Progress in the Development of Subunit Vaccines against Malaria

 ,
, 



Abstract
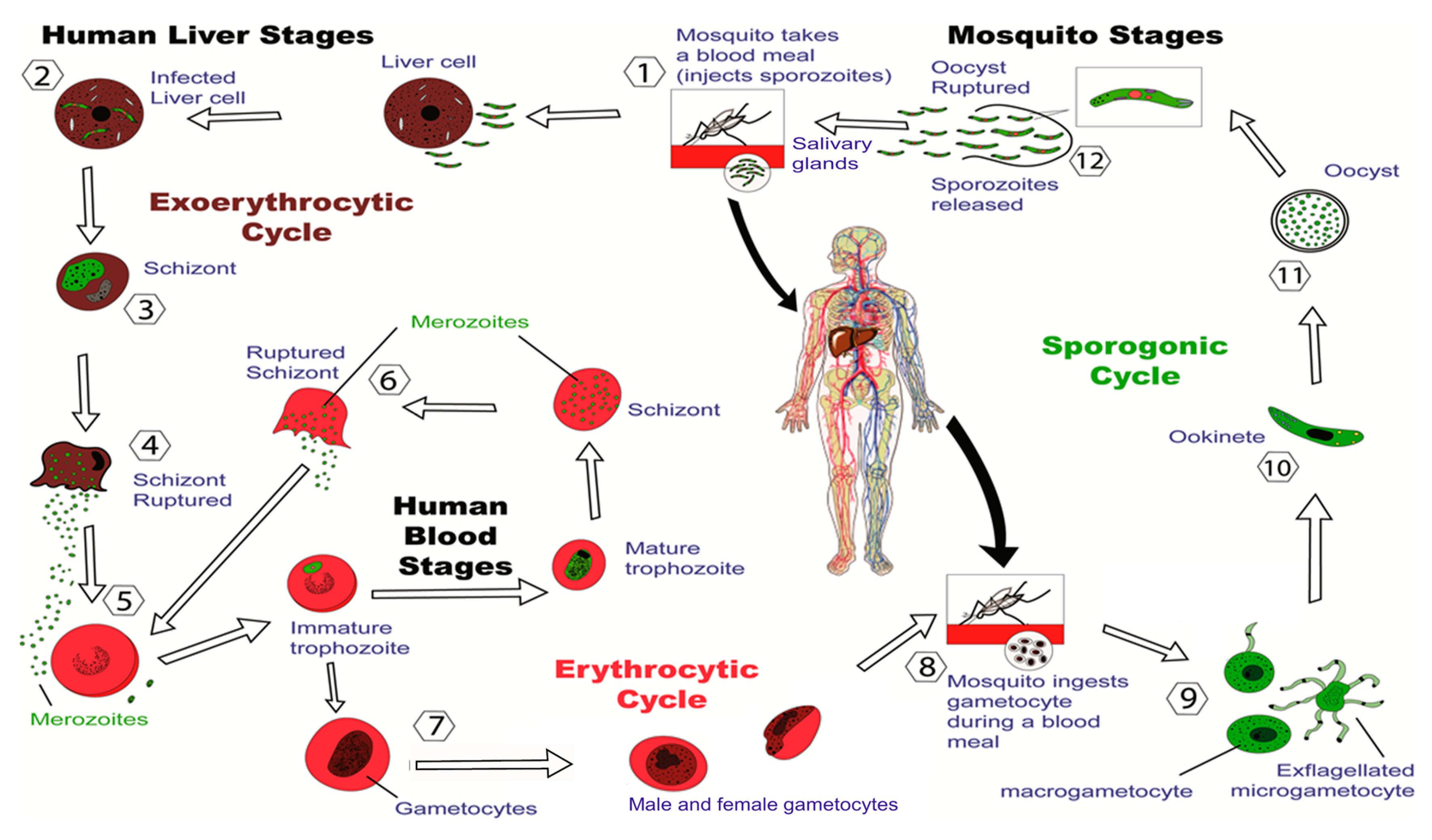
:1. Introduction
2. Peptide-Based Vaccines
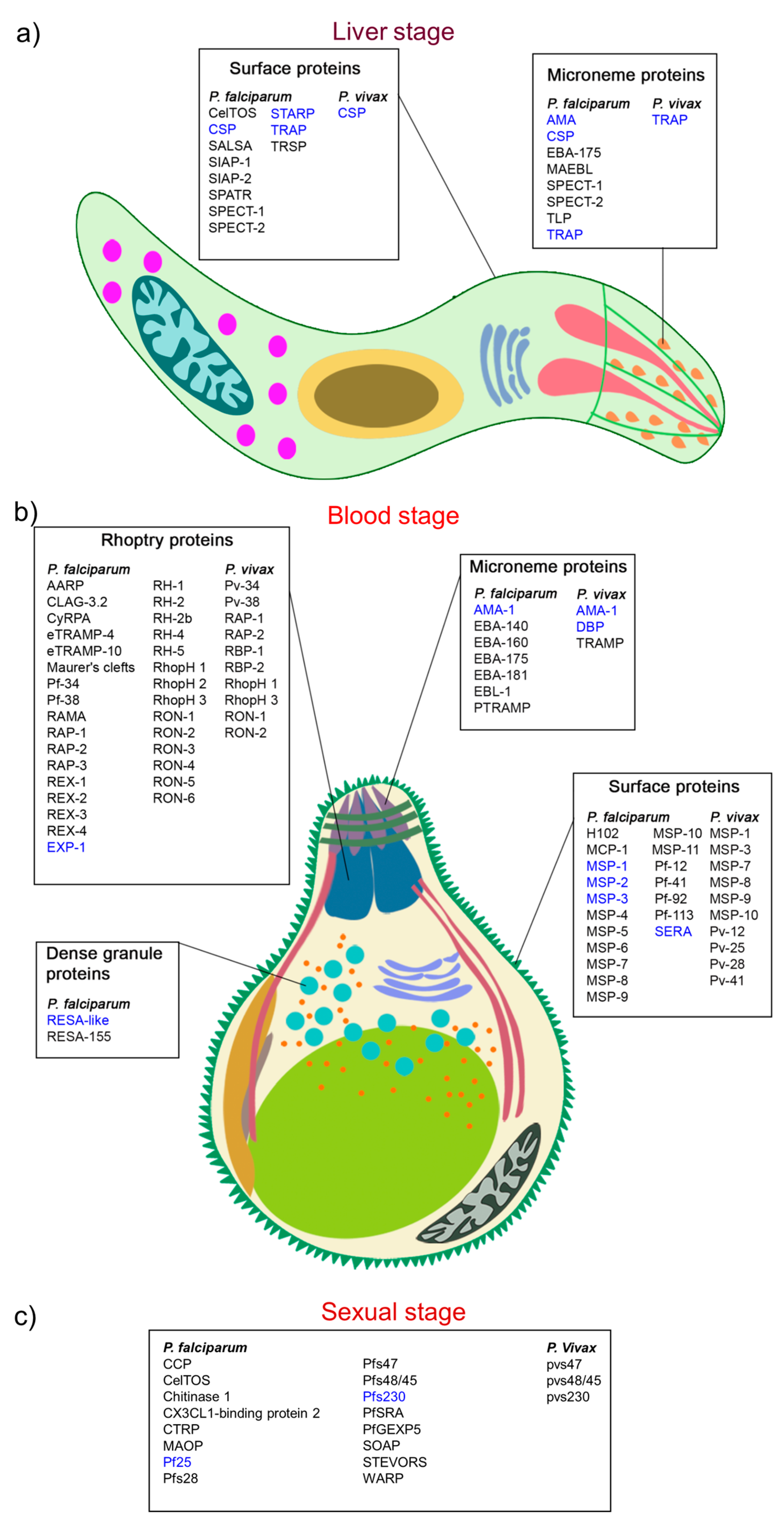
2.1. Pre-Erythrocytic-Stage Vaccines
2.1.1. Circumsporozoite Protein
2.1.2. Other Liver-Stage Peptide Specific Sequences
2.2. Erythrocytic-Stage Vaccines
2.2.1. MSP and Glutamate-Rich Protein
2.2.2. SPf66
2.2.3. Apical Membrane Antigen (AMA-1)
3. Recent Clinical Trials, Challenges and Prospects
3.1. Clinical Trials
3.2. Challenges and Prospects
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Hay, S.I.; Omumbo, J.A.; Craig, M.H.; Snow, R.W. Earth observation, geographic information systems and Plasmodium falciparum malaria in sub-Saharan Africa. Adv. Parasit. 2000, 47, 173–215. [Google Scholar]
- Humphreys, M. Malaria: Poverty, race, and public health in the United States; Johns Hopkins University Press: Baltimore, ML, USA, 2001; p. 196. [Google Scholar]
- Singh, B.; Sung, L.K.; Matusop, A.; Radhakrishnan, A.; Shamsul, S.S.G.; Cox-Singh, J.; Thomas, A.; Conway, D.J. A large focus of naturally acquired Plasmodium knowlesi infections in human beings. Lancet 2004, 363, 1017–1024. [Google Scholar] [CrossRef]
- Guerra, C.A.; Gikandi, P.W.; Tatem, A.J.; Noor, A.M.; Smith, D.L.; Hay, S.I.; Snow, R.W. The limits and intensity of Plasmodium falciparum transmission: Implications for malaria control and elimination worldwide. PloS Med. 2008, 5, 300–311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guerra, C.A.; Howes, R.E.; Patil, A.P.; Gething, P.W.; Van Boeckel, T.P.; Temperley, W.H.; Kabaria, C.W.; Tatem, A.J.; Manh, B.H.; Elyazar, I.R.F.; et al. The international limits and population at risk of Plasmodium vivax transmission in 2009. PloS Negl. Trop D 2010, 4, e774. [Google Scholar] [CrossRef] [PubMed]
- Gallup, J.L.; Sachs, J.D. The economic burden of malaria. Am. J. Trop Med. Hyg. 2001, 64, 85–96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Menendez, C. Malaria during pregnancy-a priority area of malaria research and control. Parasitol. Today 1995, 11, 178–183. [Google Scholar] [CrossRef]
- Monasch, R.; Reinisch, A.; Steketee, R.W.; Korenromp, E.L.; Alnwick, D.; Bergevin, Y. Child coverage with mosquito nets and malaria treatment from population-based surveys in African countries: A baseline for monitoring progress in roll back malaria. Am. J. Trop Med. Hyg. 2004, 71, 232–238. [Google Scholar] [CrossRef] [Green Version]
- Najera, J.A.; Gonzalez-Silva, M.; Alonso, P.L. Some lessons for the future from the global malaria eradication programme (1955-1969). PloS Med. 2011, 8, e1000412. [Google Scholar] [CrossRef] [Green Version]
- van den Berg, H. Global status of DDT and its alternatives for use in vector control to prevent disease. Environ. Health Persp. 2009, 117, 1656–1663. [Google Scholar] [CrossRef]
- Mabaso, M.L.H.; Sharp, B.; Lengeler, C. Historical review of malarial control in southern African with emphasis on the use of indoor residual house-spraying. Trop. Med. Int. Health 2004, 9, 846–856. [Google Scholar] [CrossRef]
- Tren, R.; Bate, R.; Liberty Institute (New Delhi India). When Politics Kills: Malaria and the DDT Story; Liberty Institute. xiv: New Delhi, India, 2000; p. 64. [Google Scholar]
- Peters, W. Plasmodium: Resistance to antimalarial drugs. Ann. Parasitol. Hum. Comp. 1990, 65 (Suppl. 1), 103–106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.G.; Xu, C.C.; Wong, Y.K.; Li, Y.J.; Liao, F.L.; Jiang, T.L.; Tu, Y.Y. Artemisinin, the magic drug discovered from traditional Chinese medicine. Engineering 2019, 5, 32–39. [Google Scholar] [CrossRef]
- Dahl, E.L.; Rosenthal, P.J. Multiple antibiotics exert delayed effects against the Plasmodium falciparum anicoplast. Antimicrob. Agents Chemother. 2007, 51, 3485–3490. [Google Scholar] [CrossRef] [Green Version]
- Gaillard, T.; Madamet, M.; Tsombeng, F.F.; Dormoi, J.; Pradines, B. Antibiotics in malaria therapy: Which antibiotics except tetracyclines and macrolides may be used against malaria? Malar. J. 2016, 15, 556. [Google Scholar] [CrossRef] [Green Version]
- Wynands, R.; Affolderbach, C.; Hollberg, L.; Kitching, J.; Knappe, S.; Stahler, M. Miniaturized laser magnetometers and clocks. Icono 2001 Quantum At. Opt. High Precis. Meas. Opt. Opt. Inf. Process. Transm. Storage 2002, 4750, 126–137. [Google Scholar]
- Perlmann, P.; Troye-Blomberg, M. Malaria and the immune system in humans. Malar. Immunol. 2002, 80, 229–242. [Google Scholar]
- Greenwood, B.; Targett, G. The mysteries of immunity to malaria. Lancet 2011, 377, 1729–1730. [Google Scholar] [CrossRef]
- Doolan, D.L.; Dobano, C.; Baird, J.K. Acquired immunity to malaria. Clin. Microbiol. Rev. 2009, 22, 13–36. [Google Scholar] [CrossRef] [Green Version]
- Ramharter, M.; Winkler, H.; Kremsner, P.G.; Adegnika, A.A.; Willheim, M.; Winkler, S. Age-dependency of Plasmodium falciparum-specific and non-specific T cell cytokine responses in individuals from a malaria-endemic area. Eur. Cytokine Netw. 2005, 16, 135–143. [Google Scholar]
- Nussenzweig, R.S.; Vanderberg, J.; Most, H.; Orton, C. Protective immunity produced by the injection of x-irradiated sporozoites of Plasmodium berghei. Nature 1967, 216, 160–162. [Google Scholar] [CrossRef]
- Giddam, A.K.; Reiman, J.M.; Zaman, M.; Skwarczynski, M.; Toth, I.; Good, M.F. A semi-synthetic whole parasite vaccine designed to protect against blood stage malaria. Acta Biomater. 2016, 44, 295–303. [Google Scholar] [CrossRef] [PubMed]
- Clyde, D.F.; Mccarthy, V.C.; Miller, R.M.; Woodward, W.E. Immunization of man against falciparum and vivax malaria by use of attenuated sporozoites. Am. J. Trop Med. Hyg. 1975, 24, 397–401. [Google Scholar] [CrossRef] [PubMed]
- Hoffman, S.L.; Goh, L.M.L.; Luke, T.C.; Schneider, I.; Le, T.P.; Doolan, D.L.; Sacci, J.; de la Vega, P.; Dowler, M.; Paul, C.; et al. Protection of humans against malaria by immunization with radiation-attenuated Plasmodium falciparum sporozoites. J. Infect. Dis. 2002, 185, 1155–1164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Florens, L.; Washburn, M.P.; Raine, J.D.; Anthony, R.M.; Grainger, M.; Haynes, J.D.; Moch, J.K.; Muster, N.; Sacci, J.B.; Tabb, D.L.; et al. A proteomic view of the Plasmodium falciparum life cycle. Nature 2002, 419, 520–526. [Google Scholar] [CrossRef] [PubMed]
- Bray, R.S.; Garnham, P.C.C. The life-cycle of primate malaria parasites. Brit. Med. Bull. 1982, 38, 117–122. [Google Scholar] [CrossRef] [PubMed]
- Huff, C.G. Life cycle of malarial parasites. Annu. Rev. Microbiol. 1947, 1, 43–60. [Google Scholar] [CrossRef]
- Gosling, R.; von Seidlein, L. The future of the RTS,S/AS01 malaria vaccine: An alternative development plan. PloS Med. 2016, 13, e1001994. [Google Scholar] [CrossRef] [Green Version]
- Tinto, H.; D’Alessandro, U.; Sorgho, H.; Valea, I.; Tahita, M.C.; Kabore, W.; Kiemde, F.; Lompo, P.; Ouedraogo, S.; Derra, K.; et al. Efficacy and safety of RTS,S/AS01 malaria vaccine with or without a booster dose in infants and children in Africa: Final results of a phase 3, individually randomised, controlled trial. Lancet 2015, 386, 31–45. [Google Scholar]
- Regules, J.A.; Cicatelli, S.B.; Bennett, J.W.; Paolino, K.M.; Twomey, P.S.; Moon, J.E.; Kathcart, A.K.; Hauns, K.D.; Komisar, J.L.; Qabar, A.N.; et al. Fractional third and fourth dose of RTS,S/AS01 malaria candidate vaccine: A phase 2a controlled human malaria parasite infection and immunogenicity study. J. Infect. Dis. 2016, 214, 762–771. [Google Scholar] [CrossRef] [Green Version]
- Nielsen, C.M.; Vekemans, J.; Lievens, M.; Kester, K.E.; Regules, J.A.; Ockenhouse, C.F. RTS,S malaria vaccine efficacy and immunogenicity during Plasmodium falciparum challenge is associated with HLA genotype. Vaccine 2018, 36, 1637–1642. [Google Scholar] [CrossRef]
- Mackinnon, M.J.; Marsh, K. The selection landscape of malaria parasites. Science 2010, 328, 866–871. [Google Scholar] [CrossRef] [PubMed]
- Bertolino, P.; Bowen, D.G. Malaria and the liver: Immunological hide-and-seek or subversion of immunity from within? Front. Microbiol. 2015, 6, 41. [Google Scholar] [PubMed] [Green Version]
- Hill, A.V. Vaccines against malaria. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2011, 366, 2806–28014. [Google Scholar] [CrossRef] [Green Version]
- Draper, S.J.; Angov, E.; Horii, T.; Miller, L.H.; Srinivasan, P.; Theisen, M.; Biswas, S. Recent advances in recombinant protein-based malaria vaccines. Vaccine 2015, 33, 7433–7443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skwarczynski, M.; Toth, I. Peptide-based synthetic vaccines. Chem. Sci. 2016, 7, 842–854. [Google Scholar] [CrossRef] [Green Version]
- Malonis, R.J.; Lai, J.R.; Vergnolle, O. Peptide-based vaccines: Current progress and future challenges. Chem. Rev. 2020, 120, 3210–3229. [Google Scholar] [CrossRef] [Green Version]
- Herrington, D.A.; Clyde, D.F.; Losonsky, G.; Cortesia, M.; Murphy, J.R.; Davis, J.; Baqar, S.; Felix, A.M.; Heimer, E.P.; Gillessen, D.; et al. Safety and immunogenicity in man of a synthetic peptide malaria vaccine against Plasmodium-falciparum sporozoites. Nature 1987, 328, 257–259. [Google Scholar] [CrossRef]
- Gerberding, J.L. The new vaccine frontier. SD Med. 2013, Spec no, 123–126. [Google Scholar]
- Beeson, J.G.; Drew, D.R.; Boyle, M.J.; Feng, G.; Fowkes, F.J.I.; Richards, J.S. Merozoite surface proteins in red blood cell invasion, immunity and vaccines against malaria. Fems Microbiol. Rev. 2016, 40, 343–372. [Google Scholar] [CrossRef] [Green Version]
- Gruner, A.C.; Snounou, G.; Brahimi, K.; Letourneur, F.; Renia, L.; Druilhe, P. Pre-erythrocytic antigens of Plasmodium falciparum: From rags to riches? Trends Parasitol. 2003, 19, 74–78. [Google Scholar] [CrossRef]
- Richards, J.S.; Beeson, J.G. The future for blood-stage vaccines against malaria. Immunol. Cell Biol. 2009, 87, 377–390. [Google Scholar] [CrossRef]
- Bolad, A.; Berzins, K. Antigenic diversity of Plasmodium falciparum and antibody-mediated parasite neutralization. Scand. J. Immunol. 2000, 52, 233–239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kengne-Ouafo, J.A.; Sutherland, C.J.; Binka, F.N.; Awandare, G.A.; Urban, B.C.; Dinko, B. Immune responses to the sexual stages of plasmodium falciparum parasites. Front. Immunol. 2019, 10, 136. [Google Scholar] [CrossRef] [PubMed]
- Tachibana, M.; Suwanabun, N.; Kaneko, O.; Iriko, H.; Otsuki, H.; Sattabongkot, J.; Kaneko, A.; Herrera, S.; Torii, M.; Tsuboi, T. Plasmodium vivax gametocyte proteins, Pvs48/45 and Pvs47, induce transmission-reducing antibodies by DNA immunization. Vaccine 2015, 33, 1901–1908. [Google Scholar] [CrossRef] [PubMed]
- White, M.T.; Bejon, P.; Olotu, A.; Griffin, J.T.; Riley, E.M.; Kester, K.E.; Ockenhouse, C.F.; Ghani, A.C. The relationship between RTS,S vaccine-induced antibodies, CD4(+) T cell responses and protection against Plasmodium falciparum infection. PLoS ONE 2013, 8, e61395. [Google Scholar] [CrossRef] [Green Version]
- Garcon, N.; Heppner, D.G.; Cohen, J. Development of RTS,S/AS02: A purified subunit-based malaria vaccine candidate formulated with a novel adjuvant. Expert Rev. Vaccines 2003, 2, 231–238. [Google Scholar] [CrossRef]
- Parra-Lopez, C.A.; Bernal-Estevez, D.; Eduardo Vargas, L.; Pulido-Calixto, C.; Salazar, L.M.; Calvo-Calle, J.M.; Stern, L.J. An unstable Th epitope of P. falciparum fosters central memory T cells and anti-CS antibody responses. PLoS ONE 2014, 9, e0100639. [Google Scholar] [CrossRef] [Green Version]
- Powell, T.J.; Tang, J.; DeRome, M.E.; Mitchell, R.A.; Jacobs, A.; Deng, Y.; Palath, N.; Cardenas, E.; Boyd, J.G.; Nardin, E. Plasmodium falciparum synthetic LbL microparticle vaccine elicits protective neutralizing antibody and parasite-specific cellular immune responses. Vaccine 2013, 31, 1898–1904. [Google Scholar] [CrossRef] [Green Version]
- Mitchell, R.A.; Altszuler, R.; Frevert, U.; Nardin, E.H. Skin scarification with Plasmodium falciparum peptide vaccine using synthetic TLR agonists as adjuvants elicits malaria sporozoite neutralizing immunity. Sci. Rep. 2016, 6, 32575. [Google Scholar] [CrossRef]
- Schwenk, R.; Banania, G.; Epstein, J.; Kim, Y.; Peters, B.; Belmonte, M.; Ganeshan, H.; Huang, J.; Reyes, S.; Stryhn, A.; et al. Ex vivo tetramer staining and cell surface phenotyping for early activation markers CD38 and HLA-DR to enumerate and characterize malaria antigen-specific CD8(+) T-cells induced in human volunteers immunized with a Plasmodium falciparum adenovirus-vectored malaria vaccine expressing AMA1. Malar. J. 2013, 12, 376. [Google Scholar]
- Mueller, M.S.; Renard, A.; Boato, F.; Vogel, D.; Naegeli, M.; Zurbriggen, R.; Robinson, J.A.; Pluschke, G. Induction of parasite growth-inhibitory antibodies by a virosomal formulation of a peptidomimetic of loop I from domain III of Plasmodium falciparum apical membrane antigen 1. Infect. Immun. 2003, 71, 4749–4758. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okitsu, S.L.; Silvie, O.; Westerfeld, N.; Curcic, M.; Kammer, A.R.; Mueller, M.S.; Sauerwein, R.W.; Robinson, J.A.; Genton, B.; Mazier, D.; et al. A virosomal malaria peptide vaccine elicits a long-lasting sporozoite-inhibitory antibody response in a phase 1a clinical trial. PLoS ONE 2007, 2, e1278. [Google Scholar] [CrossRef] [PubMed]
- Cech, P.G.; Aebi, T.; Abdallah, M.S.; Mpina, M.; Machunda, E.B.; Westerfeld, N.; Stoffel, S.A.; Zurbriggen, R.; Pluschke, G.; Tanner, M.; et al. Virosome-formulated Plasmodium falciparum AMA-1 & CSP derived peptides as malaria vaccine: Randomized phase 1b trial in semi-immune adults & children. PLoS ONE 2011, 6, e22273. [Google Scholar]
- Guerinmarchand, C.; Druilhe, P.; Galey, B.; Londono, A.; Patarapotikul, J.; Beaudoin, R.L.; Dubeaux, C.; Tartar, A.; Mercereaupuijalon, O.; Langsley, G. A liver-stage-specific antigen of Plasmodium-falciparum characterized by gene cloning. Nature 1987, 329, 164–167. [Google Scholar] [CrossRef]
- Pratt-Riccio, L.R.; De Souza Perce-Da-Silva, D.; Da Costa Lima-Junior, J.; Pratt Riccio, E.K.; Ribeiro-Alves, M.; Santos, F.; Arruda, M.; Camus, D.; Druilhe, P.; Oliveira-Ferreira, J.; et al. Synthetic antigens derived from Plasmodium falciparum sporozoite, liver, and blood stages: Naturally acquired immune response and human leukocyte antigen associations in individuals living in a Brazilian endemic area. Am. J. Trop Med. Hyg. 2017, 97, 1581–1592. [Google Scholar] [CrossRef] [Green Version]
- Daubersies, P.; Thomas, A.W.; Millet, P.; Brahimi, K.; Langermans, J.A.M.; Ollomo, B.; Ben Mohamed, L.; Slierendregt, B.; Eling, W.; Van Belkum, A.; et al. Protection against Plasmodium falciparum malaria in chimpanzees by immunization with the conserved preerythrocytic liver-stage antigen 3. Nat. Med. 2000, 6, 1258–1263. [Google Scholar] [CrossRef]
- Daubersies, P.; Ollomo, B.; Sauzet, J.P.; Brahimi, K.; Perlaza, B.L.; Eling, W.; Moukana, H.; Rouquet, P.; de Taisne, C.; Druilhe, P. Genetic immunisation by liver stage antigen 3 protects chimpanzees against malaria despite low immune responses. PLoS ONE 2008, 3, e2659. [Google Scholar] [CrossRef]
- Vanloubbeeck, Y.; Pichyangkul, S.; Bayat, B.; Yongvanitchit, K.; Bennett, J.W.; Sattabongkot, J.; Schaecher, K.; Ockenhouse, C.F.; Cohen, J.; Yadava, A. Comparison of the immune responses induced by soluble and particulate Plasmodium vivax circumsporozoite vaccine candidates formulated in AS01 in rhesus macaques. Vaccine 2013, 31, 6216–6224. [Google Scholar] [CrossRef]
- Wilson, K.L.; Xiang, S.D.; Plebanski, M. A model to study the impact of polymorphism driven liver-stage immune evasion by malaria parasites, to help design effective cross-reactive vaccines. Front. Microbiol. 2016, 7, 303. [Google Scholar] [CrossRef]
- Mora, M.d.R.E.; Steeg, C.; Tartz, S.; Heussler, V.; Sparwasser, T.; Link, A.; Fleischer, B.; Jacobs, T. Depletion of regulatory T cells augments a vaccine-induced T effector cell response against the liver-stage of malaria but fails to increase memory. PLoS ONE 2014, 9, e0104627. [Google Scholar]
- Kumar, K.A.; Sano, G.; Boscardin, S.; Nussenzweig, R.S.; Nussenzweig, M.C.; Zavala, F.; Nussenzweig, V. The circumsporozoite protein is an immunodominant protective antigen in irradiated sporozoites. Nature 2006, 444, 937–940. [Google Scholar] [CrossRef] [PubMed]
- Zou, X.Y.; House, B.L.; Zyzak, M.D.; Richie, T.L.; Gerbasi, V.R. Towards an optimized inhibition of liver stage development assay (ILSDA) for Plasmodium falciparum. Malar. J. 2013, 12, 394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gordon, D.M.; Mcgovern, T.W.; Krzych, U.; Cohen, J.C.; Schneider, I.; Lachance, R.; Heppner, D.G.; Yuan, G.; Hollingdale, M.; Slaoui, M.; et al. Safety, immunogenicity, and efficacy of a recombinantly produced Plasmodium-falciparum circumsporozoite-protein hepatitis-B surface-antigen subunit vaccine. J. Infect. Dis. 1995, 171, 1576–1585. [Google Scholar] [CrossRef] [PubMed]
- Cohen, J.D.; Cohen, J. Use of Plasmodium antigen and adjuvant having lipid A derivative and saponin, in manufacture of medicament or in manufacture of kit, for immunizing travelers to endemic regions against productive malaria infection. Patent No. WO2007003384-A1, 11 January 2007. [Google Scholar]
- CalvoCalle, J.M.; Hammer, J.; Sinigaglia, F.; Clavijo, P.; MoyaCastro, Z.R.; Nardin, E.H. Binding of malaria T cell epitopes to DR and DQ molecules in vitro correlates with immunogenicity in vivo-Identification of a universal T cell epitope in the Plasmodium falciparum circumsporozoite protein. J. Immunol. 1997, 159, 1362–1373. [Google Scholar]
- Deoliveira, G.A.; Clavijo, P.; Nussenzweig, R.S.; Nardin, E.H. Immunogenicity of an alum-adsorbed synthetic multiple-antigen peptide-based on B-cell and T-cell epitopes of the Plasmodium-falciparum CS-protein-possible vaccine application. Vaccine 1994, 12, 1012–1017. [Google Scholar] [CrossRef]
- Nardin, E. The past decade in malaria synthetic peptide vaccine clinical trials. Hum. Vaccin. 2010, 6, 27–38. [Google Scholar] [CrossRef] [Green Version]
- Wilson, K.L.; Xiang, S.D.; Plebanski, M. Montanide, Poly I:C and nanoparticle based vaccines promote differential suppressor and effect or cell expansion: A study of induction of CD8 T cells to a minimal Plasmodium berghei epitope. Front. Microbiol. 2015, 6, 29. [Google Scholar] [CrossRef]
- Chandrudu, S.; Skwarczynski, M.; Pattinson, D.; Apte, S.H.; Doolan, D.L.; Toth, I. Synthesis and immunological evaluation of peptide-based vaccine candidates against malaria. Biochem. Compd. 2016, 4, 934141. [Google Scholar] [CrossRef] [Green Version]
- Apte, S.H.; Groves, P.L.; Skwarczynski, M.; Fujita, Y.; Chang, C.H.; Toth, I.; Doolan, D.L. Vaccination with lipid core peptides fails to induce epitope-specific T cell responses but confers non-specific protective immunity in a malaria model. PLoS ONE 2012, 7, e40928. [Google Scholar] [CrossRef]
- Schwartz, L.; Brown, G.V.; Genton, B.; Moorthy, V.S. A review of malaria vaccine clinical projects based on the WHO rainbow table. Malar. J. 2012, 11, 11. [Google Scholar] [CrossRef] [Green Version]
- Fidock, D.A.; Grasmasse, H.; Lepers, J.P.; Brahimi, K.; Benmohamed, L.; Mellouk, S.; Guerinmarchand, C.; Londono, A.; Raharimalala, L.; Meis, J.F.G.M.; et al. Plasmodium-falciparum liver stage antigen-1 is well conserved and contains potent B-cell and T-cell determinants. J. Immunol. 1994, 153, 190–204. [Google Scholar]
- Mikolajczak, S.A.; Sacci, J.B., Jr.; De La Vega, P.; Camargo, N.; VanBuskirk, K.; Krzych, U.; Cao, J.; Jacobs-Lorena, M.; Cowman, A.F.; Kappe, S.H. Disruption of the Plasmodium falciparum liver-stage antigen-1 locus causes a differentiation defect in late liver-stage parasites. Cell Microbiol. 2011, 13, 1250–1260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Druilhe, P.; Guerinmarc, C.; Guerinmarchand, C. New poly peptide used to diagnose malaria-contg. sequences from protein of sporozoite stage of plasmodium falciparum. Patent No. AU9053157-A, 1990. [Google Scholar]
- Cummings, J.F.; Spring, M.D.; Schwenk, R.J.; Ockenhouse, C.F.; Kester, K.E.; Polhemus, M.E.; Walsh, D.S.; Yoon, I.K.; Prosperi, C.; Juompan, L.Y.; et al. Recombinant Liver Stage Antigen-1 (LSA-1) formulated with AS01 or AS02 is safe, elicits high titer antibody and induces IFN-gamma/IL-2 CD4+ T cells but does not protect against experimental Plasmodium falciparum infection. Vaccine 2010, 28, 5135–5144. [Google Scholar] [CrossRef] [PubMed]
- The challenges of introducing a malaria vaccine. Bull World Health Organ 2016, 94, 640–641. [CrossRef]
- Goh, Y.S.; McGuire, D.; Renia, L. Vaccination with Sporozoites: Models and Correlates of Protection. Front. Immunol. 2019, 10, 127. [Google Scholar] [CrossRef] [PubMed]
- Mak, T.W.; Saunders, M.E. 23-Vaccines and Clinical Immunization. In The Immune Response; Mak, T.W., Saunders, M.E., Eds.; Academic Press: Burlington, NJ, USA, 2006; pp. 695–749. [Google Scholar]
- Ouattara, A.; Laurens, M.B. Vaccines against malaria. Clin. Infect. Dis. 2014, 60, 930–936. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soe, S.; Theisen, M.; Roussilhon, C.; Aye, K.S.; Druilhe, P. Association between protection against clinical malaria and antibodies to merozoite surface antigens in an area of hyperendemicity in Myanmar: Complementarity between responses to merozoite surface protein 3 and the 220-kilodalton glutamate-rich protein. Infect. Immun. 2004, 72, 247–252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nebie, I.; Tiono, A.B.; Diallo, D.A.; Samandoulougou, S.; Diarra, A.; Konate, A.T.; Cuzin-Ouattara, N.; Theisen, M.; Corradin, G.; Cousens, S.; et al. Do antibody responses to malaria vaccine candidates influenced by the level of malaria transmission protect from malaria? Trop Med. Int. Health 2008, 13, 229–237. [Google Scholar] [CrossRef]
- Amoah, L.E.; Nuvor, S.V.; Obboh, E.K.; Acquah, F.K.; Asare, K.; Singh, S.K.; Boampong, J.N.; Theisen, M.; Williamson, K.C. Natural antibody responses to Plasmodium falciparum MSP3 and GLURP(R0) antigens are associated with low parasite densities in malaria patients living in the Central Region of Ghana. Parasites Vectors 2017, 10, 395. [Google Scholar] [CrossRef]
- Jepsen, M.P.; Jogdand, P.S.; Singh, S.K.; Esen, M.; Christiansen, M.; Issifou, S.; Hounkpatin, A.B.; Ateba-Ngoa, U.; Kremsner, P.G.; Dziegiel, M.H.; et al. The malaria vaccine candidate GMZ2 elicits functional antibodies in individuals from malaria endemic and non-endemic areas. J. Infect. Dis. 2013, 208, 479–488. [Google Scholar] [CrossRef] [Green Version]
- Graves, P.; Gelband, H. Vaccines for preventing malaria (SPf66). Cochrane Database Syst. Rev. 2006, (Issue 2), Art. No.: CD005966. [Google Scholar] [CrossRef]
- U.S. medical eligibility criteria for contraceptive use, 2016. In Centers for Disease Control and Prevention-Morbidity and Mortality Weekly Report; document No. 3; The Center for Surveillance, Epidemiology, and Laboratory Services, Centers for Disease Control and Prevention (CDC), U.S. Department of Health and Human Services: Atlanta, GA, USA, 2016; Volume 65, 30329-4027.
- Holder, A.A. The carboxy-terminus of merozoite surface protein 1: Structure, specific antibodies and immunity to malaria. Parasitology 2009, 136, 1445–1456. [Google Scholar] [CrossRef]
- Chatterjee, S.; Sharma, P.; Kumar, S.; Chauhan, V.S. Fine specificity of immune responses to epitopic sequences in synthetic peptides containing B and T epitopes from the conserved Plasmodium falciparum blood-stage antigens. Vaccine 1995, 13, 1474–1481. [Google Scholar] [CrossRef]
- Lougovskoi, A.A.; Okoyeh, N.J.; Chauhan, V.S. Mice immunised with synthetic peptide from N-terminal conserved region of merozoite surface antigen-2 of human malaria parasite Plasmodium falciparum can control infection induced by Plasmodium yoelii yoelii 265BY strain. Vaccine 1999, 18, 920–930. [Google Scholar] [CrossRef]
- Sirima, S.B.; Mordmuller, B.; Milligan, P.; Ngoa, U.A.; Kironde, F.; Atuguba, F.; Tiono, A.B.; Issifou, S.; Kaddumukasa, M.; Bangre, O.; et al. A phase 2b randomized, controlled trial of the efficacy of the GMZ2 malaria vaccine in African children. Vaccine 2016, 34, 4536–4542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Theisen, M.; Adu, B.; Mordmuller, B.; Singh, S. The GMZ2 malaria vaccine: From concept to efficacy in humans. Expert Rev. Vaccines 2017, 16, 907–917. [Google Scholar] [CrossRef] [PubMed]
- Wilson, K.L.; Flanagan, K.L.; Prakash, M.D.; Plebanski, M. Malaria vaccines in the eradication era: Current status and future perspectives. Expert Rev. Vaccines 2019, 18, 133–151. [Google Scholar] [CrossRef]
- Amador, R.; Moreno, A.; Murillo, L.A.; Sierra, O.; Saavedra, D.; Rojas, M.; Mora, A.L.; Rocha, C.L.; Alvarado, F.; Falla, J.C.; et al. Safety and immunogenicity of the synthetic malaria vaccine Spf66 in a large field trial. J. Infect. Dis. 1992, 166, 139–144. [Google Scholar] [CrossRef]
- Salcedo, M.; Barreto, L.; Rojas, M.; Moya, R.; Cote, J.; Patarroyo, M.E. Studies on the humoral immune-response to a synthetic vaccine against Plasmodium-falciparum malaria. Clin. Exp. Immunol. 1991, 84, 122–128. [Google Scholar] [CrossRef]
- Valero, M.V.; Amador, L.R.; Galindo, C.; Figueroa, J.; Bello, M.S.; Murillo, L.A.; Mora, A.L.; Patarroyo, G.; Rocha, C.L.; Rojas, M.; et al. Vaccination with Spf66, a chemically synthesized vaccine, against Plasmodium-falciparum malaria in Colombia. Lancet 1993, 341, 705–710. [Google Scholar] [CrossRef]
- Alonso, P.L.; Tanner, M.; Smith, T.; Hayes, R.J.; Schellenberg, J.A.; Lopez, M.C.; Deazevedo, I.B.; Menendez, C.; Lyimo, E.; Weiss, N.; et al. A trial of the synthetic malaria vaccine Spf66 in Tanzania-rationale and design. Vaccine 1994, 12, 181–186. [Google Scholar] [CrossRef]
- Dalessandro, U.; Leach, A.; Drakeley, C.J.; Bennett, S.; Olaleye, B.O.; Fegan, G.W.; Jawara, M.; Langerock, P.; George, M.O.; Targett, G.A.T.; et al. Efficacy trial of malaria vaccine Spf66 in Gambian infants. Lancet 1995, 346, 462–467. [Google Scholar] [CrossRef]
- Nosten, F.; Luxemburger, C.; Kyle, D.E.; Ballou, W.R.; Wittes, J.; Wah, E.; Chongsuphajaisiddhi, T.; Gordon, D.M.; White, N.J.; Sadoff, J.C.; et al. Randomised double-blind placebo-controlled trial of SPf66 malaria vaccine in children in northwestern Thailand. Lancet 1996, 348, 701–707. [Google Scholar] [CrossRef]
- Horii, T.; Shirai, H.; Jie, L.; Ishii, K.J.; Palacpac, N.Q.; Tougan, T.; Hato, M.; Ohta, N.; Bobogare, A.; Arakaki, N.; et al. Evidences of protection against blood-stage infection of Plasmodium falciparum by the novel protein vaccine SE36. Parasitol. Int. 2010, 59, 380–386. [Google Scholar] [CrossRef]
- Pang, X.L.; Mitamura, T.; Horii, T. Antibodies reactive with the N-terminal domain of Plasmodium falciparum serine repeat antigen inhibit cell proliferation by agglutinating merozoites and schizonts. Infect. Immun. 1999, 67, 1821–1827. [Google Scholar] [CrossRef]
- Palacpac, N.M.Q.; Ntege, E.; Yeka, A.; Balikagala, B.; Suzuki, N.; Shirai, H.; Yagi, M.; Ito, K.; Fukushima, W.; Hirota, Y.; et al. Phase 1b randomized trial and follow-up study in Uganda of the blood-stage malaria vaccine candidate BK-SE36. PLoS ONE 2013, 8, e64073. [Google Scholar] [CrossRef] [Green Version]
- Venkatraman, N.; Anagnostou, N.; Bliss, C.; Bowyer, G.; Wright, D.; Lovgren-Bengtsson, K.; Roberts, R.; Poulton, I.; Lawrie, A.; Ewer, K.; et al. Safety and immunogenicity of heterologous prime-boost immunization with viral-vectored malaria vaccines adjuvanted with Matrix-M (TM). Vaccine 2017, 35, 6208–6217. [Google Scholar] [CrossRef]
- Cox, R.J.; Pedersen, G.; Madhun, A.S.; Svindland, S.; Saevik, M.; Breakwell, L.; Hoschler, K.; Willemsen, M.; Campitelli, L.; Nostbakken, J.K.; et al. Evaluation of a virosomal H5N1 vaccine formulated with Matrix M (TM) adjuvant in a phase I clinical trial. Vaccine 2011, 29, 8049–8059. [Google Scholar] [CrossRef]
- Bigaeva, E.; van Doorn, E.; Liu, H.; Hak, E. Meta-Analysis on Randomized Controlled Trials of Vaccines with QS-21 or ISCOMATRIX Adjuvant: Safety and Tolerability. PLoS ONE 2016, 11, e0154757. [Google Scholar] [CrossRef] [Green Version]
- Bengtsson, K.L.; Karlsson, K.H.; Magnusson, S.E.; Reimer, J.M.; Stertman, L. Matrix-M (TM) adjuvant: Enhancing immune responses by ‘setting the stage’ for the antigen. Expert Rev. Vaccines 2013, 12, 821–823. [Google Scholar] [CrossRef]
- Bengtsson, K.L.; Song, H.; Stertman, L.; Liu, Y.; Flyer, D.C.; Massare, M.J.; Xu, R.H.; Zhou, B.; Lu, H.; Kwilas, S.A.; et al. Matrix-M adjuvant enhances antibody, cellular and protective immune responses of a Zaire Ebola/Makona virus glycoprotein (GP) nanoparticle vaccine in mice. Vaccine 2016, 34, 1927–1935. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCall, M.B.B.; Kremsner, P.G.; Mordmuller, B. Correlating efficacy and immunogenicity in malaria vaccine trials. Semin. Immunol. 2018, 39, 52–64. [Google Scholar] [CrossRef] [PubMed]
- Julien, J.P.; Wardemann, H. Antibodies against Plasmodium falciparum malaria at the molecular level. Nat. Rev. Immunol. 2019, 19, 761–775. [Google Scholar] [CrossRef]
- Lee, W.C.; Russell, B.; Sobota, R.M.; Ghaffar, K.; Howland, S.W.; Wong, Z.X.; Maier, A.G.; Dorin-Semblat, D.; Biswas, S.; Gamain, B.; et al. Plasmodium-infected erythrocytes induce secretion of IGFBP7 to form type II rosettes and escape phagocytosis. Elife 2020, 9, e51546. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.B.; Smith, J.D.; Kappe, S.H.I. Advances and challenges in malaria vaccine development. Expert Rev. Mol. Med. 2009, 11, e39. [Google Scholar] [CrossRef] [Green Version]
- Ssemaganda, A.; Giddam, A.K.; Zaman, M.; Skwarczynski, M.; Toth, I.; Stanisic, D.I.; Good, M.F. Induction of Plasmodium-specific immune responses using liposome-based vaccines. Front. Immunol. 2019, 10, 135. [Google Scholar] [CrossRef]
- Vandoolaeghe, P.; Schuerman, L. The RTS,S/AS01 malaria vaccine in children 5 to 17 months of age at first vaccination. Expert Rev. Vaccines 2016, 15, 1481–1493. [Google Scholar] [CrossRef] [Green Version]
- Jindal, H.; Bhatt, B.; Malik, J.S.; Sk, S.; Mehta, B. Malaria vaccine: A step toward elimination. Hum. Vaccin. Immunother. 2014, 10, 1752–1754. [Google Scholar] [CrossRef] [Green Version]
- Mitran, C.J.; Mena, A.; Gnidehou, S.; Banman, S.; Arango, E.; Lima, B.A.S.; Lugo, H.; Ganesan, A.; Salanti, A.; Mbonye, A.K.; et al. Antibodies to cryptic epitopes in distant homologues underpin a mechanism of heterologous immunity between Plasmodium vivax PvDBP and Plasmodium falciparum VAR2CSA. MBio 2019, 10. [Google Scholar] [CrossRef] [Green Version]
- Pandey, R.K.; Bhatt, T.K.; Prajapati, V.K. Novel immunoinformatics approaches to design multi-epitope subunit vaccine for malaria by investigating Anopheles salivary protein. Sci. Rep. 2018, 8, 1125. [Google Scholar] [CrossRef] [Green Version]
- Karch, C.P.; Doll, T.; Paulillo, S.M.; Nebie, I.; Lanar, D.E.; Corradin, G.; Burkhard, P. The use of a P. falciparum specific coiled-coil domain to construct a self-assembling protein nanoparticle vaccine to prevent malaria. J. Nanobiotechnol. 2017, 15, 62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loeffler, F.F.; Pfeil, J.; Heiss, K. High-density peptide arrays for malaria vaccine development. Methods Mol. Biol. (CliftonN.J.) 2016, 1403, 569–582. [Google Scholar]
- Guleria, V.; Jaiswal, V. Comparative transcriptome analysis of different stages of Plasmodium falciparum to explore vaccine and drug candidates. Genomics 2019, 112, 796–804. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Vaccine Name | Parasite | Source of Peptide Antigens | Adjuvant | Immune Response | Clinical/Animal Trials | References |
|---|---|---|---|---|---|---|
| RTS,S (GlaxoSmithKline Biologicals) | P. falciparum | CSP (207–395); S (the N-terminus of HBsAg) | AS01, AS02 | Protective humoral/cellular | Children and infants | [30,47,48] |
| QNT-5 | P. falciparum | CD4 T-cell epitope from C-terminus of the CS protein | Montanide ISA 720 | Protective humoral | HLA-DR4 transgenic mice | [49] |
| Tri-epitope CS peptide T1BT* | P. falciparum | B-cell epitope from CS repeat region and two T-cell epitopes: conserved T1 epitope and the universal epitope T* | Freund’s adjuvant | Protective cellular | C57BL/6J (H-2b) and BALB/cJ (H-2d) mice | [50] |
| CS repeat peptide | P. falciparum | A protective B-cell peptide epitope of CS repeat region | TLR agonists | Protective humoral | C57BL/6 mice | [51] |
| NMRC-M3V-Ad-PfCA | P. falciparum | CS protein; apical membrane antigen-1 (AMA-1) | ND | Protective cellular | Human adults | [52] |
| Ad-CA, Ad-C | P. falciparum | CS protein; AMA-1 | Protective cellular | Human adults | [52] | |
| AMA49-L1 and AMA49-C1 | P. falciparum | Cyclic and linear AMA1(446–490) peptide | ND | Protective humoral | BALB/c mice | [53] |
| UK-39 and AMA49-C1 | P. falciparum | UK-39 from CSP; AMA49-C1 peptides from AMA-1 | ND | Protective humoral | Human adults | [54] |
| AMA-1 and CSP | P. falciparum | SCP; AMA-1 | ND | Protective cellular | Human adults | [55] |
| LSA-1 | P. falciparum | LSA-1 | ND | No immune response | BALB/c mice | [56] |
| LSA-1 | P. falciparum | LSA-1 | ND | Protective humoral | Human adults | [57] |
| LSA-3 | P. falciparum | LSA-3 | ND | Protective cellular | Chimpanzees | [58,59] |
| VMP001, | P. vivax | N- and C- terminal regions of the CSV and a truncated repeat region that contains repeat sequences from both the VK210 (type 1) and the VK247 (type 2) parasites | AS01 | Protective humoral/cellular | Rhesus monkeys | [60] |
| CSV-S,S | P. vivax | VMP001 and S (the N-terminus of HBsAg) | AS01 | Protective humoral/cellular | Rhesus monkeys | [60] |
| KI | P. berghei | pb9 (SYIPSAEKI, referred to as KI) from CS protein | Montanide, Poly I:C | Protective humoral/cellular | BALB/c mice | [61] |
| ACT-CS | P. berghei | recombinant Bordetella adenylate cyclase toxoid fused with an MHC class I-restricted epitope of the CS protein | ND | Protective humoral/cellular | DEREG-mice | [62] |
| Vaccine Name | Parasite | Source of Peptide Antigen | Adjuvant | Immune Response | Clinical Trials/Animals | References |
|---|---|---|---|---|---|---|
| MSP-1, MSP-3b, and GLURP | P. falciparum | MSP; GLURP27–500; GLURP489–705; GLURP705–1178 | ND | Protective humoral | Human adults | [82] |
| MSP-3, GLURP, and AMA1 | P. falciparum | MSP-3181–276; GLURP94–489; R2 (repeat region amino acids 705–1178 | ND | Protective humoral | Human adults | [83] |
| MSP-3, GLURP | P. falciparum | MSP181–276; GLURP27–500 | ND | Recognized by natural immune response | Human adults | [84] |
| GMZ2 | P. falciparum | Fusion of GLURP27–500 and MSP3212–380 | Al(OH)3 | Recognized by natural immune response | Human adults | [85] |
| Spf66 | P. falciparum | Three merozoite-derived antigens and NANP epitope of CSP | ND | No/little efficacy | Human adults | [78,86,87] |
| Vaccine Name | Target Protein | Vaccination Protocol | Clinical Phase | Antigen Source |
|---|---|---|---|---|
| Pre-erythrocytic stage | ||||
| RTS,S/AS01E | Pf CSP (207–395) and HepBsAg | IM | IV | S. cerevisiae |
| ChAd63/MVA ME-TRAP | TRAP + ME epitopes (CS, LSA1, LSA3, STARP, EXP1, pb9) | IM | IIb | Simian adenovirus ChAd63, MVA |
| ChAd63/MVA ME-TRAP + Matrix M™ | TRAP + ME epitopes (CS, LSA1, LSA3, STARP, EXP1, pb9) | IM | I | Simian adenovirus ChAd63, MVA |
| PfSPZ | ND | DVI | ND | Simian adenovirus ChAd63, MVA |
| PfCelTOS FMP012 | CelTOS (cell-traversal protein for ookinetes and sporozoites) | IM | Ia | E. coli, B834 |
| CSVAC | CS | IM | I | Adenovirus ChAd63 and MVA |
| R21/AS01B | CSP | ND | Ia | ND |
| R21/Matrix-M1 | CSP | ND | Ib | ND |
| R21 (RTS,S-biosimilar)/ME-TRAP | CSP | SC, ID, IM | IIa | P. pastoris |
| Blood stage | ||||
| GMZ2 | GLURP, MSP3 | IM | II | L. lactis |
| PfAMA1-DiCo | AMA-1-DiCo | IM | I | P. pastoris |
| P27A | P27A | IM | ND | Synthetic peptide |
| MSP3[181–276] | MSP3 | SC | IIb | Synthetic peptide |
| SE36 | N-terminal of serine repeat antigen (SERA5) | SC, IM | Ib | E. coli |
| PfPEBS | ND | ND | II | E. coli |
| ChAd63 RH5 +/− MVA RH5 | RH | IM | Ia | ChAd63 and MVA |
| PRIMVAC | VAR2CSA | IM | Ia/b | E. coli |
| PAMVAC | VAR2CSA | IM | Ia/b | Drosophila S2 cells |
| Sexual stage | ||||
| Pfs25 VLP | Pfs25 | IM | I/IIa | N. benthamiana |
| Pfs25-EPA/Alhydrogel | Pfs25 | IM | Ib | P. pastoris |
| Pfs230D1M-EPA/Alhydrogel and/or Pfs25-EPA/Alhydrogel | Pfs25M, Pfs230D1M | IM | I | P. pastoris |
| Pfs230D1M-EPA/Alhydrogel and Pfs25-EPA/AS01 | Pfs25M, Pfs230D1M | IM | I | P. pastoris |
| ChAd63 Pfs25-IMX313/MVA Pfs25-IMX313 | Pfs25 | IM | Ia | Chimpanzee Adenovirus 63, MVA |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Skwarczynski, M.; Chandrudu, S.; Rigau-Planella, B.; Islam, M.T.; Cheong, Y.S.; Liu, G.; Wang, X.; Toth, I.; Hussein, W.M. Progress in the Development of Subunit Vaccines against Malaria. Vaccines 2020, 8, 373. https://doi.org/10.3390/vaccines8030373
Skwarczynski M, Chandrudu S, Rigau-Planella B, Islam MT, Cheong YS, Liu G, Wang X, Toth I, Hussein WM. Progress in the Development of Subunit Vaccines against Malaria. Vaccines. 2020; 8(3):373. https://doi.org/10.3390/vaccines8030373
Chicago/Turabian StyleSkwarczynski, Mariusz, Saranya Chandrudu, Berta Rigau-Planella, Md. Tanjir Islam, Yee S. Cheong, Genan Liu, Xiumin Wang, Istvan Toth, and Waleed M. Hussein. 2020. "Progress in the Development of Subunit Vaccines against Malaria" Vaccines 8, no. 3: 373. https://doi.org/10.3390/vaccines8030373
APA StyleSkwarczynski, M., Chandrudu, S., Rigau-Planella, B., Islam, M. T., Cheong, Y. S., Liu, G., Wang, X., Toth, I., & Hussein, W. M. (2020). Progress in the Development of Subunit Vaccines against Malaria. Vaccines, 8(3), 373. https://doi.org/10.3390/vaccines8030373