Limited Cross-Protection against Infectious Bronchitis Provided by Recombinant Infectious Bronchitis Viruses Expressing Heterologous Spike Glycoproteins

 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethical Statement
2.2. Cells and Viruses
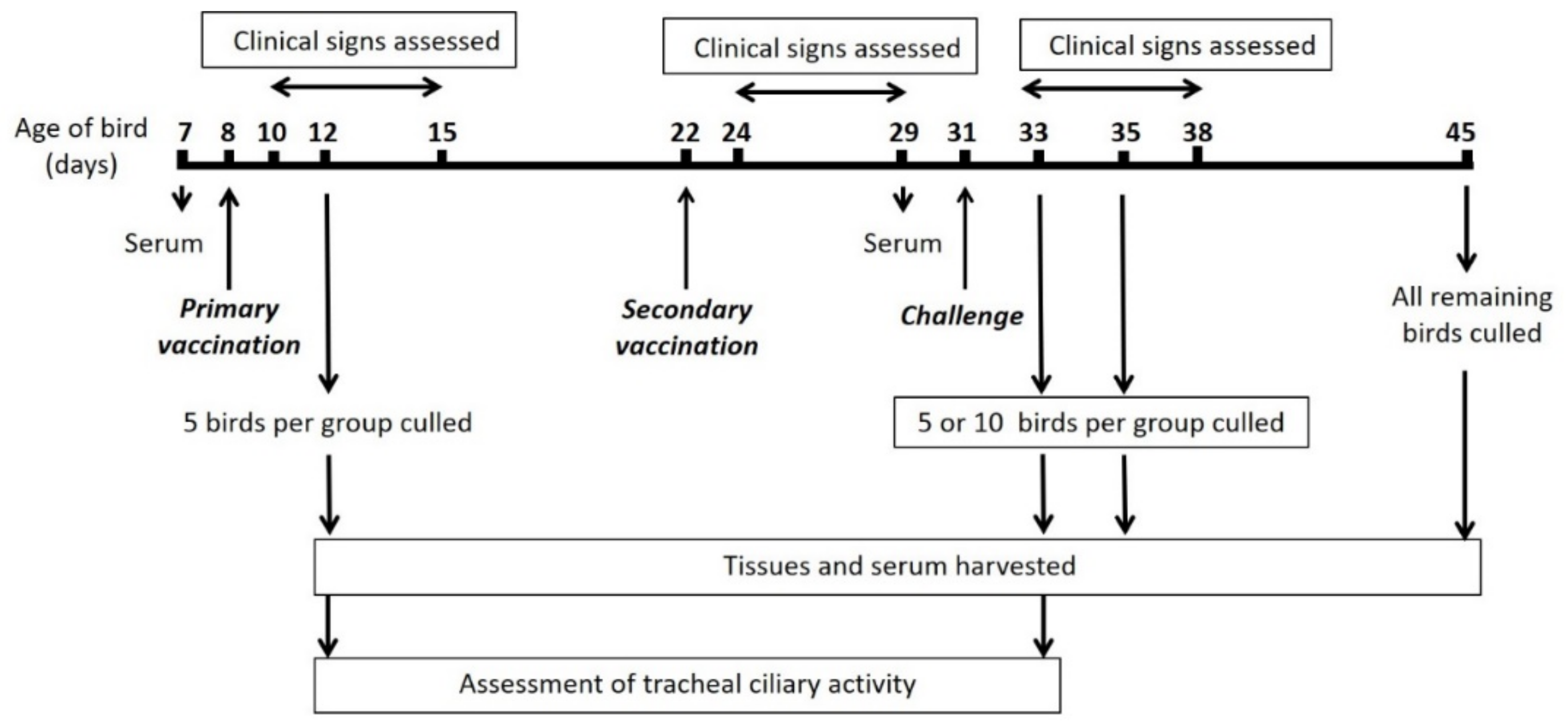
2.3. In Vivo Characterisation
2.4. Ciliary Activity Assay
2.5. Assessment of Infectious Virus in Tracheal Samples
2.6. Infectious Bronchitis Virus ELISA
2.7. Detection of Viral RNA
2.8. Statistics
3. Results
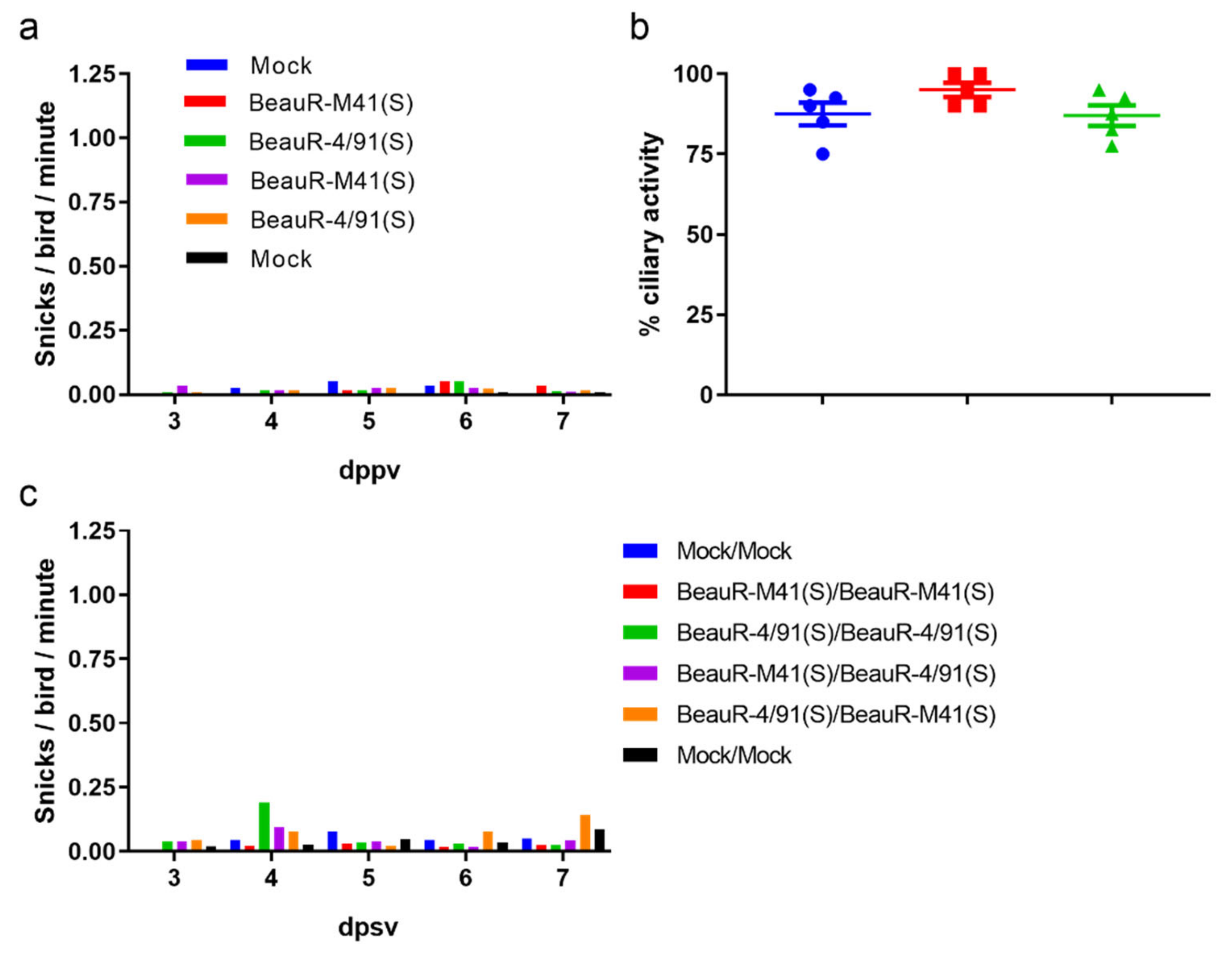
3.1. Confirmation that the S Gene from a Pathogenic Strain Does Not Confer Virulence to a Non-Pathogenic Strain
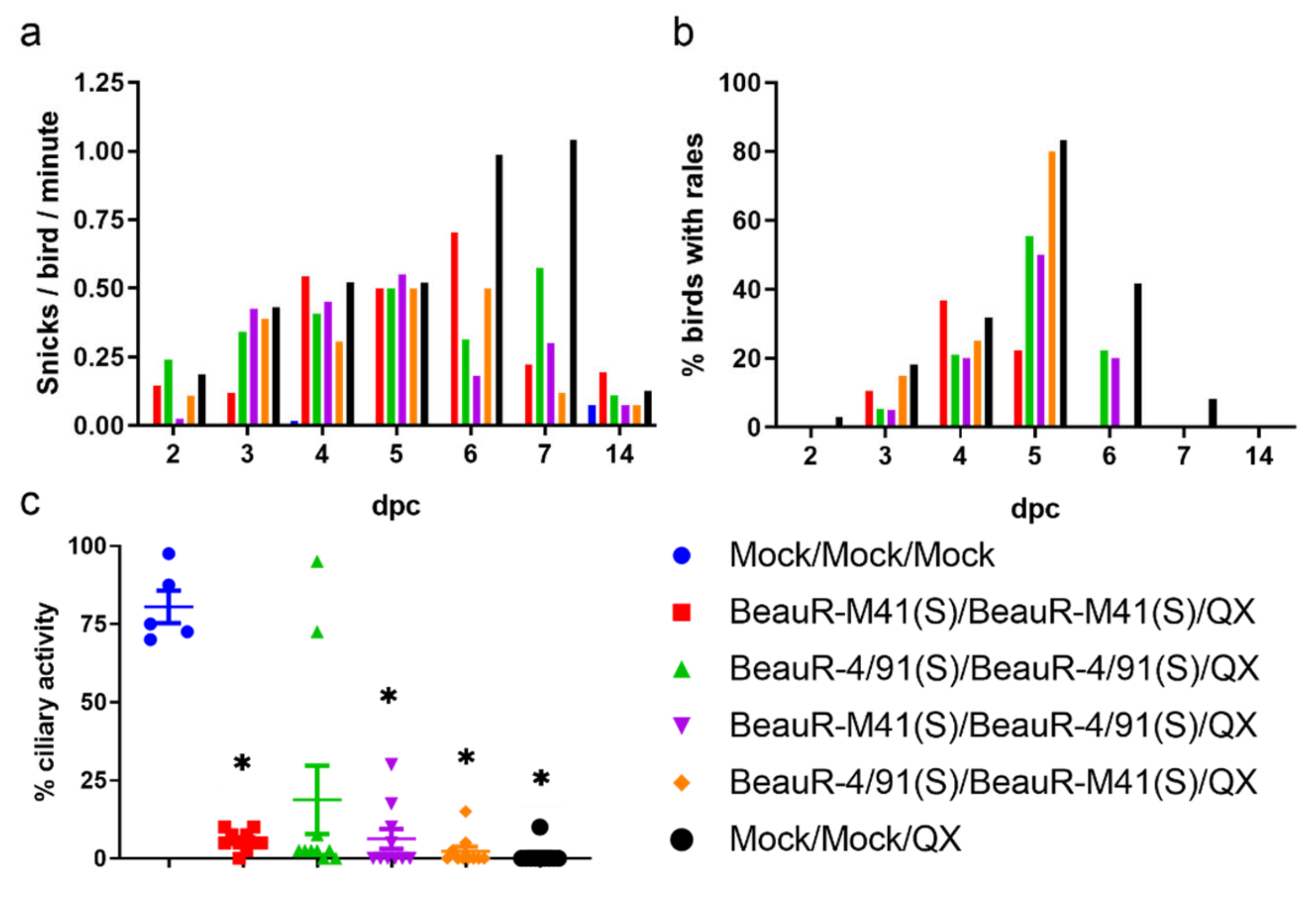
3.2. Chickens Vaccinated with BeauR-M41(S) or BeauR-4/91(S) Exhibited Reduced Clinical Signs in Response to QX Challenge
3.3. Vaccination with BeauR-M41(S) or BeauR-4/91(S) Failed to Protect against Tracheal Ciliostasis Induced by a QX Heterologous Challenge
3.4. Vaccination with a Heterologous Spike Protein Resulted in Reduced Viral Load in Homologous Boosted Vaccinated Groups after QX Challenge
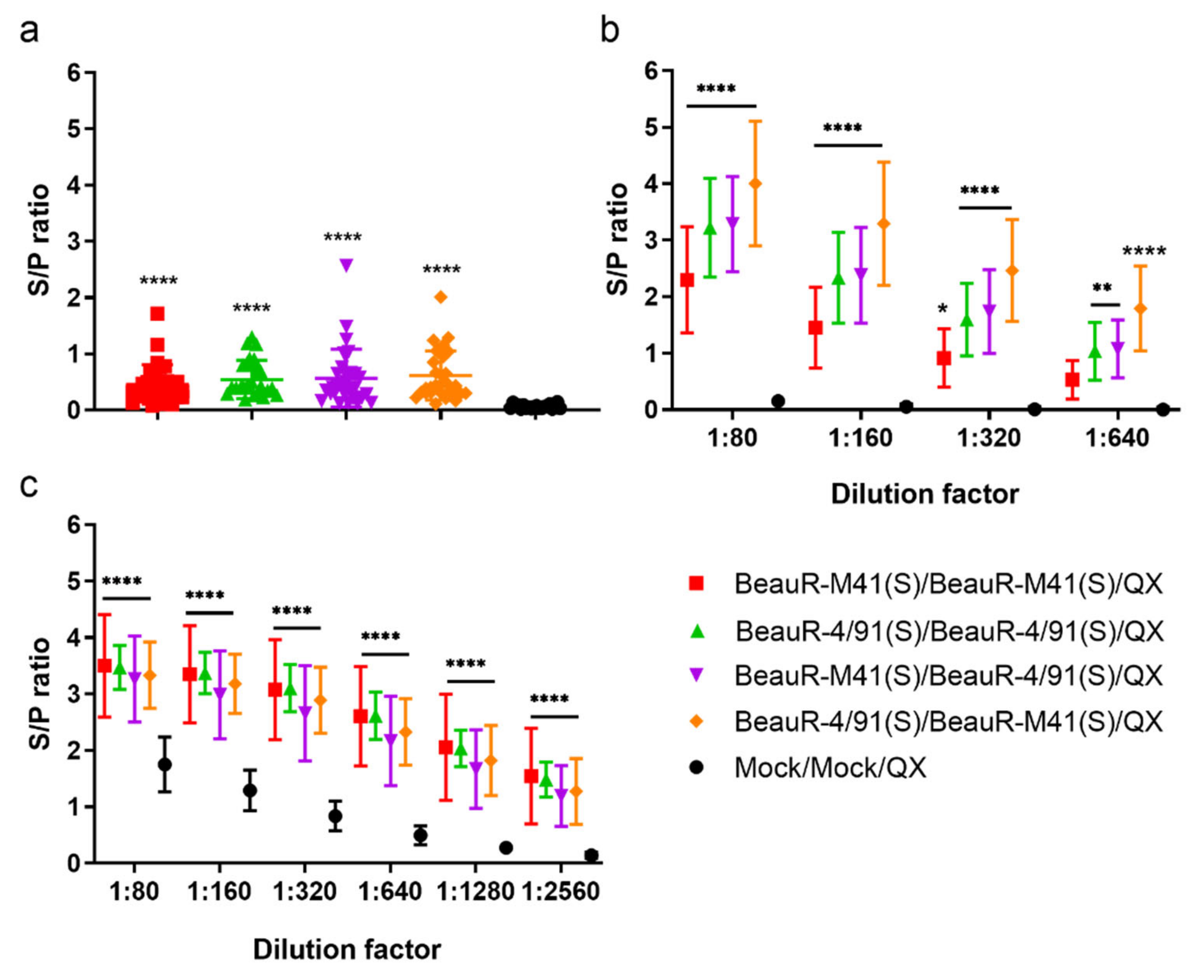
3.5. Vaccination with rIBV Induced the Production of IBV-Specific Antibodies and Primed Chickens for a Boosted Humoral Response to Challenge
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Dinan, A.M.; Keep, S.; Bickerton, E.; Britton, P.; Firth, A.E.; Brierley, I. Comparative Analysis of Gene Expression in Virulent and Attenuated Strains of Infectious Bronchitis Virus at Subcodon Resolution. J. Virol. 2019, 93, e00714. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raj, G.D.; Jones, R.C. Infectious bronchitis virus: Immunopathogenesis of infection in the chicken. Avian Pathol. 1997, 26, 677–706. [Google Scholar] [CrossRef] [PubMed]
- Cavanagh, D. Coronavirus avian infectious bronchitis virus. Veter Res. 2007, 38, 281–297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matthijs, M.G.R.; Ariaans, M.P.; Dwars, R.M.; Van Eck, J.H.; Bouma, A.; Stegeman, J.; Vervelde, L. Course of infection and immune responses in the respiratory tract of IBV infected broilers after superinfection with E. coli. Veter Immunol. Immunopathol. 2009, 127, 77–84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vandekerchove, D.; De Herdt, P.; Laevens, H.; Butaye, P.; Meulemans, G.; Pasmans, F. Significance of interactions between Escherichia coli and respiratory pathogens in layer hen flocks suffering from colibacillosis-associated mortality. Avian Pathol. 2004, 33, 298–302. [Google Scholar] [CrossRef] [Green Version]
- Koch, G.; Hartog, L.; Kant, A.; Van Roozelaar, D.J. Antigenic domains on the peplomer protein of avian infectious bronchitis virus: correlation with biological functions. J. Gen. Virol. 1990, 71, 1929–1935. [Google Scholar] [CrossRef]
- Shang, J.; Zheng, Y.; Yang, Y.; Liu, C.; Geng, Q.; Luo, C.; Zhang, W.; Li, F. Cryo-EM structure of infectious bronchitis coronavirus spike protein reveals structural and functional evolution of coronavirus spike proteins. PLoS Pathog. 2018, 14, e1007009. [Google Scholar] [CrossRef] [Green Version]
- Cavanagh, D.; Davis, P.J. Coronavirus IBV: Removal of Spike Glycopolypeptide S1 by Urea Abolishes Infectivity and Haemagglutination but Not Attachment to Cells. J. Gen. Virol. 1986, 67, 1443–1448. [Google Scholar] [CrossRef]
- Ignjatovic, J.; Galli, L. The S1 glycoprotein but not the N or M proteins of avian infectious bronchitis virus induces protection in vaccinated chickens. Arch. Virol. 1994, 138, 117–134. [Google Scholar] [CrossRef]
- Promkuntod, N.; Van Eijndhoven, R.; De Vrieze, G.; Grone, A.; Verheije, M.H. Mapping of the receptor-binding domain and amino acids critical for attachment in the spike protein of avian coronavirus infectious bronchitis virus. Virology 2014, 448, 26–32. [Google Scholar] [CrossRef] [Green Version]
- Toro, H.; Zhao, W.; Breedlove, C.; Zhang, Z.; Van Santen, V.L.; Yu, Q. Infectious Bronchitis Virus S2 Expressed from Recombinant Virus Confers Broad Protection Against Challenge. Avian Dis. 2014, 58, 83–89. [Google Scholar] [CrossRef]
- Ellis, S.; Keep, S.; Britton, P.; De Wit, S.; Bickerton, E.; Vervelde, L. Recombinant Infectious Bronchitis Viruses Expressing Chimeric Spike Glycoproteins Induce Partial Protective Immunity against Homologous Challenge despite Limited Replication In Vivo. J. Virol. 2018, 92, e01473-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bickerton, E.; Maier, H.J.; Stevenson-Leggett, P.; Armesto, M.; Britton, P. The S2 Subunit of Infectious Bronchitis Virus Beaudette Is a Determinant of Cellular Tropism. J. Virol. 2018, 92, JVI.01044–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bickerton, E.; Dowgier, G.; Britton, P. Recombinant infectious bronchitis viruses expressing heterologous S1 subunits: potential for a new generation of vaccines that replicate in Vero cells. J. Gen. Virol. 2018, 99, 1681–1685. [Google Scholar] [CrossRef] [PubMed]
- Hodgson, T.; Casais, R.; Dove, B.; Britton, P.; Cavanagh, D. Recombinant Infectious Bronchitis Coronavirus Beaudette with the Spike Protein Gene of the Pathogenic M41 Strain Remains Attenuated but Induces Protective Immunity. J. Virol. 2004, 78, 13804–13811. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Armesto, M.; Evans, S.; Cavanagh, D.; Abu-Median, A.B.; Keep, S.; Britton, P. A recombinant avian infectious bronchitis virus expressing a heterologous spike gene belonging to the 4/91 serotype. PLoS ONE 2011, 6, e24352. [Google Scholar] [CrossRef] [Green Version]
- Zhao, R.; Sun, J.; Qi, T.; Zhao, W.; Han, Z.; Yang, X.; Liu, S. Recombinant Newcastle disease virus expressing the infectious bronchitis virus S1 gene protects chickens against Newcastle disease virus and infectious bronchitis virus challenge. Vaccine 2017, 35, 2435–2442. [Google Scholar] [CrossRef]
- Song, C.S.; Jang, H.K.; Lee, Y.J.; Izumiya, Y.; Mikami, T.; Lee, C.W.; Sung, H.W.; Kim, J.H.; Mo, I.P. Induction of protective immunity in chickens vaccinated with infectious bronchitis virus S1 glycoprotein expressed by a recombinant baculovirus. J. Gen. Virol. 1998, 79, 719–723. [Google Scholar] [CrossRef] [Green Version]
- Shirvani, E.; Paldurai, A.; Manoharan, V.K.; Varghese, B.P.; Samal, S.K. A Recombinant Newcastle Disease Virus (NDV) Expressing S Protein of Infectious Bronchitis Virus (IBV) Protects Chickens against IBV and NDV. Sci. Rep. 2018, 8, 11951. [Google Scholar] [CrossRef]
- Compton, S.R.; Ball-Goodrich, L.J.; Zeiss, C.J.; Johnson, L.K.; A Johnson, E.; Macy, J. Pathogenesis of mouse hepatitis virus infection in gamma interferon-deficient mice is modulated by co-infection with Helicobacter hepaticus. Comp. Med. 2003, 53, 197–206. [Google Scholar]
- Terregino, C.; Toffan, A.; Beato, M.S.; De Nardi, R.; Vascellari, M.; Meini, A.; Ortali, G.; Mancin, M.; Capua, I. Pathogenicity of a QX strain of infectious bronchitis virus in specific pathogen free and commercial broiler chickens, and evaluation of protection induced by a vaccination programme based on the Ma5 and 4/91 serotypes. Avian Pathol. 2008, 37, 487–493. [Google Scholar] [CrossRef] [PubMed]
- Cavanagh, D.; Elus, M.M.; Cook, J.K.A. Relationship between sequence variation in the S1 spike protein of infectious bronchitis virus and the extent of cross-protection in vivo. Avian Pathol. 1997, 26, 63–74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cook, J.K.A.; Chesher, J.; Baxendale, W.; Greenwood, N.; Huggins, M.B.; Orbell, S.J. Protection of chickens against renal damage caused by a nephropathogenic infectious bronchitis virus. Avian Pathol. 2001, 30, 423–426. [Google Scholar] [CrossRef] [Green Version]
- Lim, T.-H.; Kim, M.-S.; Jang, J.-H.; Lee, D.-H.; Park, J.-K.; Youn, H.-N.; Lee, J.-B.; Park, S.-Y.; Choi, I.-S.; Song, C.S. Live attenuated nephropathogenic infectious bronchitis virus vaccine provides broad cross protection against new variant strains. Poult. Sci. 2012, 91, 89–94. [Google Scholar] [CrossRef] [PubMed]
- Bru, T.; Vila, R.; Cabana, M.; Geerligs, H. Protection of chickens vaccinated with combinations of commercial live infectious bronchitis vaccines containing Massachusetts, Dutch and QX-like serotypes against challenge with virulent infectious bronchitis viruses 793B and IS/1494/06 Israel variant 2. Avian Pathol. 2016, 46, 52–58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Wit, J.J.; Cook, J.K.A.; Van Der Heijden, H.M.J.F. Infectious bronchitis virus variants: A review of the history, current situation and control measures. Avian Pathol. 2011, 40, 223–235. [Google Scholar] [CrossRef]
- Ladman, B.S.; Loupos, A.B.; Gelb, J. Infectious bronchitis virus S1 gene sequence comparison is a better predictor of challenge of immunity in chickens than serotyping by virus neutralization. Avian Pathol. 2006, 35, 127–133. [Google Scholar] [CrossRef]
- Liu, S.; Zhang, X.; Wang, Y.; Li, C.; Liu, Q.; Han, Z.; Zhang, Q.; Kong, X.; Tong, G. Evaluation of the protection conferred by commercial vaccines and attenuated heterologous isolates in China against the CK/CH/LDL/97I strain of infectious bronchitis coronavirus. Veter J. 2009, 179, 130–136. [Google Scholar] [CrossRef]
- Wang, Y.D.; Wang, Y.L.; Zhang, Z.; Fan, G.; Jiang, Y.; Liu, X.; Ding, J.; Wang, S. Isolation and identification of glandular stomach type IBV (QX IBV) in chickens. Chin. J. Anim. Quar. 1998, 15, 1–3. [Google Scholar]
- Beato, M.S.; De Battisti, C.; Terregino, C.; Drago, A.; Capua, I.; Ortali, G. Evidence of circulation of a Chinese strain of infectious bronchitis virus (QXIBV) in Italy. Veter Rec. 2005, 156, 720. [Google Scholar] [CrossRef]
- Worthington, K.J.; Currie, R.J.W.; Jones, R.C. A reverse transcriptase-polymerase chain reaction survey of infectious bronchitis virus genotypes in Western Europe from 2002 to 2006. Avian Pathol. 2008, 37, 247–257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amin, O.G.M.; Valastro, V.; Salviato, A.; Drago, A.; Cattoli, G.; Monne, I. Circulation of QX-like infectious bronchitis virus in the Middle East. Veter Rec. 2012, 171, 530. [Google Scholar] [CrossRef] [PubMed]
- Ducatez, M.F.; Martin, A.M.; Owoade, A.A.; Olatoye, I.O.; Alkali, B.R.; Maikano, I.; Snoeck, C.J.; Sausy, A.; Cordioli, P.; Muller, C.P. Characterization of a new genotype and serotype of infectious bronchitis virus in Western Africa. J. Gen. Virol. 2009, 90, 2679–2685. [Google Scholar] [CrossRef] [PubMed]
- Khataby, K.; Souiri, A.; Kasmi, Y.; Loutfi, C.; Ennaji, M.M. Current situation, genetic relationship and control measures of infectious bronchitis virus variants circulating in African regions. J. Basic Appl. Zool. 2016, 76, 20–30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bijlenga, G.; Cook, J.K.A.; Gelb, J.J.; De Wit, J.J. Development and use of the H strain of avian infectious bronchitis virus from the Netherlands as a vaccine: A review. Avian Pathol. 2010, 33, 550–557. [Google Scholar] [CrossRef] [Green Version]
- Geerligs, H.; Boelm, G.-J.; Meinders, C.A.M.; Stuurman, B.G.E.; Symons, J.; Tarres-Call, J.; Bru, T.; Vilà, R.; Mombarg, M.; Karaca, K.; et al. Efficacy and safety of an attenuated live QX-like infectious bronchitis virus strain as a vaccine for chickens. Avian Pathol. 2011, 40, 93–102. [Google Scholar] [CrossRef]
- Oade, M.S.; Keep, S.; Freimanis, G.L.; Orton, R.J.; Britton, P.; Hammond, J.A.; Bickerton, E. Attenuation of Infectious Bronchitis Virus in Eggs Results in Different Patterns of Genomic Variation across Multiple Replicates. J. Virol. 2019, 93, e00492. [Google Scholar] [CrossRef] [Green Version]
- Casais, R.; Thiel, V.; Siddell, S.G.; Cavanagh, D.; Britton, P. Reverse Genetics System for the Avian Coronavirus Infectious Bronchitis Virus. J. Virol. 2001, 75, 12359–12369. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.S.; Zhang, Y.; Wang, H.; Fan, W.Q.; Yang, X.; Zhang, A.-Y.; Zeng, F.Y.; Zhang, Z.K.; Cao, H.P.; Zeng, C. Establishment of reverse genetics system for infectious bronchitis virus attenuated vaccine strain H120. Veter Microbiol. 2013, 162, 53–61. [Google Scholar] [CrossRef]
- Van Beurden, S.J.; Berends, A.J.; Krämer-Kühl, A.; Spekreijse, D.; Chénard, G.; Philipp, H.-C.; Mundt, E.; Rottier, P.J.; Verheije, M.H. A reverse genetics system for avian coronavirus infectious bronchitis virus based on targeted RNA recombination. Virol. J. 2017, 14, 109. [Google Scholar] [CrossRef]
- Hennion, R.M.; Hill, G. The Preparation of Chicken Kidney Cell Cultures for Virus Propagation. In Methods in Molecular Biology; Maier, H.J., Bickerton, E., Britton, P., Eds.; Springer Science and Business Media LLC: Berlin/Heidelberg, Germany, 2014; Volume 1282, pp. 57–62. [Google Scholar]
- Hennion, R.M. The Preparation of Chicken Tracheal Organ Cultures for Virus Isolation, Propagation, and Titration. In Methods in Molecular Biology; Maier, H.J., Bickerton, E., Britton, P., Eds.; Springer Science and Business Media LLC: Berlin/Heidelberg, Germany, 2014; Volume 1282, pp. 51–56. [Google Scholar]
- Casais, R.; Dove, B.; Cavanagh, D.; Britton, P. Recombinant Avian Infectious Bronchitis Virus Expressing a Heterologous Spike Gene Demonstrates that the Spike Protein Is a Determinant of Cell Tropism. J. Virol. 2003, 77, 9084–9089. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Armesto, M.; Cavanagh, D.; Britton, P. The Replicase Gene of Avian Coronavirus Infectious Bronchitis Virus Is a Determinant of Pathogenicity. PLoS ONE 2009, 4, e7384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Wit, J.J.; Boelm, G.J.; Van Gerwe, T.J.W.M.; Swart, W.A.J.M. The required sample size in vaccination-challenge experiments with infectious bronchitis virus, a meta-analysis. Avian Pathol. 2013, 42, 9–16. [Google Scholar] [CrossRef] [PubMed]
- Pharmacopeia, E. Avian Infectious Bronchitis Vaccine (Live); European Directorate for the Quality of Medicines and HealthCare (EDQM), Council of Europe: Strasbourg, France, 2010. [Google Scholar]
- Reed, L.; Muench, H. A Simple Method of Estimating Fifty per Cent Endpoints12. Am. J. Epidemiol. 1938, 27, 493–497. [Google Scholar] [CrossRef]
- Callison, S.A.; Hilt, D.A.; Boynton, T.O.; Sample, B.F.; Robison, R.; Swayne, D.E.; Jackwood, M.W. Development and evaluation of a real-time Taqman RT-PCR assay for the detection of infectious bronchitis virus from infected chickens. J. Virol. Methods 2006, 138, 60–65. [Google Scholar] [CrossRef] [PubMed]
- Cook, J.K.A.; Orbell, S.J.; Woods, M.A.; Huggins, M.B. Breadth of protection of the respiratory tract provided by different live-attenuated infectious bronchitis vaccines against challenge with infectious bronchitis viruses of heterologous serotypes. Avian Pathol. 1999, 28, 477–485. [Google Scholar] [CrossRef] [Green Version]
- Awad, F.; Hutton, S.; Forrester, A.; Baylis, M.; Ganapathy, K. Heterologous live infectious bronchitis virus vaccination in day-old commercial broiler chicks: Clinical signs, ciliary health, immune responses and protection against variant infectious bronchitis viruses. Avian Pathol. 2016, 45, 169–177. [Google Scholar] [CrossRef]
- Wang, C.-H.; Hsieh, M.-C.; Chang, P.-C. Isolation, pathogenicity, and H 120 Protection Efficacy of Infectious Bronchitis Viruses Isolated in Taiwan. Avian Dis. 1996, 40, 620. [Google Scholar] [CrossRef]
- Cavanagh, D.; Davis, P.J.; Darbyshire, J.H.; Peters, R.W. Coronavirus IBV: Virus Retaining Spike Glycopolypeptide S2 but Not S1 Is Unable to Induce Virus-neutralizing or Haemagglutination-inhibiting Antibody, or Induce Chicken Tracheal Protection. J. Gen. Virol. 1986, 67, 1435–1442. [Google Scholar] [CrossRef]
- Mockett, A.P.A.; Cavanagh, D.; Brown, T.D.K. Monoclonal Antibodies to the S1 Spike and Membrane Proteins of Avian Infectious Bronchitis Coronavirus Strain Massachusetts M41. J. Gen. Virol. 1984, 65, 2281–2286. [Google Scholar] [CrossRef]
- Cavanagh, D.; Davis, P.J.; Mockett, A. Amino acids within hypervariable region 1 of avian coronavirus IBV (Massachusetts serotype) spike glycoprotein are associated with neutralization epitopes. Virus Res. 1988, 11, 141–150. [Google Scholar] [CrossRef]
- Kant, A.; Koch, G.; van Roozelaar, D.J.; Kusters, J.G.; Poelwijk, F.A.J.; van der Zeijst, B.A.M. Location of antigenic sites defined by neutralising monoclonal antibodies on the S1 avian infectious bronchitis virus glycopolypeptide. J. Gen. Virol. 1992, 73, 591–596. [Google Scholar] [CrossRef] [PubMed]
- Geilhausen, H.E.; Ligon, F.B.; Lukert, P.D. The pathogenesis of virulent and avirulent avian infectious bronchitis virus. Arch. Virol. 1973, 40, 285–290. [Google Scholar] [CrossRef] [PubMed]
- De Wit, J.J.; Wilgen, J.N.-V.; Hoogkamer, A.; Van De Sande, H.; Zuidam, G.J.; Fabri, T.H.F. Induction of cystic oviducts and protection against early challenge with infectious bronchitis virus serotype D388 (genotype QX) by maternally derived antibodies and by early vaccination. Avian Pathol. 2011, 40, 463–471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Wit, J.J.; Cook, J.K.A. Factors influencing the outcome of infectious bronchitis vaccination and challenge experiments. Avian Pathol. 2014, 43, 485–497. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jordan, B. Vaccination against infectious bronchitis virus: A continuous challenge. Veter Microbiol. 2017, 206, 137–143. [Google Scholar] [CrossRef]
- Larsen, F.T.; Bed’Hom, B.; Naghizadeh, M.; Kjaerup, R.B.; Zohari, S.; Dalgaard, T.S. Immunoprofiling of peripheral blood from infectious bronchitis virus vaccinated MHC-B chicken lines—Monocyte MHC-II expression as a potential correlate of protection. Dev. Comp. Immunol. 2019, 96, 93–102. [Google Scholar] [CrossRef]
- Klieve, A.; Cumming, R. Infectious bronchitis: safety and protection in chickens with maternal antibody. Aust. Veter J. 1988, 65, 396–397. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primary Vaccination | No. of Birds Sampled | Secondary Vaccination | Challenge | No. of Birds Sampled | Total no. of Birds Per Group |
|---|---|---|---|---|---|
| Mock | 5 birds on 4 dppv | Mock | Mock | 5 birds on 2, 4 and 14 dpc | 20 |
| BeauR-M41(S) | 5 birds on 4 dppv | BeauR-M41(S) | QX | 10 birds on 2 and 4 dpc; 9 * birds on 14 dpc | 35 |
| BeauR-4/91(S) | 5 birds on 4 dppv | BeauR-4/91(S) | QX | 10 birds on 2 and 4 dpc; 9 birds on 14 dpc | 34 # |
| BeauR-M41(S) | 0 | BeauR-4/91(S) | QX | 10 birds on 2, 4 and 14 dpc | 30 |
| BeauR-4/91(S) | 0 | BeauR-M41(S) | QX | 10 birds on 2, 4 and 14 dpc | 30 |
| Mock | 0 | Mock | QX | 10 birds on 2 and 4 dpc; 12 birds on 14 dpc | 32 |
| Primary Vaccination/ Secondary Vaccination/ Challenge | % Ciliary Activity (Mean ± SD) | No. of Birds with 90% Ciliary Activity/Total No. of Birds Examined | % of Group Protected |
|---|---|---|---|
| Mock/Mock/Mock | 80.50 ± 11.65 | 1/5 | N/A |
| BeauR/M41(S)/BeauR-M41(S)/QX | 5.75 ± 3.13 | 0/10 | 0 |
| BeauR-4/91(S)/BeauR-4/91(S)/QX | 18.75 ± 34.73 | 1/10 | 10 |
| BeauR-M41(S)/BeauR-4/91(S)/QX | 6.25 ± 10.22 | 0/10 | 0 |
| BeauR-4/91(S)/BeauR-M41(S)/QX | 2.25 ± 4.78 | 0/10 | 0 |
| Mock/Mock/QX | 1.00 ± 3.16 | 0/10 | 0 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Keep, S.; Sives, S.; Stevenson-Leggett, P.; Britton, P.; Vervelde, L.; Bickerton, E. Limited Cross-Protection against Infectious Bronchitis Provided by Recombinant Infectious Bronchitis Viruses Expressing Heterologous Spike Glycoproteins. Vaccines 2020, 8, 330. https://doi.org/10.3390/vaccines8020330
Keep S, Sives S, Stevenson-Leggett P, Britton P, Vervelde L, Bickerton E. Limited Cross-Protection against Infectious Bronchitis Provided by Recombinant Infectious Bronchitis Viruses Expressing Heterologous Spike Glycoproteins. Vaccines. 2020; 8(2):330. https://doi.org/10.3390/vaccines8020330
Chicago/Turabian StyleKeep, Sarah, Samantha Sives, Phoebe Stevenson-Leggett, Paul Britton, Lonneke Vervelde, and Erica Bickerton. 2020. "Limited Cross-Protection against Infectious Bronchitis Provided by Recombinant Infectious Bronchitis Viruses Expressing Heterologous Spike Glycoproteins" Vaccines 8, no. 2: 330. https://doi.org/10.3390/vaccines8020330
APA StyleKeep, S., Sives, S., Stevenson-Leggett, P., Britton, P., Vervelde, L., & Bickerton, E. (2020). Limited Cross-Protection against Infectious Bronchitis Provided by Recombinant Infectious Bronchitis Viruses Expressing Heterologous Spike Glycoproteins. Vaccines, 8(2), 330. https://doi.org/10.3390/vaccines8020330