The Neisseria gonorrhoeae Vaccine Candidate NHBA Elicits Antibodies That Are Bactericidal, Opsonophagocytic and That Reduce Gonococcal Adherence to Epithelial Cells


Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Growth Conditions
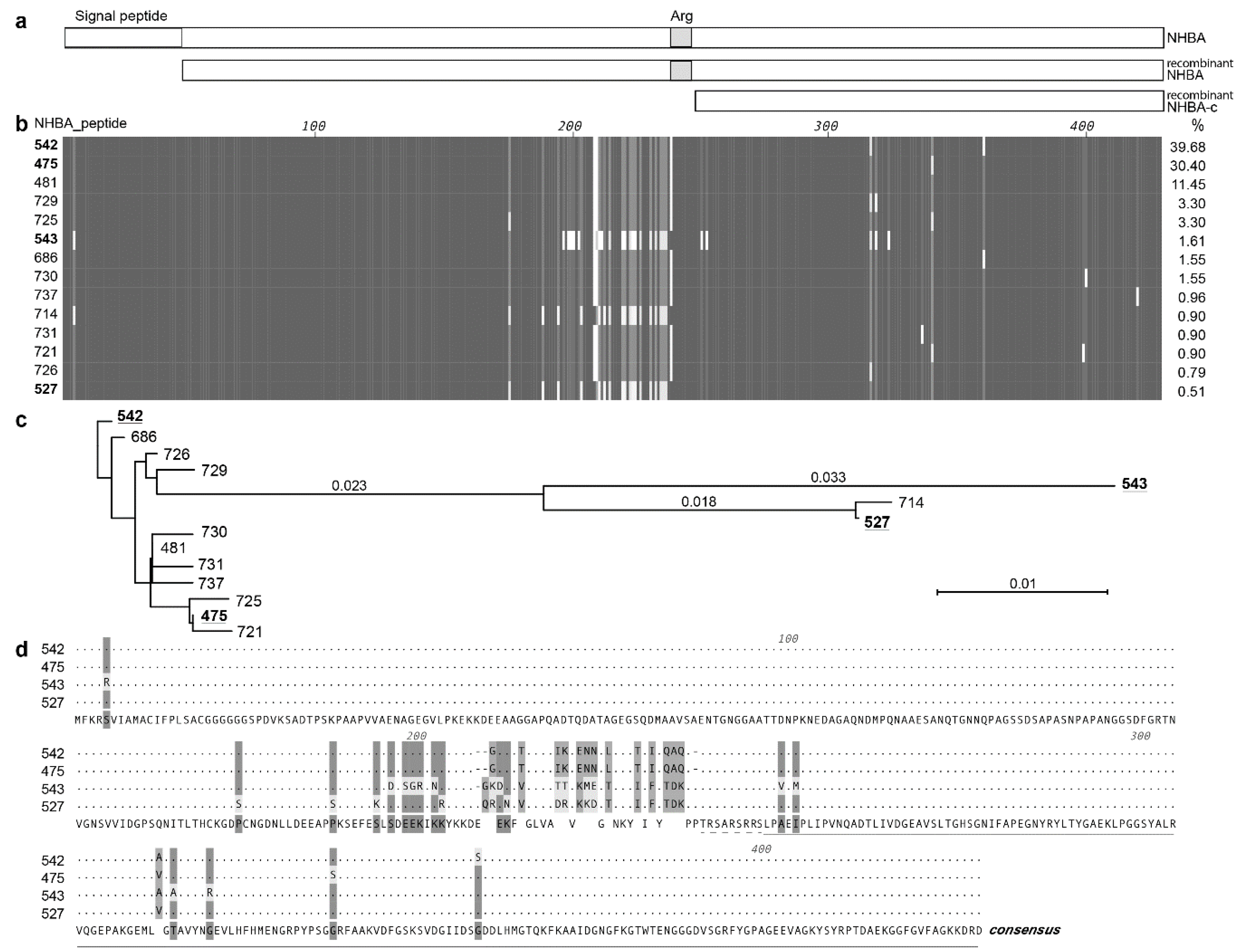
2.2. Sequence Analysis
2.3. Construction of N. Gonorrhoeae NHBA Mutant Strains
2.4. Recombinant Protein Expression
2.5. Generation of Polyclonal Antibodies
2.6. Enzyme-Linked Immunosorbent Assays (ELISAs)
2.7. Serum Bactericidal Activity (SBA) and Opsonophagocytic Killing (OPA) Assays
2.8. Flow Cytometry Analysis
2.9. Surface Plasmon Resonance (SPR)
2.10. Epithelial Cell Adherence Assays
3. Results
3.1. NHBA Is Highly Conserved in N. Gonorrhoeae
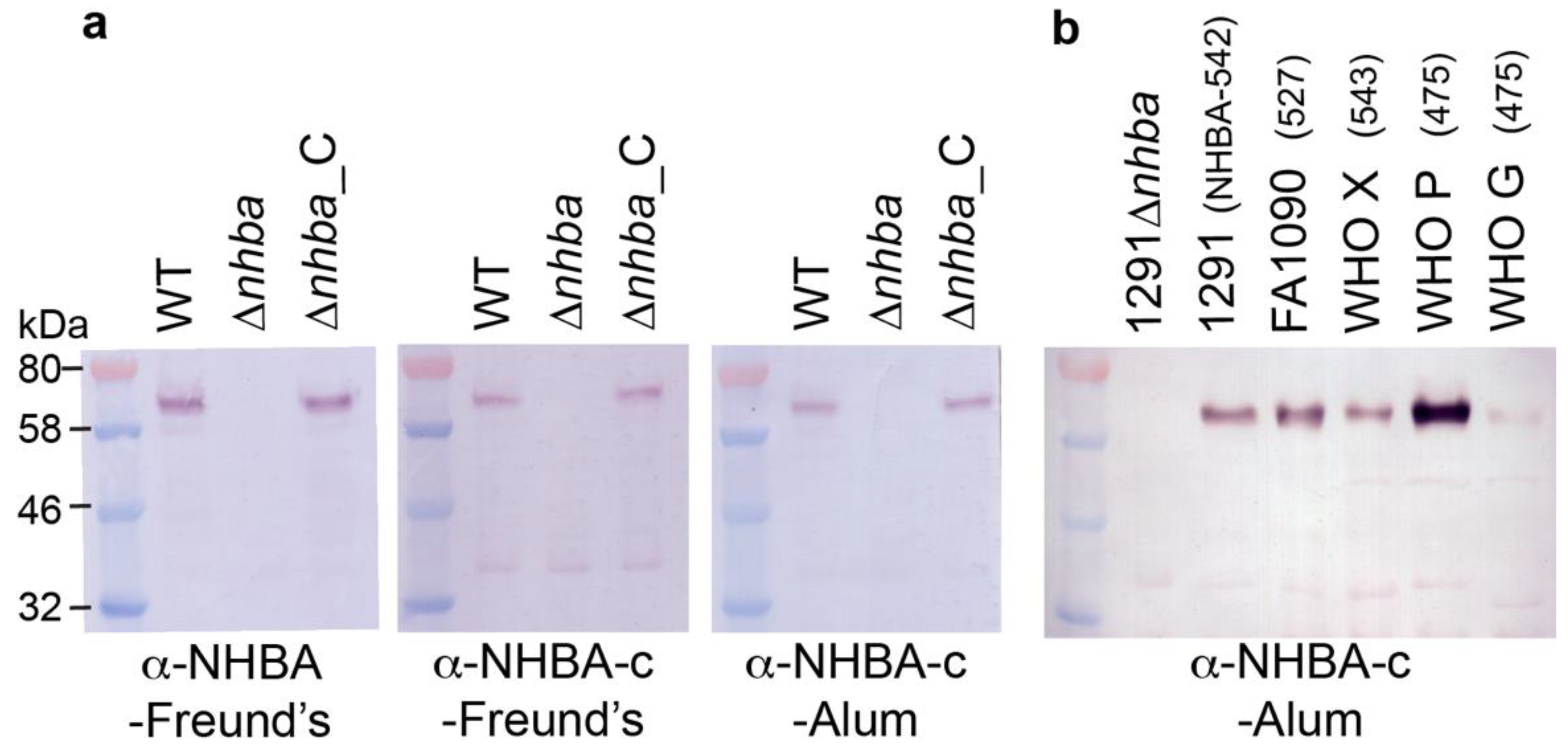
3.2. The Recombinant Full Length NHBA and the C-Terminal NHBA Fragment Are Immunogenic and Induce Antibodies that Recognize NHBA Variants from a Range of Gonococcal Strains
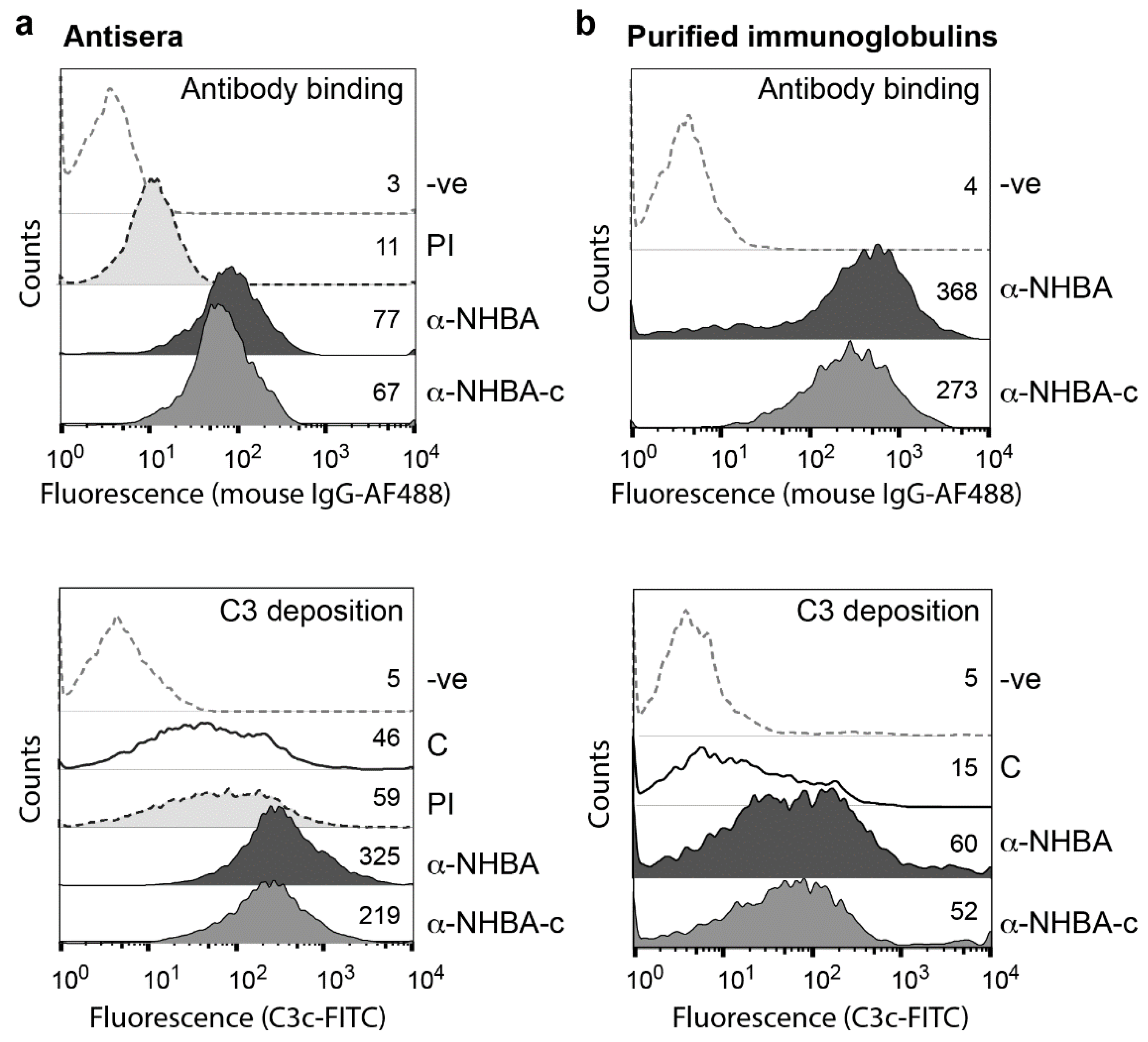
3.3. NHBA Antibodies Promote C3-Fragment Deposition
3.4. NHBA Antibodies Have Bactericidal and Opsonophagocytic Activity
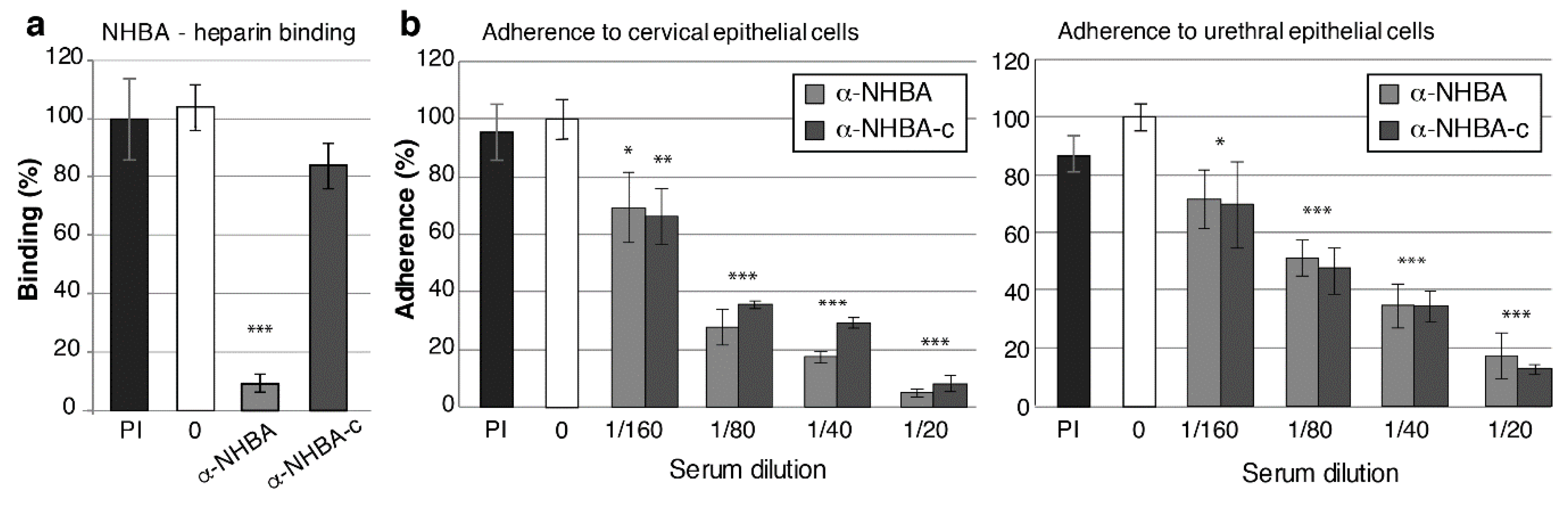
3.5. NHBA Antibodies Reduce NHBA Binding to Heparin, and Gonococcal Adherence to Host Cells
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Whiley, D.M.; Jennison, A.; Pearson, J.; Lahra, M.M. Genetic characterisation of Neisseria gonorrhoeae resistant to both ceftriaxone and azithromycin. Lancet Infect. Dis. 2018, 18, 717–718. [Google Scholar] [CrossRef]
- European Centre for Disease Prevention and Control. Extensively Drug-Resistant (XDR) Neisseria Gonorrhoeae in the United Kingdom and Australia. European Centre for Disease Prevention and Control. Available online: https://www.ecdc.europa.eu/en/publications-data/rapid-risk-assessment-extensively-drug-resistant-xdr-neisseria-gonorrhoeae-united (accessed on 8 April 2020).
- Global Priority List of Antibiotic-Resistant Bacteria to Guide Research, Discovery, and Development of New Antibiotics. Available online: http://www.who.int/medicines/publications/global-priority-list-antibiotic-resistant-bacteria/en/ (accessed on 8 April 2020).
- Centre for Disease Control and Prevention. Available online: http://www.cdc.gov/drugresistance/threat-report-2013/pdf/ar-threats-2013-508.pdf (accessed on 8 April 2020).
- Department of Health. Responding to the Threat of Antimicrobial Resistance: Australia’s First National Antimicrobial Resistance Strategy 2015–2019; Department of Health, Australian Government: Canberra, Australia, 2015.
- World Health Organisation. Global Incidence and Prevalence of Selected Curable Sexually Transmitted Infections—2008. World Health Organisation, 2012. Available online: http://www.who.int/reproductivehealth/publications/rtis/stisestimates/en/ (accessed on 7 March 2020).
- CDC. Sexually Transmitted Disease Surveillance 2017. Available online: https://www.cdc.gov/std/stats 2018 (accessed on 7 March 2020).
- Kirby Institute. HIV, viral hepatitis and sexually transmissible infections in Australia: Annual surveillance report 2018; The Kirby Institute: UNSW, Sydney, Australia, 2018. [Google Scholar]
- Edwards, J.L.; Jennings, M.P.; Apicella, M.A.; Seib, K.L. Is gonococcal disease preventable? The importance of understanding immunity and pathogenesis in vaccine development. Crit. Rev. Microbiol. 2016, 42, 928–941. [Google Scholar] [CrossRef] [Green Version]
- Rice, P.A.; Shafer, W.M.; Ram, S.; Jerse, A.E. Neisseria gonorrhoeae: Drug Resistance, Mouse Models, and Vaccine Development. Annu. Rev. Microbiol. 2017, 71, 665–686. [Google Scholar] [CrossRef] [Green Version]
- Petousis-Harris, H.; Paynter, J.; Morgan, J.; Saxton, P.; McArdle, B.; Goodyear-Smith, F.; Black, S. Effectiveness of a group B outer membrane vesicle meningococcal vaccine against gonorrhoea in New Zealand: A retrospective case-control study. Lancet 2017, 390, 1603–1610. [Google Scholar] [CrossRef]
- Semchenko, E.A.; Tan, A.; Borrow, R.; Seib, K.L. The serogroup B meningococcal vaccine Bexsero elicits antibodies to Neisseria gonorrhoeae. Clin. Infect. Dis. 2018, 69, 1101–1111. [Google Scholar] [CrossRef] [PubMed]
- Serruto, D.; Bottomley, M.J.; Ram, S.; Giuliani, M.M.; Rappuoli, R. The new multicomponent vaccine against meningococcal serogroup B, 4CMenB: Immunological, functional and structural characterization of the antigens. Vaccine 2012, 30, B87–B97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Esposito, V.; Musi, V.; de Chiara, C.; Veggi, D.; Serruto, D.; Scarselli, M.; Kelly, G.; Pizza, M.; Pastore, A. Structure of the C-terminal domain of Neisseria heparin binding antigen (NHBA), one of the main antigens of a novel vaccine against Neisseria meningitidis. J. Biol. Chem. 2011, 286, 41767–41775. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mubaiwa, T.D.; Hartley-Tassell, L.E.; Semchenko, E.A.; Day, C.J.; Jennings, M.P.; Seib, K.L. The Bexsero Neisseria meningitidis serogroup B vaccine antigen NHBA is a high-affinity chondroitin sulfate binding protein. Sci. Rep. 2018, 8, 6512. [Google Scholar] [CrossRef]
- Serruto, D.; Spadafina, T.; Ciucchi, L.; Lewis, L.A.; Ram, S.; Tontini, M.; Santini, L.; Biolchi, A.; Seib, K.L.; Giuliani, M.M.; et al. Neisseria meningitidis GNA2132, a heparin-binding protein that induces protective immunity in humans. Proc. Natl. Acad. Sci. USA 2010, 107, 3770–3775. [Google Scholar] [CrossRef] [Green Version]
- Vacca, I.; Del Tordello, E.; Gasperini, G.; Pezzicoli, A.; Di Fede, M.; Rossi Paccani, S.; Marchi, S.; Mubaiwa, T.D.; Hartley-Tassell, L.E.; Jennings, M.P.; et al. Neisserial Heparin Binding Antigen (NHBA) Contributes to the Adhesion of Neisseria meningitidis to Human Epithelial Cells. PLoS ONE 2016, 11, e0162878. [Google Scholar] [CrossRef] [Green Version]
- Maritan, M.; Veggi, D.; Cozzi, R.; Dello Iacono, L.; Bartolini, E.; Lo Surdo, P.; Maruggi, G.; Spraggon, G.; Bottomley, M.J.; Malito, E. Structures of NHBA elucidate a broadly conserved epitope identified by a vaccine induced antibody. PLoS ONE 2018, 13, e0201922. [Google Scholar] [CrossRef] [PubMed]
- Maritan, M.; Cozzi, R.; Lo Surdo, P.; Veggi, D.; Bottomley, M.J.; Malito, E. Crystal structures of human Fabs targeting the Bexsero meningococcal vaccine antigen NHBA. Acta Crystallogr. F. Struct. Biol. Commun. 2017, 73, 305–314. [Google Scholar] [CrossRef] [PubMed]
- Pizza, M.; Scarlato, V.; Masignani, V.; Giuliani, M.M.; Arico, B.; Comanducci, M.; Jennings, G.T.; Baldi, L.; Bartolini, E.; Capecchi, B.; et al. Identification of vaccine candidates against serogroup B meningococcus by whole-genome sequencing. Science 2000, 287, 1816–1820. [Google Scholar] [CrossRef] [PubMed]
- Giuliani, M.M.; Adu-Bobie, J.; Comanducci, M.; Arico, B.; Savino, S.; Santini, L.; Brunelli, B.; Bambini, S.; Biolchi, A.; Capecchi, B.; et al. A universal vaccine for serogroup B meningococcus. Proc. Natl. Acad. Sci. USA 2006, 103, 10834–10839. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Welsch, J.A.; Moe, G.R.; Rossi, R.; Adu-Bobie, J.; Rappuoli, R.; Granoff, D.M. Antibody to genome-derived neisserial antigen 2132, a Neisseria meningitidis candidate vaccine, confers protection against bacteremia in the absence of complement-mediated bactericidal activity. J. Infect. Dis. 2003, 188, 1730–1740. [Google Scholar] [CrossRef] [Green Version]
- Plested, J.S.; Granoff, D.M. Vaccine-induced opsonophagocytic immunity to Neisseria meningitidis group B. Clin. Vaccine Immunol. 2008, 15, 799–804. [Google Scholar] [CrossRef] [Green Version]
- Del Tordello, E.; Vacca, I.; Ram, S.; Rappuoli, R.; Serruto, D. Neisseria meningitidis NalP cleaves human complement C3, facilitating degradation of C3b and survival in human serum. Proc. Natl. Acad. Sci. USA 2014, 111, 427–432. [Google Scholar] [CrossRef] [Green Version]
- Muzzi, A.; Mora, M.; Pizza, M.; Rappuoli, R.; Donati, C. Conservation of meningococcal antigens in the genus Neisseria. Mbio 2013, 4, e00163-13. [Google Scholar] [CrossRef] [Green Version]
- Semchenko, E.A.; Mubaiwa, T.D.; Day, C.J.; Seib, K.L. The gonococcal Neisserial Heparin Binding Antigen (NHBA) is involved in microcolony formation and contributes to serum resistance and adherence to epithelial cells. J. Infect. Dis. 2019, 10. [Google Scholar] [CrossRef]
- Steichen, C.T.; Shao, J.Q.; Ketterer, M.R.; Apicella, M.A. Gonococcal cervicitis: A role for biofilm in pathogenesis. J. Infect. Dis. 2008, 198, 1856–1861. [Google Scholar] [CrossRef] [Green Version]
- Jen, F.E.C.; Semchenko, E.A.; Day, C.J.; Seib, K.L.; Jennings, M.P. The Neisseria gonorrhoeae Methionine Sulfoxide Reductase (MsrA/B) Is a Surface Exposed, Immunogenic, Vaccine Candidate. Front. Immunol. 2019, 10. [Google Scholar] [CrossRef] [PubMed]
- Semchenko, E.A.; Day, C.J.; Seib, K.L. MetQ of Neisseria gonorrhoeae Is a Surface-Expressed Antigen That Elicits Bactericidal and Functional Blocking Antibodies. Infect. Immun. 2017, 85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shaughnessy, J.; Lewis, L.A.; Zheng, B.; Carr, C.; Bass, I.; Gulati, S.; DeOliveira, R.B.; Gose, S.; Reed, G.W.; Botto, M.; et al. Human Factor H Domains 6 and 7 Fused to IgG1 Fc Are Immunotherapeutic against Neisseria gonorrhoeae. J. Immunol. 2018, 201, 2700–2709. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Potter, A.J.; Kidd, S.P.; Edwards, J.L.; Falsetta, M.L.; Apicella, M.A.; Jennings, M.P.; McEwan, A.G. Thioredoxin reductase is essential for protection of Neisseria gonorrhoeae against killing by nitric oxide and for bacterial growth during interaction with cervical epithelial cells. J. Infect. Dis. 2009, 199, 227–235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harvey, H.A.; Post, D.M.; Apicella, M.A. Immortalization of human urethral epithelial cells: A model for the study of the pathogenesis of and the inflammatory cytokine response to Neisseria gonorrhoeae infection. Infect. Immun. 2002, 70, 5808–5815. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rajam, G.; Stella, M.; Kim, E.; Paulos, S.; Boccadifuoco, G.; Serino, L.; Carlone, G.; Medini, D. Meningococcal Antigen Typing System (MATS)-Based Neisseria meningitidis Serogroup B Coverage Prediction for the MenB-4C Vaccine in the United States. mSphere 2017, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Partridge, E.; Lujan, E.; Giuntini, S.; Vu, D.M.; Granoff, D.M. The role of anti-NHba antibody in bactericidal activity elicited by the meningococcal serogroup B vaccine, MenB-4C. Vaccine 2017, 35, 4236–4244. [Google Scholar] [CrossRef]
- Seib, K.L.; Brunelli, B.; Brogioni, B.; Palumbo, E.; Bambini, S.; Muzzi, A.; Dimarcello, F.; Marchi, S.; van der Ende, A.; Aricó, B.; et al. Characterization of Diverse Subvariants of the Meningococcal Factor H (fH) Binding Protein for Their Ability To Bind fH, To Mediate Serum Resistance, and To Induce Bactericidal Antibodies. Infect. Immun. 2011, 79, 970–981. [Google Scholar] [CrossRef] [Green Version]
- Giuntini, S.; Pajon, R.; Ram, S.; Granoff, D.M. Binding of complement factor H to PorB3 and NspA enhances resistance of Neisseria meningitidis to anti-factor H binding protein bactericidal activity. Infect. Immun. 2015, 83, 1536–1545. [Google Scholar] [CrossRef] [Green Version]
- Zhu, T.; McClure, R.; Harrison, O.B.; Genco, C.; Massari, P. Integrated Bioinformatic Analyses and Immune Characterization of New Neisseria gonorrhoeae Vaccine Antigens Expressed during Natural Mucosal Infection. Vaccines (Basel) 2019, 7, 153. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, K.A.; Schneider, H.; Lindstrom, J.A.; Boslego, J.W.; Warren, R.A.; Van de Verg, L.; Deal, C.D.; McClain, J.B.; Griffiss, J.M. Experimental gonococcal urethritis and reinfection with homologous gonococci in male volunteers. Sex Transm. Dis. 2001, 28, 555–564. [Google Scholar] [CrossRef] [PubMed]
- Michaelsen, T.E.; Kolberg, J.; Aase, A.; Herstad, T.K.; Hoiby, E.A. The four mouse IgG isotypes differ extensively in bactericidal and opsonophagocytic activity when reacting with the P1.16 epitope on the outer membrane PorA protein of Neisseria meningitidis. Scand J. Immunol. 2004, 59, 34–39. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sera 1 | Total IgG ELISA Titre | ELISA Titre | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Vs. Whole-Cells 2 | Vs. Recombinant NHBA 3 | ||||||||
| WT | ∆NHBA | ∆NHBA_C | Total IgG | IgG1 | IgG2a | IgG2b | IgG3 | IgM | |
| NHBA-Freund’s | 128,000 | 16,000 | 128,000 | 40,960,000 | 81,920,000 | 25,600 | 51,200 | 102,400 | 409,600 |
| NHBA-c Freund’s | 64,000 | 8000 | 64,000 | 20,480,000 | 40,960,000 | 51,200 | 204,800 | 12,800 | 204,800 |
| NHBA-c alum | 32,000 | 4000 | 32,000 | 10,240,000 | 40,960,000 | 25,600 | 51,200 | 6400 | 102,400 |
| Strain | NHBA Variant | NHBA Expression 1 | Sera 2 | |||||
|---|---|---|---|---|---|---|---|---|
| NHBA Freund’s | NHBA-c Freund’s | NHBA-c alum | ||||||
| SBA | OPA | SBA | OPA | SBA | OPA | |||
| 1291 | 542 | ++ | 100 | 400 | 200 | 800 | 100 | 800 |
| FA1090 | 527 | +++ | 100 | 100 | 100 | 100 | 50 | 100 |
| WHO G | 543 | + | 200 | 200 | 200 | 400 | 100 | 100 |
| WHO X | 475 | ++ | 1600 | 6400 | 1600 | 6400 | 1600 | 3200 |
| WHO P | 475 | ++++ | 200 | 800 | 200 | 800 | 200 | 400 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Semchenko, E.A.; Day, C.J.; Seib, K.L. The Neisseria gonorrhoeae Vaccine Candidate NHBA Elicits Antibodies That Are Bactericidal, Opsonophagocytic and That Reduce Gonococcal Adherence to Epithelial Cells. Vaccines 2020, 8, 219. https://doi.org/10.3390/vaccines8020219
Semchenko EA, Day CJ, Seib KL. The Neisseria gonorrhoeae Vaccine Candidate NHBA Elicits Antibodies That Are Bactericidal, Opsonophagocytic and That Reduce Gonococcal Adherence to Epithelial Cells. Vaccines. 2020; 8(2):219. https://doi.org/10.3390/vaccines8020219
Chicago/Turabian StyleSemchenko, Evgeny A., Christopher J. Day, and Kate L. Seib. 2020. "The Neisseria gonorrhoeae Vaccine Candidate NHBA Elicits Antibodies That Are Bactericidal, Opsonophagocytic and That Reduce Gonococcal Adherence to Epithelial Cells" Vaccines 8, no. 2: 219. https://doi.org/10.3390/vaccines8020219
APA StyleSemchenko, E. A., Day, C. J., & Seib, K. L. (2020). The Neisseria gonorrhoeae Vaccine Candidate NHBA Elicits Antibodies That Are Bactericidal, Opsonophagocytic and That Reduce Gonococcal Adherence to Epithelial Cells. Vaccines, 8(2), 219. https://doi.org/10.3390/vaccines8020219