MVA Vectored Vaccines Encoding Rift Valley Fever Virus Glycoproteins Protect Mice against Lethal Challenge in the Absence of Neutralizing Antibody Responses

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Generation of Recombinant rMVA Encoding RVFV Gn and Gc Glycoproteins
2.2. Western Blot Analysis
2.3. Indirect Immunofluorescence and Laser Confocal Microscopy
2.4. Immunoprecipitation Analysis
2.5. Immunization, Sampling for Immunological Assays and RVFV Challenge
2.6. Assessment of RVFV Serum Neutralizing Antibodies
2.7. Analysis of T-Cell Responses Against RVFV Glycoproteins
2.8. Passive Transfer of Antibodies
2.9. Statistical Analysis
3. Results
3.1. Expression of Recombinant Gn and Gc Glycoproteins in rMVA Infected Cells
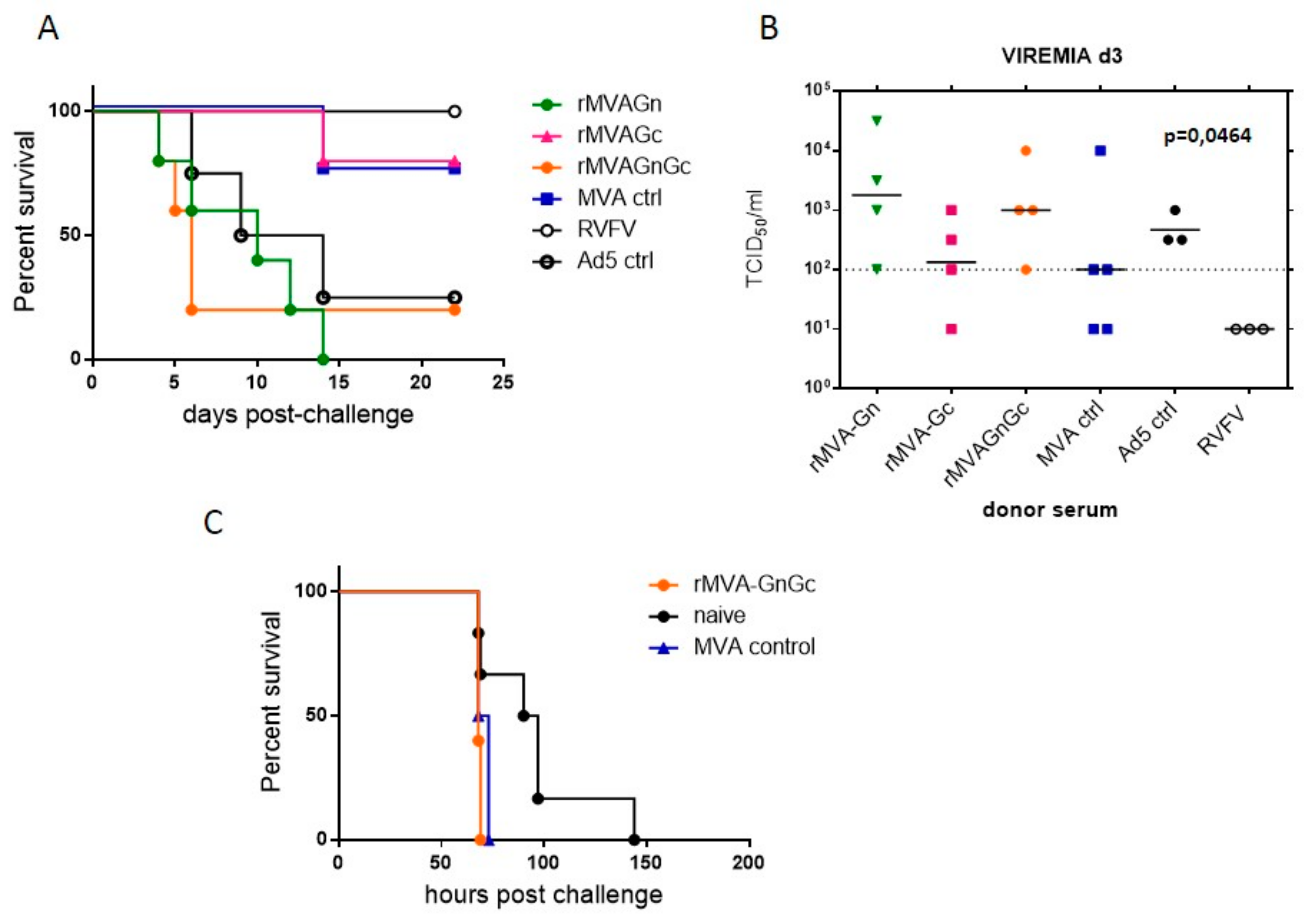
3.2. Efficacy Assessment of MVA Vaccines in Mice
3.3. Analysis of Humoral Responses in rMVA Vaccinated Mice
3.4. Analysis of Cellular Immune Responses to Vaccination
3.5. Assessment of Efficacy of Humoral Responses by Passive Serum Transfer Experiments
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Pepin, M.; Bouloy, M.; Bird, B.H.; Kemp, A.; Paweska, J. Rift Valley fever virus (Bunyaviridae: Phlebovirus): An update on pathogenesis, molecular epidemiology, vectors, diagnostics and prevention. Vet. Res. 2010, 41, 61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chevalier, V.; Pepin, M.; Plee, L.; Lancelot, R. Rift Valley fever--a threat for Europe? Euro. Surveill. 2010, 15, 19506. [Google Scholar] [PubMed]
- Napp, S.; Chevalier, V.; Busquets, N.; Calistri, P.; Casal, J.; Attia, M.; Elbassal, R.; Hosni, H.; Farrag, H.; Hassan, N.; et al. Understanding the legal trade of cattle and camels and the derived risk of Rift Valley Fever introduction into and transmission within Egypt. PLoS Negl. Trop. Dis. 2018, 12, e0006143. [Google Scholar] [CrossRef] [Green Version]
- Moutailler, S.; Krida, G.; Schaffner, F.; Vazeille, M.; Failloux, A.B. Potential vectors of Rift Valley fever virus in the Mediterranean region. Vector Borne Zoonotic Dis. 2008, 8, 749–753. [Google Scholar] [CrossRef]
- Gomez, C.E.; Najera, J.L.; Krupa, M.; Perdiguero, B.; Esteban, M. MVA and NYVAC as vaccines against emergent infectious diseases and cancer. Curr. Gene Ther. 2011, 11, 189–217. [Google Scholar] [CrossRef]
- Cottingham, M.G.; Carroll, M.W. Recombinant MVA vaccines: Dispelling the myths. Vaccine 2013, 31, 4247–4251. [Google Scholar] [CrossRef]
- Gomez, C.E.; Perdiguero, B.; Garcia-Arriaza, J.; Esteban, M. Clinical applications of attenuated MVA poxvirus strain. Expert Rev. Vaccines 2013, 12, 1395–1416. [Google Scholar] [CrossRef]
- Volz, A.; Fux, R.; Langenmayer, M.C.; Sutter, G. Modified vaccinia virus ankara (MVA)--development as recombinant vaccine and prospects for use in veterinary medicine. Berl. Munch. Tierarztl. Wochenschr. 2015, 128, 464–472. [Google Scholar]
- Bird, B.H.; Nichol, S.T. Breaking the chain: Rift Valley fever virus control via livestock vaccination. Curr. Opin. Virol. 2012, 2, 315–323. [Google Scholar] [CrossRef]
- Besselaar, T.G.; Blackburn, N.K. The synergistic neutralization of Rift Valley fever virus by monoclonal antibodies to the envelope glycoproteins. Arch. Virol. 1992, 125, 239–250. [Google Scholar] [CrossRef]
- Besselaar, T.G.; Blackburn, N.K. The effect of neutralizing monoclonal antibodies on early events in Rift Valley fever virus infectivity. Res. Virol. 1994, 145, 13–19. [Google Scholar] [CrossRef]
- Besselaar, T.G.; Blackburn, N.K.; Meenehan, G.M. Antigenic analysis of Rift Valley fever virus isolates: Monoclonal antibodies distinguish between wild-type and neurotropic virus strains. Res. Virol. 1991, 142, 469–474. [Google Scholar] [CrossRef]
- Suzich, J.A.; Kakach, L.T.; Collett, M.S. Expression strategy of a phlebovirus: Biogenesis of proteins from the Rift Valley fever virus M segment. J. Virol. 1990, 64, 1549–1555. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ikegami, T. Molecular biology and genetic diversity of Rift Valley fever virus. Antiviral Res. 2012, 95, 293–310. [Google Scholar] [CrossRef] [Green Version]
- Halldorsson, S.; Li, S.; Li, M.; Harlos, K.; Bowden, T.A.; Huiskonen, J.T. Shielding and activation of a viral membrane fusion protein. Nat. Commun. 2018, 9, 349. [Google Scholar] [CrossRef] [Green Version]
- Allen, E.R.; Krumm, S.A.; Raghwani, J.; Halldorsson, S.; Elliott, A.; Graham, V.A.; Koudriakova, E.; Harlos, K.; Wright, D.; Warimwe, G.M.; et al. A Protective Monoclonal Antibody Targets a Site of Vulnerability on the Surface of Rift Valley Fever Virus. Cell Rep. 2018, 25, 3750–3758. [Google Scholar] [CrossRef] [Green Version]
- Lopez-Gil, E.; Lorenzo, G.; Hevia, E.; Borrego, B.; Eiden, M.; Groschup, M.; Gilbert, S.C.; Brun, A. A Single Immunization with MVA Expressing GnGc Glycoproteins Promotes Epitope-specific CD8+-T Cell Activation and Protects Immune-competent Mice against a Lethal RVFV Infection. PLoS Negl. Trop. Dis. 2013, 7, e2309. [Google Scholar] [CrossRef] [Green Version]
- Lorenzo, G.; Lopez-Gil, E.; Ortego, J.; Brun, A. Efficacy of different DNA and MVA prime-boost vaccination regimens against a Rift Valley fever virus (RVFV) challenge in sheep 12 weeks following vaccination. Vet. Res. 2018, 49, 21. [Google Scholar] [CrossRef] [Green Version]
- Warimwe, G.M.; Lorenzo, G.; Lopez-Gil, E.; Reyes-Sandoval, A.; Cottingham, M.G.; Spencer, A.J.; Collins, K.A.; Dicks, M.D.; Milicic, A.; Lall, A.; et al. Immunogenicity and efficacy of a chimpanzee adenovirus-vectored Rift Valley fever vaccine in mice. Virol. J. 2013, 10, 349. [Google Scholar] [CrossRef] [Green Version]
- Warimwe, G.M.; Gesharisha, J.; Carr, B.V.; Otieno, S.; Otingah, K.; Wright, D.; Charleston, B.; Okoth, E.; Elena, L.G.; Lorenzo, G.; et al. Chimpanzee Adenovirus Vaccine Provides Multispecies Protection against Rift Valley Fever. Sci. Rep. 2016, 6, 20617. [Google Scholar] [CrossRef]
- Malin, A.S.; Huygen, K.; Content, J.; Mackett, M.; Brandt, L.; Andersen, P.; Smith, S.M.; Dockrell, H.M. Vaccinia expression of Mycobacterium tuberculosis-secreted proteins: Tissue plasminogen activator signal sequence enhances expression and immunogenicity of M. tuberculosis Ag85. Microbes Infect. 2000, 2, 1677–1685. [Google Scholar] [CrossRef]
- Jackel, S.; Eiden, M.; Dauber, M.; Balkema-Buschmann, A.; Brun, A.; Groschup, M.H. Generation and application of monoclonal antibodies against Rift Valley fever virus nucleocapsid protein NP and glycoproteins Gn and Gc. Arch. Virol. 2014, 159, 535–546. [Google Scholar] [CrossRef]
- Busquets, N.; Xavier, F.; Martin-Folgar, R.; Lorenzo, G.; Galindo-Cardiel, I.; del Val, B.P.; Rivas, R.; Iglesias, J.; Rodriguez, F.; Solanes, D.; et al. Experimental infection of young adult European breed sheep with Rift Valley fever virus field isolates. Vector Borne Zoonotic Dis. 2010, 10, 689–696. [Google Scholar] [CrossRef] [PubMed]
- Lorenzo, G.; Martin-Folgar, R.; Hevia, E.; Boshra, H.; Brun, A. Protection against lethal Rift Valley fever virus (RVFV) infection in transgenic IFNAR(-/-) mice induced by different DNA vaccination regimens. Vaccine 2010, 28, 2937–2944. [Google Scholar] [CrossRef] [PubMed]
- Busquets, N.; Lorenzo, G.; Lopez-Gil, E.; Rivas, R.; Solanes, D.; Galindo-Cardiel, I.; Abad, F.X.; Rodriguez, F.; Bensaid, A.; Warimwe, G.; et al. Efficacy assessment of an MVA vectored Rift Valley Fever vaccine in lambs. Antiviral Res. 2014, 108, 165–172. [Google Scholar] [CrossRef] [PubMed]
- De Boer, S.M.; Kortekaas, J.; Antonis, A.F.; Kant, J.; van Oploo, J.L.; Rottier, P.J.; Moormann, R.J.; Bosch, B.J. Rift Valley fever virus subunit vaccines confer complete protection against a lethal virus challenge. Vaccine 2010, 28, 2330–2339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beck, Z.; Prohaszka, Z.; Fust, G. Traitors of the immune system-enhancing antibodies in HIV infection: Their possible implication in HIV vaccine development. Vaccine 2008, 26, 3078–3085. [Google Scholar] [CrossRef]
- Hohdatsu, T.; Yamada, M.; Tominaga, R.; Makino, K.; Kida, K.; Koyama, H. Antibody-dependent enhancement of feline infectious peritonitis virus infection in feline alveolar macrophages and human monocyte cell line U937 by serum of cats experimentally or naturally infected with feline coronavirus. J. Vet. Med. Sci. 1998, 60, 49–55. [Google Scholar] [CrossRef] [Green Version]
- Takada, A.; Feldmann, H.; Ksiazek, T.G.; Kawaoka, Y. Antibody-dependent enhancement of Ebola virus infection. J. Virol. 2003, 77, 7539–7544. [Google Scholar] [CrossRef] [Green Version]
- Takada, A.; Kawaoka, Y. Antibody-dependent enhancement of viral infection: Molecular mechanisms and in vivo implications. Rev. Med. Virol. 2003, 13, 387–398. [Google Scholar] [CrossRef]
- Tirado, S.M.; Yoon, K.J. Antibody-dependent enhancement of virus infection and disease. Viral Immunol. 2003, 16, 69–86. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.F.; Tseng, S.P.; Yen, C.H.; Yang, J.Y.; Tsao, C.H.; Shen, C.W.; Chen, K.H.; Liu, F.T.; Liu, W.T.; Chen, Y.M.; et al. Antibody-dependent SARS coronavirus infection is mediated by antibodies against spike proteins. Biochem. Biophys. Res. Commun. 2014, 451, 208–214. [Google Scholar] [CrossRef] [PubMed]
- Price, P.J.; Banki, Z.; Scheideler, A.; Stoiber, H.; Verschoor, A.; Sutter, G.; Lehmann, M.H. Complement component C5 recruits neutrophils in the absence of C3 during respiratory infection with modified vaccinia virus Ankara. J. Immunol. 2015, 194, 1164–1168. [Google Scholar] [CrossRef] [Green Version]
- Sullivan, B.L.; Takefman, D.M.; Spear, G.T. Complement can neutralize HIV-1 plasma virus by a C5-independent mechanism. Virology 1998, 248, 173–181. [Google Scholar] [CrossRef] [PubMed] [Green Version]






© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
López-Gil, E.; Moreno, S.; Ortego, J.; Borrego, B.; Lorenzo, G.; Brun, A. MVA Vectored Vaccines Encoding Rift Valley Fever Virus Glycoproteins Protect Mice against Lethal Challenge in the Absence of Neutralizing Antibody Responses. Vaccines 2020, 8, 82. https://doi.org/10.3390/vaccines8010082
López-Gil E, Moreno S, Ortego J, Borrego B, Lorenzo G, Brun A. MVA Vectored Vaccines Encoding Rift Valley Fever Virus Glycoproteins Protect Mice against Lethal Challenge in the Absence of Neutralizing Antibody Responses. Vaccines. 2020; 8(1):82. https://doi.org/10.3390/vaccines8010082
Chicago/Turabian StyleLópez-Gil, Elena, Sandra Moreno, Javier Ortego, Belén Borrego, Gema Lorenzo, and Alejandro Brun. 2020. "MVA Vectored Vaccines Encoding Rift Valley Fever Virus Glycoproteins Protect Mice against Lethal Challenge in the Absence of Neutralizing Antibody Responses" Vaccines 8, no. 1: 82. https://doi.org/10.3390/vaccines8010082
APA StyleLópez-Gil, E., Moreno, S., Ortego, J., Borrego, B., Lorenzo, G., & Brun, A. (2020). MVA Vectored Vaccines Encoding Rift Valley Fever Virus Glycoproteins Protect Mice against Lethal Challenge in the Absence of Neutralizing Antibody Responses. Vaccines, 8(1), 82. https://doi.org/10.3390/vaccines8010082