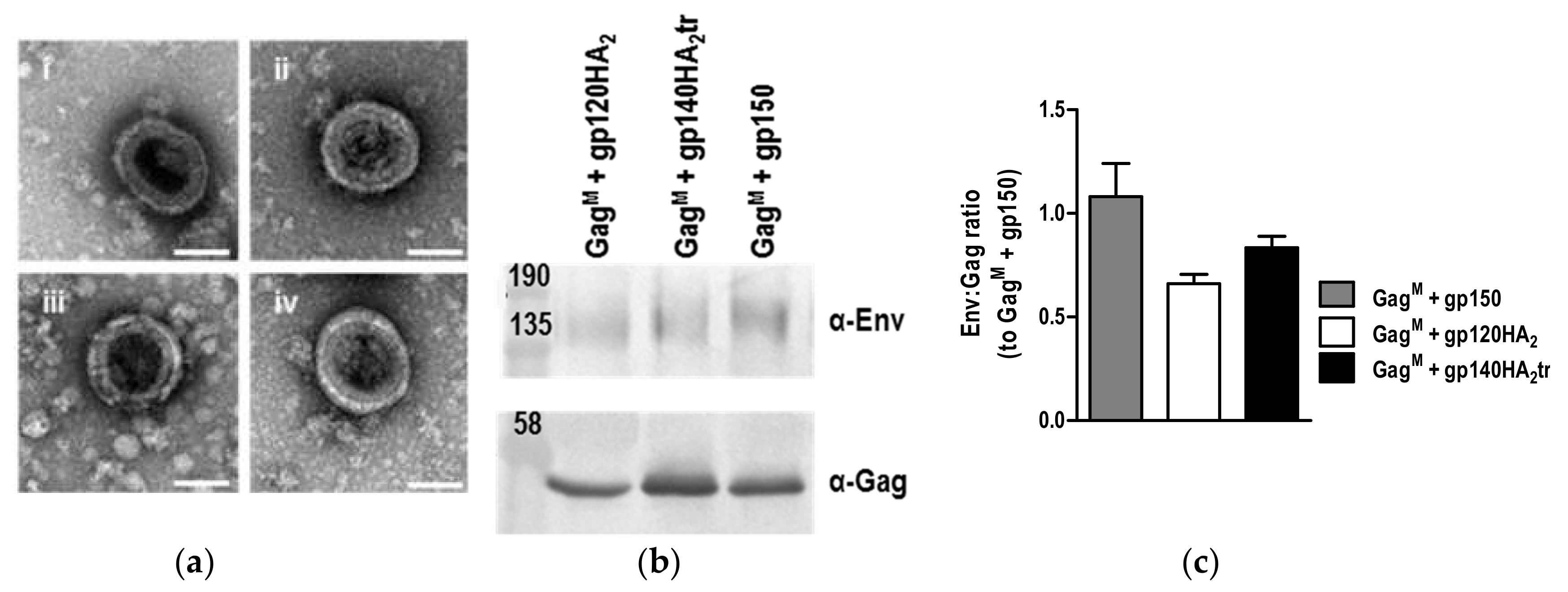
3.3. Characterization of VLPs
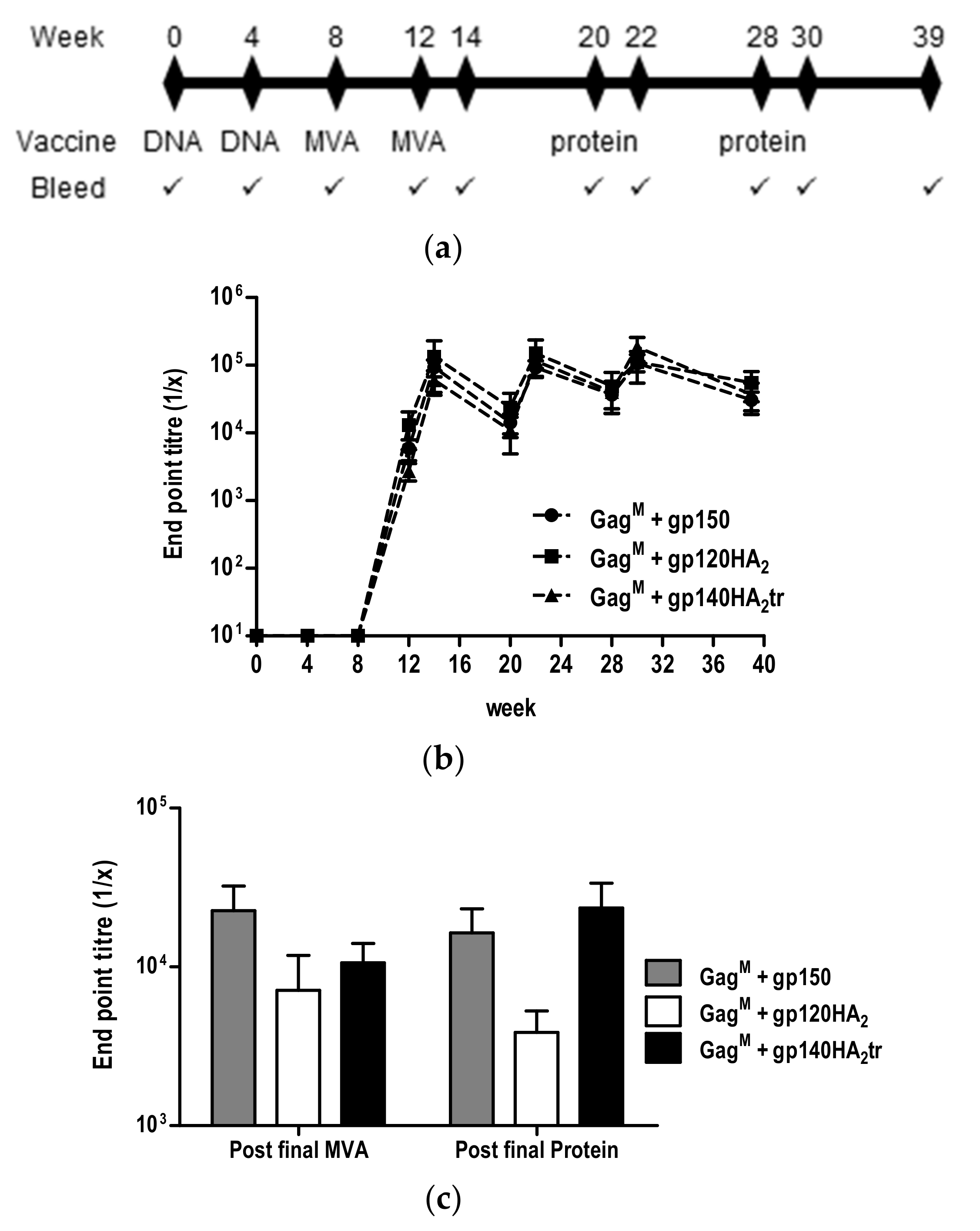
3.4. Immunogenicity Testing
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- UNAIDS. Global HIV & AIDs Stastics - 2019 Fact Sheet. Available online: https://www.unaids.org/en/resources/fact-sheet (accessed on 26 December 2019).
- Alter, G.; Barouch, D. Immune Correlate-Guided HIV Vaccine Design. Cell Host Microbe 2018, 24, 25–33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schiller, J.; Chackerian, B. Why HIV virions have low numbers of envelope spikes: implications for vaccine development. PLoS Pathog. 2014, 10, e1004254. [Google Scholar] [CrossRef] [PubMed]
- Bachmann, M.F.; Zinkernagel, R.M. The influence of virus structure on antibody responses and virus serotype formation. Immunol. Today 1996, 17, 553–558. [Google Scholar] [CrossRef]
- Chackerian, B. Virus-like particles: flexible platforms for vaccine development. Expert Rev. Vaccines 2007, 6, 381–390. [Google Scholar] [CrossRef] [PubMed]
- Ingale, J.; Stano, A.; Guenaga, J.; Sharma, S.K.; Nemazee, D.; Zwick, M.B.; Wyatt, R.T. High-Density Array of Well-Ordered HIV-1 Spikes on Synthetic Liposomal Nanoparticles Efficiently Activate B Cells. Cell Rep. 2016, 15, 1986–1999. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuste, E.; Reeves, J.D.; Doms, R.W.; Desrosiers, R.C. Modulation of Env content in virions of simian immunodeficiency virus: correlation with cell surface expression and virion infectivity. J. Virol. 2004, 78, 6775–6785. [Google Scholar] [CrossRef] [Green Version]
- Deml, L.; Schirmbeck, R.; Reimann, J.; Wolf, H.; Wagner, R. Recombinant human immunodeficiency Pr55gag virus-like particles presenting chimeric envelope glycoproteins induce cytotoxic T-cells and neutralizing antibodies. Virology 1997, 235, 26–39. [Google Scholar] [CrossRef] [Green Version]
- Vzorov, A.N.; Wang, L.; Chen, J.; Wang, B.Z.; Compans, R.W. Effects of modification of the HIV-1 Env cytoplasmic tail on immunogenicity of VLP vaccines. Virology 2016, 489, 141–150. [Google Scholar] [CrossRef] [Green Version]
- Wang, B.Z.; Liu, W.; Kang, S.M.; Alam, M.; Huang, C.; Ye, L.; Sun, Y.; Li, Y.; Kothe, D.L.; Pushko, P.; et al. Incorporation of high levels of chimeric human immunodeficiency virus envelope glycoproteins into virus-like particles. J. Virol. 2007, 81, 10869–10878. [Google Scholar] [CrossRef] [Green Version]
- Visciano, M.L.; Diomede, L.; Tagliamonte, M.; Tornesello, M.L.; Asti, V.; Bomsel, M.; Buonaguro, F.M.; Lopalco, L.; Buonaguro, L. Generation of HIV-1 Virus-Like Particles expressing different HIV-1 glycoproteins. Vaccine 2011, 29, 4903–4912. [Google Scholar] [CrossRef]
- Bresk, C.A.; Hofer, T.; Wilmschen, S.; Krismer, M.; Beierfuss, A.; Effantin, G.; Weissenhorn, W.; Hogan, M.J.; Jordan, A.P.O.; Gelman, R.S.; et al. Induction of Tier 1 HIV Neutralizing Antibodies by Envelope Trimers Incorporated into a Replication Competent Vesicular Stomatitis Virus Vector. Viruses 2019, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Diepen, M.T.; Chapman, R.; Douglass, N.; Galant, S.; Moore, P.L.; Margolin, E.; Ximba, P.; Morris, L.; Rybicki, E.P.; Williamson, A.L. Prime-Boost Immunizations with DNA, Modified Vaccinia Virus Ankara, and Protein-Based Vaccines Elicit Robust HIV-1 Tier 2 Neutralizing Antibodies against the CAP256 Superinfecting Virus. J. Virol. 2019, 93. [Google Scholar] [CrossRef] [Green Version]
- Bhiman, J.N.; Anthony, C.; Doria-Rose, N.A.; Karimanzira, O.; Schramm, C.A.; Khoza, T.; Kitchin, D.; Botha, G.; Gorman, J.; Garrett, N.J.; et al. Viral variants that initiate and drive maturation of V1V2-directed HIV-1 broadly neutralizing antibodies. Nat. Med. 2015, 21, 1332–1336. [Google Scholar] [CrossRef] [PubMed]
- Gorman, J.; Soto, C.; Yang, M.M.; Davenport, T.M.; Guttman, M.; Bailer, R.T.; Chambers, M.; Chuang, G.Y.; DeKosky, B.J.; Doria-Rose, N.A.; et al. Structures of HIV-1 Env V1V2 with broadly neutralizing antibodies reveal commonalities that enable vaccine design. Nat. Struct. Mol. Biol. 2016, 23, 81–90. [Google Scholar] [CrossRef] [PubMed]
- Moore, P.L.; Sheward, D.; Nonyane, M.; Ranchobe, N.; Hermanus, T.; Gray, E.S.; Abdool Karim, S.S.; Williamson, C.; Morris, L. Multiple pathways of escape from HIV broadly cross-neutralizing V2-dependent antibodies. J. Virol. 2013, 87, 4882–4894. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanzer, F.L.; Shephard, E.G.; Palmer, K.E.; Burger, M.; Williamson, A.L.; Rybicki, E.P. The porcine circovirus type 1 capsid gene promoter improves antigen expression and immunogenicity in a HIV-1 plasmid vaccine. Virology journal 2011, 8, 51. [Google Scholar] [CrossRef] [Green Version]
- Chapman, R.; Jongwe, T.I.; Douglass, N.; Chege, G.; Williamson, A.L. Heterologous prime-boost vaccination with DNA and MVA vaccines, expressing HIV-1 subtype C mosaic Gag virus-like particles, is highly immunogenic in mice. PLoS ONE 2017, 12, e0173352. [Google Scholar] [CrossRef]
- Jongwe, T.I.; Chapman, R.; Douglass, N.; Chetty, S.; Chege, G.; Williamson, A.L. HIV-1 Subtype C Mosaic Gag Expressed by BCG and MVA Elicits Persistent Effector T Cell Responses in a Prime-Boost Regimen in Mice. PLoS ONE 2016, 11, e0159141. [Google Scholar] [CrossRef]
- Samal, S.; Bansal, M.; Das, S. Method to identify efficiently cleaved, membrane-bound, functional HIV-1 (Human Immunodeficiency Virus-1) envelopes. MethodsX 2019, 6, 837–849. [Google Scholar] [CrossRef]
- McLellan, J.S.; Pancera, M.; Carrico, C.; Gorman, J.; Julien, J.P.; Khayat, R.; Louder, R.; Pejchal, R.; Sastry, M.; Dai, K.; et al. Structure of HIV-1 gp120 V1/V2 domain with broadly neutralizing antibody PG9. Nature 2011, 480, 336–343. [Google Scholar] [CrossRef]
- Van Diepen, M.T.; Chapman, R.; Moore, P.L.; Margolin, E.; Hermanus, T.; Morris, L.; Ximba, P.; Rybicki, E.P.; Williamson, A.L. The adjuvant AlhydroGel elicits higher antibody titres than AddaVax when combined with HIV-1 subtype C gp140 from CAP256. PLoS ONE 2018, 13, e0208310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, S.K.; de Val, N.; Bale, S.; Guenaga, J.; Tran, K.; Feng, Y.; Dubrovskaya, V.; Ward, A.B.; Wyatt, R.T. Cleavage-independent HIV-1 Env trimers engineered as soluble native spike mimetics for vaccine design. Cell Rep. 2015, 11, 539–550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanders, R.W.; Vesanen, M.; Schuelke, N.; Master, A.; Schiffner, L.; Kalyanaraman, R.; Paluch, M.; Berkhout, B.; Maddon, P.J.; Olson, W.C.; et al. Stabilization of the soluble, cleaved, trimeric form of the envelope glycoprotein complex of human immunodeficiency virus type 1. J. Virol. 2002, 76, 8875–8889. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pancera, M.; McLellan, J.S.; Wu, X.; Zhu, J.; Changela, A.; Schmidt, S.D.; Yang, Y.; Zhou, T.; Phogat, S.; Mascola, J.R.; et al. Crystal structure of PG16 and chimeric dissection with somatically related PG9: structure-function analysis of two quaternary-specific antibodies that effectively neutralize HIV-1. J. Virol. 2010, 84, 8098–8110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, X.; Changela, A.; O’Dell, S.; Schmidt, S.D.; Pancera, M.; Yang, Y.; Zhang, B.; Gorny, M.K.; Phogat, S.; Robinson, J.E.; et al. Immunotypes of a quaternary site of HIV-1 vulnerability and their recognition by antibodies. J. Virol. 2011, 85, 4578–4585. [Google Scholar] [CrossRef] [Green Version]
- Sanders, R.W.; Derking, R.; Cupo, A.; Julien, J.P.; Yasmeen, A.; de Val, N.; Kim, H.J.; Blattner, C.; de la Pena, A.T.; Korzun, J.; et al. A next-generation cleaved, soluble HIV-1 Env trimer, BG505 SOSIP.664 gp140, expresses multiple epitopes for broadly neutralizing but not non-neutralizing antibodies. PLoS Pathog. 2013, 9, e1003618. [Google Scholar] [CrossRef] [Green Version]
- Andrabi, R.; Voss, J.E.; Liang, C.H.; Briney, B.; McCoy, L.E.; Wu, C.Y.; Wong, C.H.; Poignard, P.; Burton, D.R. Identification of Common Features in Prototype Broadly Neutralizing Antibodies to HIV Envelope V2 Apex to Facilitate Vaccine Design. Immunity 2015, 43, 959–973. [Google Scholar] [CrossRef]
- Pancera, M.; Shahzad-Ul-Hussan, S.; Doria-Rose, N.A.; McLellan, J.S.; Bailer, R.T.; Dai, K.; Loesgen, S.; Louder, M.K.; Staupe, R.P.; Yang, Y.; et al. Structural basis for diverse N-glycan recognition by HIV-1-neutralizing V1-V2-directed antibody PG16. Nat. Struct. Mol. Biol. 2013, 20, 804–813. [Google Scholar] [CrossRef] [Green Version]
- Doria-Rose, N.A.; Schramm, C.A.; Gorman, J.; Moore, P.L.; Bhiman, J.N.; DeKosky, B.J.; Ernandes, M.J.; Georgiev, I.S.; Kim, H.J.; Pancera, M.; et al. Developmental pathway for potent V1V2-directed HIV-neutralizing antibodies. Nature 2014, 509, 55–62. [Google Scholar] [CrossRef]
- Moore, P.L.; Gray, E.S.; Sheward, D.; Madiga, M.; Ranchobe, N.; Lai, Z.; Honnen, W.J.; Nonyane, M.; Tumba, N.; Hermanus, T.; et al. Potent and broad neutralization of HIV-1 subtype C by plasma antibodies targeting a quaternary epitope including residues in the V2 loop. J. Virol. 2011, 85, 3128–3141. [Google Scholar] [CrossRef] [Green Version]
- Stano, A.; Leaman, D.P.; Kim, A.S.; Zhang, L.; Autin, L.; Ingale, J.; Gift, S.K.; Truong, J.; Wyatt, R.T.; Olson, A.J.; et al. Dense Array of Spikes on HIV-1 Virion Particles. J. Virol. 2017, 91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.Y.; Song, W.T.; Li, Y.; Chen, W.J.; Yang, D.; Zhong, G.C.; Zhou, H.Z.; Ren, C.Y.; Yu, H.T.; Ling, H. Improved expression of secretory and trimeric proteins in mammalian cells via the introduction of a new trimer motif and a mutant of the tPA signal sequence. Appl. Microbiol. Biotechnol. 2011, 91, 731–740. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Farfan-Arribas, D.J.; Shen, S.; Chou, T.H.; Hirsch, A.; He, F.; Lu, S. Relative contributions of codon usage, promoter efficiency and leader sequence to the antigen expression and immunogenicity of HIV-1 Env DNA vaccine. Vaccine 2006, 24, 4531–4540. [Google Scholar] [CrossRef] [PubMed]
- Bale, S.; Martine, A.; Wilson, R.; Behrens, A.J.; Le Fourn, V.; de Val, N.; Sharma, S.K.; Tran, K.; Torres, J.L.; Girod, P.A.; et al. Cleavage-Independent HIV-1 Trimers From CHO Cell Lines Elicit Robust Autologous Tier 2 Neutralizing Antibodies. Front Immunol. 2018, 9, 1116. [Google Scholar] [CrossRef]
- Sanders, R.W.; van Gils, M.J.; Derking, R.; Sok, D.; Ketas, T.J.; Burger, J.A.; Ozorowski, G.; Cupo, A.; Simonich, C.; Goo, L.; et al. HIV-1 VACCINES. HIV-1 neutralizing antibodies induced by native-like envelope trimers. Science 2015, 349, aac4223. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
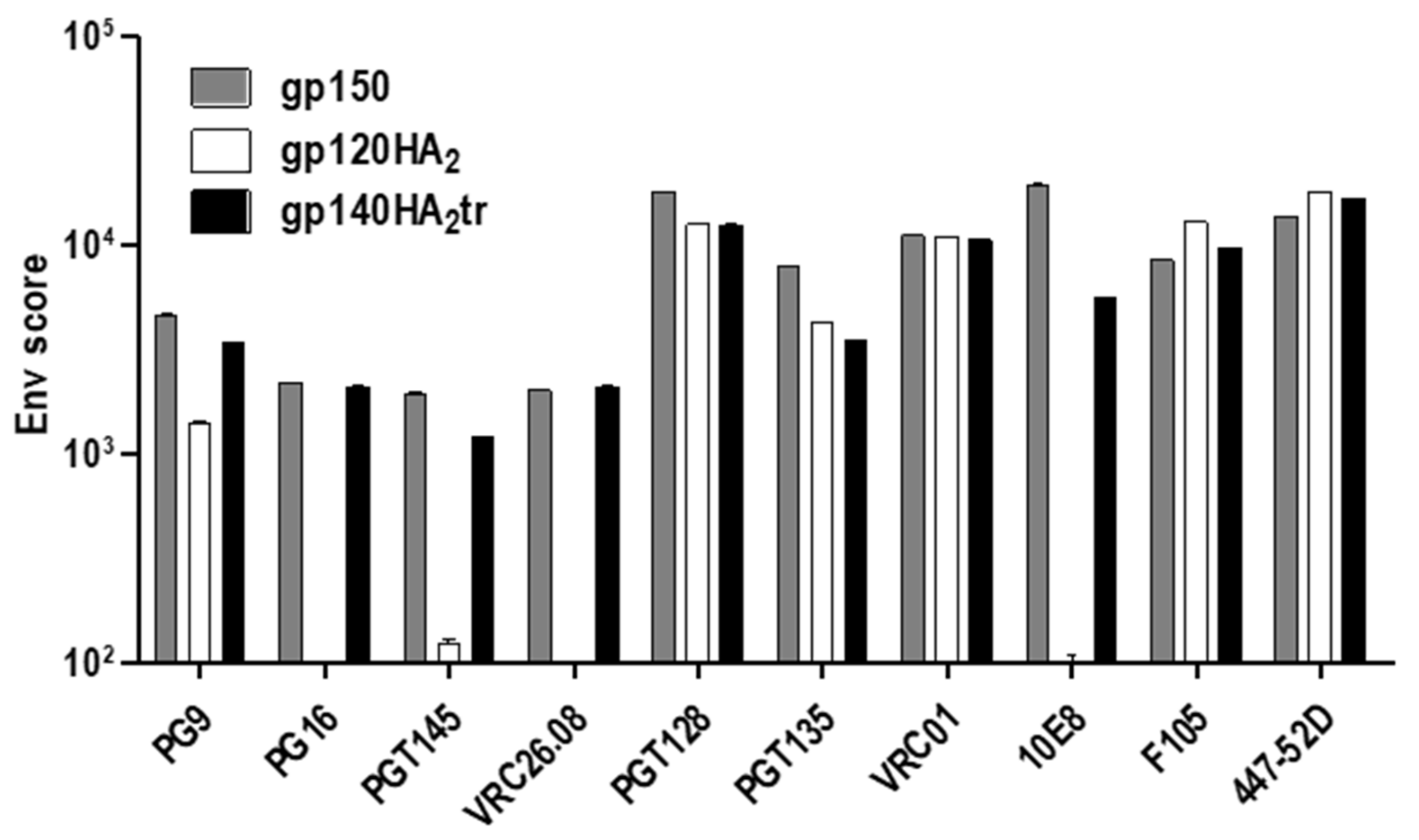{kind=link}
{kind=link}
{kind=link}
| Antibody | Neutralization | Epitope | Native-Like Trimer | Live Cell Mapping * | ||
|---|---|---|---|---|---|---|
| gp150 | gp120HA2 | gp140HA2tr | ||||
| PGT128 | Broad | V3-glycan supersite | x | √ | √ | √ |
| PGT135 | Broad | V3-glycan supersite | x | √ | √ | √ |
| 447-52D | Narrow | V3-glycan | x | √ | √ | √ |
| VRC01 | Broad | CD4 binding-site | x | √ | √ | √ |
| F105 | Narrow | CD4 binding-site | x | √ | √ | √ |
| PG9 | Broad | V2 apex | x | √ | √ | √ |
| PG16 | Broad | V2 apex | Yes | √ | X | √ |
| PGT145 | Broad | V2 apex | Yes | √ | X | √ |
| CAP256-VRC26.08 | Broad | V2 apex | Yes | √ | X | √ |
| 10E8 | Broad | MPER | x | √ | X | Weak |
| Clade C Tier 1A | Clade C Tier 1B | Clade C Tier 1B | Clade C Tier 2 | Control | ||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| MW965.26 | 6644 | 1107356 | CAP256 SU | MLV | ||||||||||||||||||
| ID50 after: | ID50 after: | ID50 after: | ID50 after: | ID50 after: | ||||||||||||||||||
| Regimen | Rabbit | pre | M1 | M2 | P1 | P2 | pre | M1 | M2 | P1 | P2 | pre | M1 | M2 | P1 | P2 | pre | M1 | M2 | P1 | P2 | pre-wk30 |
| DDMMPP gp150 GagM | 6826 | <20 | 195 | 3426 | 5286 | 7840 | <20 | <20 | 85 | 45 | 170 | <20 | <20 | 32 | <20 | 29 | <20 | <20 | 70 | 486 | 1294 | <20 |
| 6827 | <20 | 119 | 3892 | 14601 | 2748 | <20 | <20 | 120 | 196 | 104 | <20 | <20 | 35 | <20 | <20 | <20 | <20 | 53 | 519 | 174 | <20 | |
| 6828 | <20 | 81 | 7239 | 664 | 4348 | <20 | <20 | 172 | 20 | 136 | <20 | <20 | 62 | <20 | 21 | <20 | <20 | <20 | <20 | <20 | <20 | |
| 6830 | <20 | 108 | 12641 | 20778 | 3920 | <20 | <20 | 299 | 172 | 143 | <20 | <20 | 40 | <20 | <20 | <20 | <20 | 60 | 332 | 296 | <20 | |
| 6850 | <20 | <20 | 1999 | 1987 | 787 | <20 | <20 | 78 | 40 | 41 | <20 | <20 | <20 | <20 | <20 | <20 | <20 | <20 | 32 | 54 | <20 | |
| median | <20 | 108 | 3892 | 5286 | 3920 | <20 | <20 | 120 | 45 | 136 | <20 | <20 | 35 | <20 | <20 | <20 | <20 | 53 | 332 | 174 | <20 | |
| DDMMPP gp120HA2 GagM | 6832 | <20 | <20 | 470 | 3594 | 1115 | <20 | <20 | <20 | 30 | <20 | <20 | <20 | 21 | <20 | <20 | <20 | <20 | <20 | <20 | <20 | <20 |
| 6833 | <20 | 166 | 82 | 96 | 211 | <20 | <20 | <20 | <20 | <20 | <20 | <20 | <20 | <20 | <20 | <20 | <20 | <20 | <20 | <20 | <20 | |
| 6834 | <20 | 36 | 42 | 305 | 148 | <20 | <20 | <20 | 21 | <20 | <20 | <20 | <20 | <20 | <20 | <20 | <20 | <20 | <20 | <20 | <20 | |
| 6835 | <20 | 25 | 2410 | 1050 | 3680 | <20 | <20 | 130 | 74 | 83 | <20 | <20 | <20 | <20 | 23 | <20 | <20 | <20 | <20 | <20 | <20 | |
| median | <20 | 31 | 276 | 678 | 663 | <20 | <20 | <20 | 26 | <20 | <20 | <20 | <20 | <20 | <20 | <20 | <20 | <20 | <20 | <20 | <20 | |
| DDMMPP gp140HA2tr GagM | 6836 | <20 | 35 | 8193 | 6134 | 1786 | <20 | <20 | 595 | 175 | 158 | <20 | <20 | 47 | <20 | <20 | <20 | <20 | <20 | <20 | <20 | <20 |
| 6838 | <20 | 97 | 1060 | 6524 | 3235 | <20 | <20 | 113 | 154 | 113 | <20 | <20 | <20 | <20 | 25 | <20 | <20 | <20 | <20 | <20 | <20 | |
| 6839 | <20 | 53 | 578 | 8835 | 2578 | <20 | <20 | 32 | 262 | 89 | <20 | <20 | <20 | 24 | <20 | <20 | <20 | <20 | 242 | 151 | <20 | |
| 6840 | <20 | 44 | 101 | 384 | 298 | <20 | <20 | <20 | <20 | <20 | <20 | <20 | <20 | <20 | <20 | <20 | <20 | <20 | 103 | 44 | <20 | |
| 6842 | <20 | 87 | NT | 5783 | 2816 | <20 | <20 | NT | 112 | 128 | <20 | <20 | NT | <20 | <20 | <20 | <20 | NT | 171 | 220 | <20 | |
| median | <20 | 53 | 819 | 6134 | 2578 | <20 | <20 | 73 | 154 | 113 | <20 | <20 | <20 | <20 | <20 | <20 | <20 | <20 | 103 | 44 | <20 | |
| Virion | X1632 | 398F1 | 25710 | BJX2000 | CE0217 | CE1176 | CH119 | CNE8 | CNE55 | TRO.11 | X2278 | 246F3 | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Clade | G | A | C | CRF07 | C | C | CRF07 | CRF01 | CRF01 | B | B | AC | |
| Group | Rabbit | ID50 at week 30 | |||||||||||
| DDMMPP GagM + gp150 | 6826 | <20 | <20 | <20 | <20 | <20 | <20 | <20 | <20 | <20 | <20 | <20 | <20 |
| 6827 | <20 | 21 | <20 | <20 | <20 | <20 | <20 | <20 | <20 | <20 | <20 | <20 | |
| 6830 | <20 | 24 | <20 | <20 | <20 | <20 | <20 | <20 | <20 | <20 | <20 | <20 | |
| 6850 | <20 | <20 | <20 | <20 | <20 | <20 | <20 | <20 | <20 | <20 | <20 | <20 | |
| DDMMPP GagM + gp140HA2tr | 6839 | <20 | 22 | <20 | <20 | <20 | <20 | <20 | <20 | <20 | <20 | <20 | <20 |
| 6840 | <20 | 22 | <20 | <20 | <20 | <20 | <20 | <20 | <20 | <20 | <20 | <20 | |
| 6842 | <20 | 24 | <20 | <20 | <20 | <20 | <20 | <20 | <20 | <20 | <20 | <20 | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chapman, R.; van Diepen, M.; Galant, S.; Kruse, E.; Margolin, E.; Ximba, P.; Hermanus, T.; Moore, P.; Douglass, N.; Williamson, A.-L.; et al. Immunogenicity of HIV-1 Vaccines Expressing Chimeric Envelope Glycoproteins on the Surface of Pr55 Gag Virus-Like Particles. Vaccines 2020, 8, 54. https://doi.org/10.3390/vaccines8010054
Chapman R, van Diepen M, Galant S, Kruse E, Margolin E, Ximba P, Hermanus T, Moore P, Douglass N, Williamson A-L, et al. Immunogenicity of HIV-1 Vaccines Expressing Chimeric Envelope Glycoproteins on the Surface of Pr55 Gag Virus-Like Particles. Vaccines. 2020; 8(1):54. https://doi.org/10.3390/vaccines8010054
Chicago/Turabian StyleChapman, Rosamund, Michiel van Diepen, Shireen Galant, Elizabeth Kruse, Emmanuel Margolin, Phindile Ximba, Tandile Hermanus, Penny Moore, Nicola Douglass, Anna-Lise Williamson, and et al. 2020. "Immunogenicity of HIV-1 Vaccines Expressing Chimeric Envelope Glycoproteins on the Surface of Pr55 Gag Virus-Like Particles" Vaccines 8, no. 1: 54. https://doi.org/10.3390/vaccines8010054
APA StyleChapman, R., van Diepen, M., Galant, S., Kruse, E., Margolin, E., Ximba, P., Hermanus, T., Moore, P., Douglass, N., Williamson, A. -L., & Rybicki, E. (2020). Immunogenicity of HIV-1 Vaccines Expressing Chimeric Envelope Glycoproteins on the Surface of Pr55 Gag Virus-Like Particles. Vaccines, 8(1), 54. https://doi.org/10.3390/vaccines8010054