Vaccines against Genital Herpes: Where Are We?


Abstract
:1. Introduction
2. Immune Response against HSV
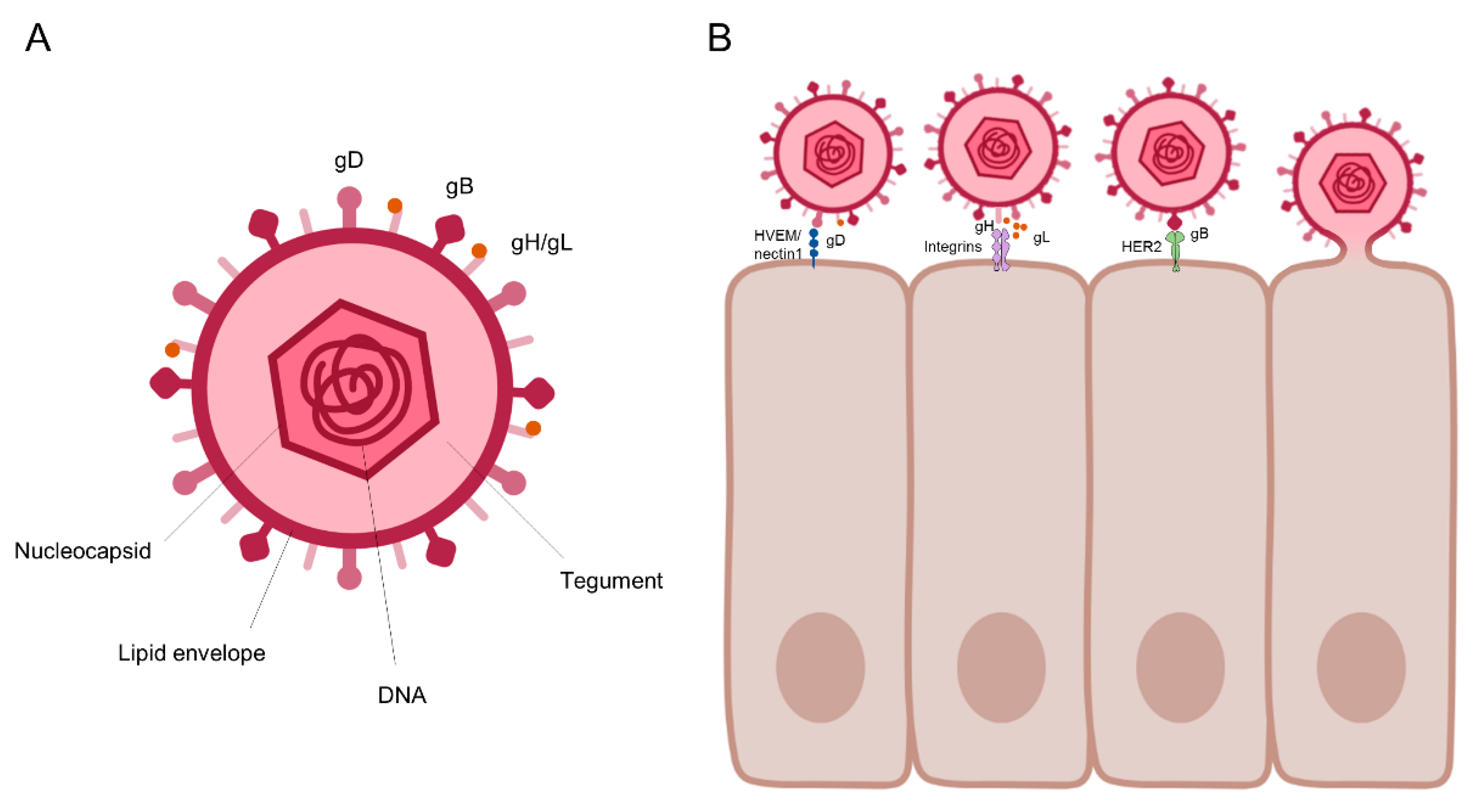
2.1. The HSV Life Cycle
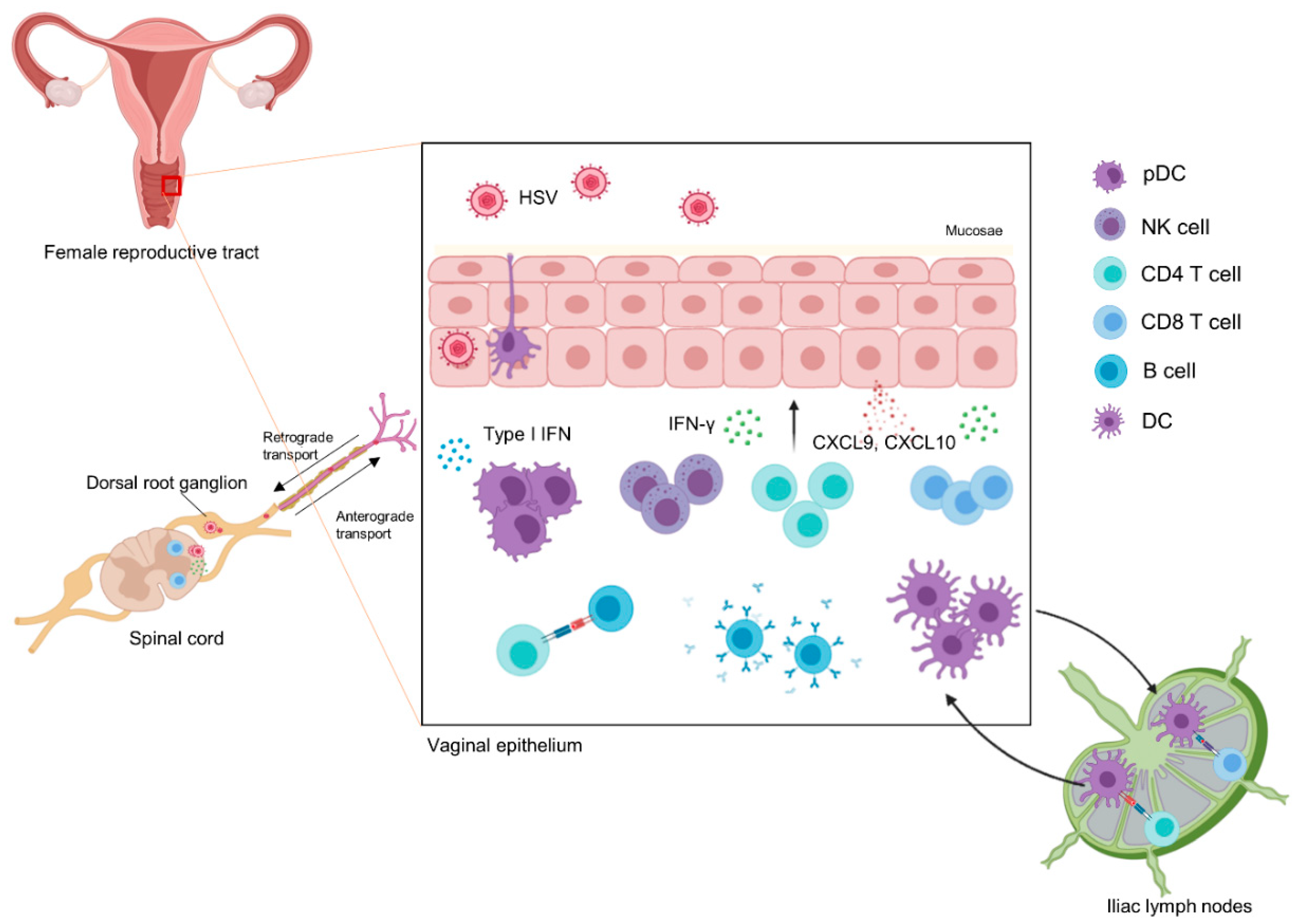
2.2. The Anti-HSV IMMUNE Response
3. Status of HSV Vaccine Development
3.1. Overview
3.2. Recent Progress
4. Limitations & Future Directions
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Gupta, R.; Wald, A.; Krantz, E.; Selke, S.; Warren, T.; Vargas-Cortes, M.; Miller, G.; Corey, L. Valacyclovir and acyclovir for suppression of shedding of herpes simplex virus in the genital tract. J. Infect. Dis. 2004, 190, 1374–1381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corey, L.; Wald, A.; Patel, R.; Sacks, S.L.; Tyring, S.K.; Warren, T.; Douglas, J.M., Jr.; Paavonen, J.; Morrow, R.A.; Beutner, K.R.; et al. Once-daily valacyclovir to reduce the risk of transmission of genital herpes. N. Engl. J. Med. 2004, 350, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Martens, M.G.; Fife, K.H.; Leone, P.A.; Dix, L.P.; Brennan, C.A. Once daily valacyclovir for reducing viral shedding in subjects newly diagnosed with genital herpes. Infect. Dis. Obstet. Gynecol. 2009, 2009, 105376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hofstetter, A.M.; Rosenthal, S.L.; Stanberry, L.R. Current thinking on genital herpes. Curr. Opin. Infect. Dis. 2014, 27, 75–83. [Google Scholar] [CrossRef]
- Bernstein, D.I.; Bellamy, A.R.; Hook, E.W., III; Levin, M.J.; Wald, A.; Ewell, M.G.; Wolff, P.A.; Deal, C.D.; Heineman, T.C.; Dubin, G.; et al. Epidemiology, clinical presentation, and antibody response to primary infection with herpes simplex virus type 1 and type 2 in young women. Clin. Infect. Dis. 2013, 56, 344–351. [Google Scholar] [CrossRef]
- Looker, K.J.; Garnett, G.P.; Schmid, G.P. An estimate of the global prevalence and incidence of herpes simplex virus type 2 infection. Bull. World Health Organ. 2008, 86, 805–812. [Google Scholar] [CrossRef]
- Whitley, R.J.; Nahmias, A.J.; Soong, S.J.; Galasso, G.G.; Fleming, C.L.; Alford, C.A. Vidarabine therapy of neonatal herpes simplex virus infection. Pediatrics 1980, 66, 495–501. [Google Scholar]
- Kimberlin, D.W. Neonatal herpes simplex infection. Clin. Microbiol. Rev. 2004, 17, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Morris, S.R.; Bauer, H.M.; Samuel, M.C.; Gallagher, D.; Bolan, G. Neonatal herpes morbidity and mortality in California, 1995–2003. Sex. Transm. Dis. 2008, 35, 14–18. [Google Scholar] [CrossRef]
- Ryder, N.; Jin, F.; McNulty, A.M.; Grulich, A.E.; Donovan, B. Increasing role of herpes simplex virus type 1 in first-episode anogenital herpes in heterosexual women and younger men who have sex with men, 1992–2006. Sex. Transm. Infect. 2009, 85, 416–419. [Google Scholar] [CrossRef]
- Flagg, E.W.; Weinstock, H. Incidence of neonatal herpes simplex virus infections in the United States, 2006. Pediatrics 2011, 127, e1–e8. [Google Scholar] [CrossRef] [PubMed]
- Prober, C.G.; Sullender, W.M.; Yasukawa, L.L.; Au, D.S.; Yeager, A.S.; Arvin, A.M. Low risk of herpes simplex virus infections in neonates exposed to the virus at the time of vaginal delivery to mothers with recurrent genital herpes simplex virus infections. N. Engl. J. Med. 1987, 316, 240–244. [Google Scholar] [CrossRef] [PubMed]
- Freeman, E.E.; Orroth, K.K.; White, R.G.; Glynn, J.R.; Bakker, R.; Boily, M.C.; Habbema, D.; Buve, A.; Hayes, R. Proportion of new HIV infections attributable to herpes simplex 2 increases over time: Simulations of the changing role of sexually transmitted infections in sub-Saharan African HIV epidemics. Sex. Transm. Infect. 2007, 83, i17–i24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masese, L.; Baeten, J.M.; Richardson, B.A.; Bukusi, E.; John-Stewart, G.; Graham, S.M.; Shafi, J.; Kiarie, J.; Overbaugh, J.; McClelland, R.S. Changes in the contribution of genital tract infections to HIV acquisition among Kenyan high-risk women from 1993 to 2012. AIDS 2015, 29, 1077–1085. [Google Scholar] [CrossRef] [PubMed]
- Owusu-Edusei, K., Jr.; Chesson, H.W.; Gift, T.L.; Tao, G.; Mahajan, R.; Ocfemia, M.C.; Kent, C.K. The estimated direct medical cost of selected sexually transmitted infections in the United States, 2008. Sex. Transm. Dis. 2013, 40, 197–201. [Google Scholar] [CrossRef] [PubMed]
- Martin, E.T.; Krantz, E.; Gottlieb, S.L.; Magaret, A.S.; Langenberg, A.; Stanberry, L.; Kamb, M.; Wald, A. A pooled analysis of the effect of condoms in preventing HSV-2 acquisition. Arch. Intern. Med. 2009, 169, 1233–1240. [Google Scholar] [CrossRef] [Green Version]
- Lingappa, J.; Nakku-Joloba, E.; Magaret, A.; Friedrich, D.; Dragavon, J.; Kambugu, F.; Joloba, M.; Whalen, C.; Coombs, R.; Celum, C.; et al. Sensitivity and specificity of herpes simplex virus-2 serological assays among HIV-infected and uninfected urban Ugandans. Int. J. STD AIDS 2010, 21, 611–616. [Google Scholar] [CrossRef] [Green Version]
- Campadelli-Fiume, G.; Menotti, L.; Avitabile, E.; Gianni, T. Viral and cellular contributions to herpes simplex virus entry into the cell. Curr. Opin. Virol. 2012, 2, 28–36. [Google Scholar] [CrossRef]
- Knipe, D.M.; Cliffe, A. Chromatin control of herpes simplex virus lytic and latent infection. Nat. Rev. Microbiol. 2008, 6, 211–221. [Google Scholar] [CrossRef]
- Smith, G. Herpesvirus transport to the nervous system and back again. Annu. Rev. Microbiol. 2012, 66, 153–176. [Google Scholar] [CrossRef] [Green Version]
- Spruance, S.L. Cutaneous herpes simplex virus lesions induced by ultraviolet radiation. A review of model systems and prophylactic therapy with oral acyclovir. Am. J. Med. 1988, 85, 43–45. [Google Scholar] [PubMed]
- Chida, Y.; Mao, X. Does psychosocial stress predict symptomatic herpes simplex virus recurrence? A meta-analytic investigation on prospective studies. Brain Behav. Immun. 2009, 23, 917–925. [Google Scholar] [CrossRef] [PubMed]
- El Hayderi, L.; Delvenne, P.; Rompen, E.; Senterre, J.M.; Nikkels, A.F. Herpes simplex virus reactivation and dental procedures. Clin. Oral Investig. 2013, 17, 1961–1964. [Google Scholar] [CrossRef] [PubMed]
- Wilson, A.C.; Mohr, I. A cultured affair: HSV latency and reactivation in neurons. Trends Microbiol. 2012, 20, 604–611. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kinnebrew, M.A.; Buffie, C.G.; Diehl, G.E.; Zenewicz, L.A.; Leiner, I.; Hohl, T.M.; Flavell, R.A.; Littman, D.R.; Pamer, E.G. Interleukin 23 production by intestinal CD103(+)CD11b(+) dendritic cells in response to bacterial flagellin enhances mucosal innate immune defense. Immunity 2012, 36, 276–287. [Google Scholar] [CrossRef] [Green Version]
- Medzhitov, R.; Janeway, C., Jr. Innate immune recognition: Mechanisms and pathways. Immunol. Rev. 2000, 173, 89–97. [Google Scholar] [CrossRef]
- Everett, R.D.; Rechter, S.; Papior, P.; Tavalai, N.; Stamminger, T.; Orr, A. PML contributes to a cellular mechanism of repression of herpes simplex virus type 1 infection that is inactivated by ICP0. J. Virol. 2006, 80, 7995–8005. [Google Scholar] [CrossRef] [Green Version]
- Everett, R.D.; Parada, C.; Gripon, P.; Sirma, H.; Orr, A. Replication of ICP0-null mutant herpes simplex virus type 1 is restricted by both PML and Sp100. J. Virol. 2008, 82, 2661–2672. [Google Scholar] [CrossRef] [Green Version]
- Brown, J.R.; Conn, K.L.; Wasson, P.; Charman, M.; Tong, L.; Grant, K.; McFarlane, S.; Boutell, C. SUMO Ligase Protein Inhibitor of Activated STAT1 (PIAS1) Is a Constituent Promyelocytic Leukemia Nuclear Body Protein That Contributes to the Intrinsic Antiviral Immune Response to Herpes Simplex Virus 1. J. Virol. 2016, 90, 5939–5952. [Google Scholar] [CrossRef] [Green Version]
- Conn, K.L.; Wasson, P.; McFarlane, S.; Tong, L.; Brown, J.R.; Grant, K.G.; Domingues, P.; Boutell, C. Novel Role for Protein Inhibitor of Activated STAT 4 (PIAS4) in the Restriction of Herpes Simplex Virus 1 by the Cellular Intrinsic Antiviral Immune Response. J. Virol. 2016, 90, 4807–4826. [Google Scholar] [CrossRef] [Green Version]
- Kim, E.T.; White, T.E.; Brandariz-Nunez, A.; Diaz-Griffero, F.; Weitzman, M.D. SAMHD1 restricts herpes simplex virus 1 in macrophages by limiting DNA replication. J. Virol. 2013, 87, 12949–12956. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Luo, S.; He, S.; Zhang, M.; Wang, P.; Li, C.; Huang, W.; Hu, B.; Griffin, G.E.; Shattock, R.J.; et al. Tetherin restricts HSV-2 release and is counteracted by multiple viral glycoproteins. Virology 2015, 475, 96–109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lilley, C.E.; Chaurushiya, M.S.; Boutell, C.; Landry, S.; Suh, J.; Panier, S.; Everett, R.D.; Stewart, G.S.; Durocher, D.; Weitzman, M.D. A viral E3 ligase targets RNF8 and RNF168 to control histone ubiquitination and DNA damage responses. EMBO J. 2010, 29, 943–955. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lilley, C.E.; Chaurushiya, M.S.; Boutell, C.; Everett, R.D.; Weitzman, M.D. The intrinsic antiviral defense to incoming HSV-1 genomes includes specific DNA repair proteins and is counteracted by the viral protein ICP0. PLoS Pathog. 2011, 7, e1002084. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maul, G.G.; Everett, R.D. The nuclear location of PML, a cellular member of the C3HC4 zinc-binding domain protein family, is rearranged during herpes simplex virus infection by the C3HC4 viral protein ICP0. J. Gen. Virol. 1994, 75, 1223–1233. [Google Scholar] [CrossRef]
- Zhang, K.; Lv, D.W.; Li, R. Conserved Herpesvirus Protein Kinases Target SAMHD1 to Facilitate Virus Replication. Cell Rep. 2019, 28, 449–459.e5. [Google Scholar] [CrossRef] [Green Version]
- Sainz, B., Jr.; Halford, W.P. Alpha/Beta interferon and gamma interferon synergize to inhibit the replication of herpes simplex virus type 1. J. Virol. 2002, 76, 11541–11550. [Google Scholar] [CrossRef] [Green Version]
- Leventon-Kriss, S.; Movshovitz, M.; Smetana, Z.; Shewach-Millet, M.; Doerner, T.; Gotlieb-Stematsky, T. Sensitivity in vitro of herpes simplex virus isolates to human fibroblast interferon. Med. Microbiol. Immunol. 1987, 176, 151–159. [Google Scholar] [CrossRef]
- Sancho-Shimizu, V.; Perez de Diego, R.; Lorenzo, L.; Halwani, R.; Alangari, A.; Israelsson, E.; Fabrega, S.; Cardon, A.; Maluenda, J.; Tatematsu, M.; et al. Herpes simplex encephalitis in children with autosomal recessive and dominant TRIF deficiency. J. Clin. Investig. 2011, 121, 4889–4902. [Google Scholar] [CrossRef] [Green Version]
- Dropulic, L.K.; Cohen, J.I. Severe viral infections and primary immunodeficiencies. Clin. Infect. Dis. 2011, 53, 897–909. [Google Scholar] [CrossRef]
- Cunningham, A.L.; Turner, R.R.; Miller, A.C.; Para, M.F.; Merigan, T.C. Evolution of recurrent herpes simplex lesions. An immunohistologic study. J. Clin. Investig. 1985, 75, 226–233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koelle, D.M.; Posavad, C.M.; Barnum, G.R.; Johnson, M.L.; Frank, J.M.; Corey, L. Clearance of HSV-2 from recurrent genital lesions correlates with infiltration of HSV-specific cytotoxic T lymphocytes. J. Clin. Investig. 1998, 101, 1500–1508. [Google Scholar] [CrossRef] [PubMed]
- Iwasaki, A. Mucosal dendritic cells. Annu. Rev. Immunol. 2007, 25, 381–418. [Google Scholar] [CrossRef] [PubMed]
- Nakanishi, Y.; Lu, B.; Gerard, C.; Iwasaki, A. CD8(+) T lymphocyte mobilization to virus-infected tissue requires CD4(+) T-cell help. Nature 2009, 462, 510–513. [Google Scholar] [CrossRef] [Green Version]
- Wong, S.B.; Bos, R.; Sherman, L.A. Tumor-specific CD4+ T cells render the tumor environment permissive for infiltration by low-avidity CD8+ T cells. J. Immunol. 2008, 180, 3122–3131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swain, S.L.; McKinstry, K.K.; Strutt, T.M. Expanding roles for CD4(+) T cells in immunity to viruses. Nat. Rev. Immunol. 2012, 12, 136–148. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.C.; Oh, D.S.; Park, J.H.; Kim, H.J.; Seo, Y.B.; Yoo, H.J.; Jang, H.S.; Shin, J.; Kim, C.W.; Kwon, M.S.; et al. Multivalent DNA vaccine protects against genital herpes by T-cell immune induction in vaginal mucosa. Antivir. Res. 2020, 177, 104755. [Google Scholar] [CrossRef]
- Hoshino, Y.; Pesnicak, L.; Cohen, J.I.; Straus, S.E. Rates of reactivation of latent herpes simplex virus from mouse trigeminal ganglia ex vivo correlate directly with viral load and inversely with number of infiltrating CD8+ T cells. J. Virol. 2007, 81, 8157–8164. [Google Scholar] [CrossRef] [Green Version]
- Simmons, A.; Tscharke, D.C. Anti-CD8 impairs clearance of herpes simplex virus from the nervous system: Implications for the fate of virally infected neurons. J. Exp. Med. 1992, 175, 1337–1344. [Google Scholar] [CrossRef] [Green Version]
- Iijima, N.; Linehan, M.M.; Zamora, M.; Butkus, D.; Dunn, R.; Kehry, M.R.; Laufer, T.M.; Iwasaki, A. Dendritic cells and B cells maximize mucosal Th1 memory response to herpes simplex virus. J. Exp. Med. 2008, 205, 3041–3052. [Google Scholar] [CrossRef]
- Sajic, D.; Patrick, A.J.; Rosenthal, K.L. Mucosal delivery of CpG oligodeoxynucleotides expands functional dendritic cells and macrophages in the vagina. Immunology 2005, 114, 213–224. [Google Scholar] [CrossRef] [PubMed]
- Schiffer, J.T.; Abu-Raddad, L.; Mark, K.E.; Zhu, J.; Selke, S.; Koelle, D.M.; Wald, A.; Corey, L. Mucosal host immune response predicts the severity and duration of herpes simplex virus-2 genital tract shedding episodes. Proc. Natl. Acad. Sci. USA 2010, 107, 18973–18978. [Google Scholar] [CrossRef] [Green Version]
- Zhu, J.; Koelle, D.M.; Cao, J.; Vazquez, J.; Huang, M.L.; Hladik, F.; Wald, A.; Corey, L. Virus-specific CD8+ T cells accumulate near sensory nerve endings in genital skin during subclinical HSV-2 reactivation. J. Exp. Med. 2007, 204, 595–603. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schiffer, J.T.; Abu-Raddad, L.; Mark, K.E.; Zhu, J.; Selke, S.; Magaret, A.; Wald, A.; Corey, L. Frequent release of low amounts of herpes simplex virus from neurons: Results of a mathematical model. Sci. Transl. Med. 2009, 1, 7ra16. [Google Scholar] [CrossRef] [Green Version]
- Khanna, K.M.; Bonneau, R.H.; Kinchington, P.R.; Hendricks, R.L. Herpes simplex virus-specific memory CD8+ T cells are selectively activated and retained in latently infected sensory ganglia. Immunity 2003, 18, 593–603. [Google Scholar] [CrossRef] [Green Version]
- Skinner, G.R.; Turyk, M.E.; Benson, C.A.; Wilbanks, G.D.; Heseltine, P.; Galpin, J.; Kaufman, R.; Goldberg, L.; Hartley, C.E.; Buchan, A. The efficacy and safety of Skinner herpes simplex vaccine towards modulation of herpes genitalis; report of a prospective double-blind placebo-controlled trial. Med. Microbiol. Immunol. 1997, 186, 31–36. [Google Scholar] [CrossRef]
- Aurelian, L.; Kokuba, H.; Smith, C.C. Vaccine potential of a herpes simplex virus type 2 mutant deleted in the PK domain of the large subunit of ribonucleotide reductase (ICP10). Vaccine 1999, 17, 1951–1963. [Google Scholar] [CrossRef]
- Casanova, G.; Cancela, R.; Alonzo, L.; Benuto, R.; Magana Mdel, C.; Hurley, D.R.; Fishbein, E.; Lara, C.; Gonzalez, T.; Ponce, R.; et al. A double-blind study of the efficacy and safety of the ICP10deltaPK vaccine against recurrent genital HSV-2 infections. Cutis 2002, 70, 235–239. [Google Scholar]
- Straus, S.E.; Wald, A.; Kost, R.G.; McKenzie, R.; Langenberg, A.G.; Hohman, P.; Lekstrom, J.; Cox, E.; Nakamura, M.; Sekulovich, R.; et al. Immunotherapy of recurrent genital herpes with recombinant herpes simplex virus type 2 glycoproteins D and B: Results of a placebo-controlled vaccine trial. J. Infect. Dis. 1997, 176, 1129–1134. [Google Scholar] [CrossRef]
- Corey, L.; Langenberg, A.G.; Ashley, R.; Sekulovich, R.E.; Izu, A.E.; Douglas, J.M., Jr.; Handsfield, H.H.; Warren, T.; Marr, L.; Tyring, S.; et al. Recombinant glycoprotein vaccine for the prevention of genital HSV-2 infection: Two randomized controlled trials. Chiron HSV Vaccine Study Group. JAMA 1999, 282, 331–340. [Google Scholar] [CrossRef]
- Stanberry, L.R.; Spruance, S.L.; Cunningham, A.L.; Bernstein, D.I.; Mindel, A.; Sacks, S.; Tyring, S.; Aoki, F.Y.; Slaoui, M.; Denis, M.; et al. Glycoprotein-D-adjuvant vaccine to prevent genital herpes. N. Engl. J. Med. 2002, 347, 1652–1661. [Google Scholar] [CrossRef] [PubMed]
- Belshe, R.B.; Leone, P.A.; Bernstein, D.I.; Wald, A.; Levin, M.J.; Stapleton, J.T.; Gorfinkel, I.; Morrow, R.L.; Ewell, M.G.; Stokes-Riner, A.; et al. Efficacy results of a trial of a herpes simplex vaccine. N. Engl. J. Med. 2012, 366, 34–43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cohen, J. Immunology. Painful failure of promising genital herpes vaccine. Science 2010, 330, 304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bernstein, D.I.; Morello, C.S.; Cardin, R.D.; Bravo, F.J.; Kraynyak, K.A.; Spector, D.H. A vaccine containing highly purified virus particles in adjuvant provides high level protection against genital infection and disease in guinea pigs challenged intravaginally with homologous and heterologous strains of herpes simplex virus type 2. Vaccine 2020, 38, 79–89. [Google Scholar] [CrossRef] [PubMed]
- Agelidis, A.; Koujah, L.; Suryawanshi, R.; Yadavalli, T.; Mishra, Y.K.; Adelung, R.; Shukla, D. An Intra-Vaginal Zinc Oxide Tetrapod Nanoparticles (ZOTEN) and Genital Herpesvirus Cocktail Can Provide a Novel Platform for Live Virus Vaccine. Front. Immunol. 2019, 10, 500. [Google Scholar] [CrossRef] [Green Version]
- Antoine, T.E.; Mishra, Y.K.; Trigilio, J.; Tiwari, V.; Adelung, R.; Shukla, D. Prophylactic, therapeutic and neutralizing effects of zinc oxide tetrapod structures against herpes simplex virus type-2 infection. Antivir. Res. 2012, 96, 363–375. [Google Scholar] [CrossRef] [Green Version]
- Hosken, N.; McGowan, P.; Meier, A.; Koelle, D.M.; Sleath, P.; Wagener, F.; Elliott, M.; Grabstein, K.; Posavad, C.; Corey, L. Diversity of the CD8 + T-cell response to herpes simplex virus type 2 proteins among persons with genital herpes. J. Virol. 2006, 80, 5509–5515. [Google Scholar] [CrossRef] [Green Version]
- Laing, K.J.; Magaret, A.S.; Mueller, D.E.; Zhao, L.; Johnston, C.; De Rosa, S.C.; Koelle, D.M.; Wald, A.; Corey, L. Diversity in CD8 (+) T cell function and epitope breadth among persons with genital herpes. J. Clin. Immunol. 2010, 30, 703–722. [Google Scholar] [CrossRef] [Green Version]
- Koelle, D.M.; Liu, Z.; McClurkan, C.L.; Cevallos, R.C.; Vieira, J.; Hosken, N.A.; Meseda, C.A.; Snow, D.C.; Wald, A.; Corey, L. Immunodominance among herpes simplex virus-specific CD8 T cells expressing a tissue-specific homing receptor. Proc. Natl. Acad. Sci. USA 2003, 100, 12899–12904. [Google Scholar] [CrossRef] [Green Version]
- Koelle, D.M.; Chen, H.B.; Gavin, M.A.; Wald, A.; Kwok, W.W.; Corey, L. CD8 CTL from genital herpes simplex lesions: Recognition of viral tegument and immediate early proteins and lysis of infected cutaneous cells. J. Immunol. 2001, 166, 4049–4058. [Google Scholar] [CrossRef]
- Koelle, D.M.; Frank, J.M.; Johnson, M.L.; Kwok, W.W. Recognition of herpes simplex virus type 2 tegument proteins by CD4 T cells infiltrating human genital herpes lesions. J. Virol. 1998, 72, 7476–7483. [Google Scholar] [CrossRef] [Green Version]
- Iyer, A.V.; Pahar, B.; Chouljenko, V.N.; Walker, J.D.; Stanfield, B.; Kousoulas, K.G. Single dose of glycoprotein K (gK)-deleted HSV-1 live-attenuated virus protects mice against lethal vaginal challenge with HSV-1 and HSV-2 and induces lasting T cell memory immune responses. Virol. J. 2013, 10, 317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stanfield, B.A.; Rider, P.J.F.; Caskey, J.; Del Piero, F.; Kousoulas, K.G. Intramuscular vaccination of guinea pigs with the live-attenuated human herpes simplex vaccine VC2 stimulates a transcriptional profile of vaginal Th17 and regulatory Tr1 responses. Vaccine 2018, 36, 2842–2849. [Google Scholar] [CrossRef] [PubMed]
- Bernstein, D.I.; Pullum, D.A.; Cardin, R.D.; Bravo, F.J.; Dixon, D.A.; Kousoulas, K.G. The HSV-1 live attenuated VC2 vaccine provides protection against HSV-2 genital infection in the guinea pig model of genital herpes. Vaccine 2019, 37, 61–68. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Wang, Z.; Xu, Y.; Zhang, Z.; Hua, R.; Liu, W.; Jiang, C.; Chen, Y.; Yang, W.; Kong, W. Optimized DNA Vaccine Enhanced by Adjuvant IL28B Induces Protective Immune Responses Against Herpes Simplex Virus Type 2 in Mice. Viral Immunol. 2017, 30, 601–614. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Zhou, Y.; Wang, Z.; Zhang, Z.; Wang, Q.; Su, W.; Chen, Y.; Zhang, Y.; Gao, F.; Jiang, C.; et al. Evaluation of recombinant adenovirus vaccines based on glycoprotein D and truncated UL25 against herpes simplex virus type 2 in mice. Microbiol. Immunol. 2017, 61, 176–184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ogasawara, M.; Suzutani, T.; Yoshida, I.; Azuma, M. Role of the UL25 gene product in packaging DNA into the herpes simplex virus capsid: Location of UL25 product in the capsid and demonstration that it binds DNA. J. Virol. 2001, 75, 1427–1436. [Google Scholar] [CrossRef] [Green Version]
- Su, H.K.; Eberle, R.; Courtney, R.J. Processing of the herpes simplex virus type 2 glycoprotein gG-2 results in secretion of a 34,000-Mr cleavage product. J. Virol. 1987, 61, 1735–1737. [Google Scholar] [CrossRef] [Green Version]
- Balachandran, N.; Hutt-Fletcher, L.M. Synthesis and processing of glycoprotein gG of herpes simplex virus type 2. J. Virol. 1985, 54, 825–832. [Google Scholar] [CrossRef] [Green Version]
- Olofsson, S.; Lundstrom, M.; Marsden, H.; Jeansson, S.; Vahlne, A. Characterization of a herpes simplex virus type 2-specified glycoprotein with affinity for N-acetylgalactosamine-specific lectins and its identification as g92K or gG. J. Gen. Virol. 1986, 67, 737–744. [Google Scholar] [CrossRef]
- Viejo-Borbolla, A.; Martinez-Martin, N.; Nel, H.J.; Rueda, P.; Martin, R.; Blanco, S.; Arenzana-Seisdedos, F.; Thelen, M.; Fallon, P.G.; Alcami, A. Enhancement of chemokine function as an immunomodulatory strategy employed by human herpesviruses. PLoS Pathog. 2012, 8, e1002497. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martinez-Martin, N.; Viejo-Borbolla, A.; Martin, R.; Blanco, S.; Benovic, J.L.; Thelen, M.; Alcami, A. Herpes simplex virus enhances chemokine function through modulation of receptor trafficking and oligomerization. Nat. Commun. 2015, 6, 6163. [Google Scholar] [CrossRef] [Green Version]
- Onnheim, K.; Ekblad, M.; Gorander, S.; Bergstrom, T.; Liljeqvist, J.A. Vaccination with the Secreted Glycoprotein G of Herpes Simplex Virus 2 Induces Protective Immunity after Genital Infection. Viruses 2016, 8, 110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Awasthi, S.; Lubinski, J.M.; Shaw, C.E.; Barrett, S.M.; Cai, M.; Wang, F.; Betts, M.; Kingsley, S.; Distefano, D.J.; Balliet, J.W.; et al. Immunization with a vaccine combining herpes simplex virus 2 (HSV-2) glycoprotein C (gC) and gD subunits improves the protection of dorsal root ganglia in mice and reduces the frequency of recurrent vaginal shedding of HSV-2 DNA in guinea pigs compared to immunization with gD alone. J. Virol. 2011, 85, 10472–10486. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Awasthi, S.; Huang, J.; Shaw, C.; Friedman, H.M. Blocking herpes simplex virus 2 glycoprotein E immune evasion as an approach to enhance efficacy of a trivalent subunit antigen vaccine for genital herpes. J. Virol. 2014, 88, 8421–8432. [Google Scholar] [CrossRef] [Green Version]
- Awasthi, S.; Hook, L.M.; Shaw, C.E.; Pahar, B.; Stagray, J.A.; Liu, D.; Veazey, R.S.; Friedman, H.M. An HSV-2 Trivalent Vaccine Is Immunogenic in Rhesus Macaques and Highly Efficacious in Guinea Pigs. PLoS Pathog. 2017, 13, e1006141. [Google Scholar] [CrossRef] [Green Version]
- Awasthi, S.; Hook, L.M.; Shaw, C.E.; Friedman, H.M. A trivalent subunit antigen glycoprotein vaccine as immunotherapy for genital herpes in the guinea pig genital infection model. Hum. Vaccin Immunother. 2017, 13, 2785–2793. [Google Scholar] [CrossRef]
- Egan, K.; Hook, L.M.; Naughton, A.; Friedman, H.M.; Awasthi, S. Herpes simplex virus type 2 trivalent protein vaccine containing glycoproteins C, D and E protects guinea pigs against HSV-1 genital infection. Hum. Vaccin Immunother. 2020, 1–5. [Google Scholar] [CrossRef]
- Hook, L.M.; Awasthi, S.; Dubin, J.; Flechtner, J.; Long, D.; Friedman, H.M. A trivalent gC2/gD2/gE2 vaccine for herpes simplex virus generates antibody responses that block immune evasion domains on gC2 better than natural infection. Vaccine 2019, 37, 664–669. [Google Scholar] [CrossRef]
- Awasthi, S.; Hook, L.M.; Pardi, N.; Wang, F.; Myles, A.; Cancro, M.P.; Cohen, G.H.; Weissman, D.; Friedman, H.M. Nucleoside-modified mRNA encoding HSV-2 glycoproteins C, D, and E prevents clinical and subclinical genital herpes. Sci. Immunol. 2019, 4. [Google Scholar] [CrossRef]
- Kollias, C.M.; Huneke, R.B.; Wigdahl, B.; Jennings, S.R. Animal models of herpes simplex virus immunity and pathogenesis. J. Neurovirol. 2015, 21, 8–23. [Google Scholar] [CrossRef]
- Sawtell, N.M.; Thompson, R.L. Herpes simplex virus type 1 latency-associated transcription unit promotes anatomical site-dependent establishment and reactivation from latency. J. Virol. 1992, 66, 2157–2169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hensel, M.T.; Marshall, J.D.; Dorwart, M.R.; Heeke, D.S.; Rao, E.; Tummala, P.; Yu, L.; Cohen, G.H.; Eisenberg, R.J.; Sloan, D.D. Prophylactic Herpes Simplex Virus 2 (HSV-2) Vaccines Adjuvanted with Stable Emulsion and Toll-Like Receptor 9 Agonist Induce a Robust HSV-2-Specific Cell-Mediated Immune Response, Protect against Symptomatic Disease, and Reduce the Latent Viral Reservoir. J. Virol. 2017, 91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bernstein, D.I.; Cardin, R.D.; Bravo, F.J.; Awasthi, S.; Lu, P.; Pullum, D.A.; Dixon, D.A.; Iwasaki, A.; Friedman, H.M. Successful application of prime and pull strategy for a therapeutic HSV vaccine. NPJ Vaccines 2019, 4, 33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stanberry, L.R.; Kern, E.R.; Richards, J.T.; Abbott, T.M.; Overall, J.C., Jr. Genital herpes in guinea pigs: Pathogenesis of the primary infection and description of recurrent disease. J. Infect. Dis. 1982, 146, 397–404. [Google Scholar] [CrossRef] [PubMed]
- Stanberry, L.R.; Kern, E.R.; Richards, J.T.; Overall, J.C., Jr. Recurrent genital herpes simplex virus infection in guinea pigs. Intervirology 1985, 24, 226–231. [Google Scholar] [CrossRef]
- Yim, K.C.; Carroll, C.J.; Tuyama, A.; Cheshenko, N.; Carlucci, M.J.; Porter, D.D.; Prince, G.A.; Herold, B.C. The cotton rat provides a novel model to study genital herpes infection and to evaluate preventive strategies. J. Virol. 2005, 79, 14632–14639. [Google Scholar] [CrossRef] [Green Version]
- Crostarosa, F.; Aravantinou, M.; Akpogheneta, O.J.; Jasny, E.; Shaw, A.; Kenney, J.; Piatak, M.; Lifson, J.D.; Teitelbaum, A.; Hu, L.; et al. A macaque model to study vaginal HSV-2/immunodeficiency virus co-infection and the impact of HSV-2 on microbicide efficacy. PLoS ONE 2009, 4, e8060. [Google Scholar] [CrossRef] [Green Version]
- Melendez, L.V.; Espana, C.; Hunt, R.D.; Daniel, M.D.; Garcia, F.G. Natural herpes simplex infection in the owl monkey (Aotus trivirgatus). Lab. Anim. Care 1969, 19, 38–45. [Google Scholar]
- Edwards, A.D.; Diebold, S.S.; Slack, E.M.; Tomizawa, H.; Hemmi, H.; Kaisho, T.; Akira, S.; Reis e Sousa, C. Toll-like receptor expression in murine DC subsets: Lack of TLR7 expression by CD8 alpha+ DC correlates with unresponsiveness to imidazoquinolines. Eur. J. Immunol. 2003, 33, 827–833. [Google Scholar] [CrossRef]
- Jongbloed, S.L.; Kassianos, A.J.; McDonald, K.J.; Clark, G.J.; Ju, X.; Angel, C.E.; Chen, C.J.; Dunbar, P.R.; Wadley, R.B.; Jeet, V.; et al. Human CD141+ (BDCA-3)+ dendritic cells (DCs) represent a unique myeloid DC subset that cross-presents necrotic cell antigens. J. Exp. Med. 2010, 207, 1247–1260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wald, A.; Koelle, D.M.; Fife, K.; Warren, T.; Leclair, K.; Chicz, R.M.; Monks, S.; Levey, D.L.; Musselli, C.; Srivastava, P.K. Safety and immunogenicity of long HSV-2 peptides complexed with rhHsc70 in HSV-2 seropositive persons. Vaccine 2011, 29, 8520–8529. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Classification | Candidate | Company | Composition | Status | Phase | Identifier 1 | Completion |
|---|---|---|---|---|---|---|---|
| Subunit vaccine | gD2 | Novartis (previous Chiron) | gD2 plus alum | Stopped after Phase II trial | Phase I | - | 1992 |
| gD2/gB2 | gD2 and gB2 plus MF59 | Phase II | - | 1997 | |||
| Simplirix/Herpevac | Glaxo-SmithKline (GSK) | gD2 and AS04 (dMPL) | Stopped after Phase III trial | Phase III | NCT00057330 | 2009 | |
| GEN-003 | Genocea | gD2 and Matrix M2 | Stopped after Phase II trial | Phase I/II | NCT01667341 | 2014 | |
| Phase II | NCT02114060 | 2016 | |||||
| NCT02300142 | 2016 | ||||||
| NCT02515175 | 2017 | ||||||
| NCT03146403 | 2018 | ||||||
| HerpV (previous AG-707) | Agenus | Peptide vaccine + QS-21 Stimulon | Stopped after Phase II trial | Phase I | NCT00231049 | 2006 | |
| Phase II | NCT01687595 | 2015 | |||||
| Live-attenuated vaccine | ICP10ΔPK | AuRx | ICP10ΔPK | Stopped after Phase I/IIa trial | Phase I/IIa | - | 2002 |
| HSV529 | Sanofi Pasteur | Replication defective HSV2, UL5, UL29 deletion | Phase II trial ongoing | Phase I | NCT01915212 | 2017 | |
| NCT02571166 | 2018 | ||||||
| Phase I/II | NCT04222985 | 2023 | |||||
| DNA vaccine | pPJV7630 | Powder-Med | Ubiquitinated and unmodified gD2 | Stopped after Phase I trial | Phase I | NCT00274300 | 2005 |
| - | Phase I | NCT00310271 | 2006 | ||||
| VCL-HB01 | Vical | gD2 +/− UL46 and Vaxfectin DNA vaccine | Stopped after Phase II trial | Phase I/II | NCT02030301 | 2016 | |
| - | Phase II | NCT02837575 | 2018 | ||||
| COR-1 | Anteris (previous ADMEDUS) | gD2 codon optimized DNA vaccine | Stopped after Phase I/IIa trial | Phase I/IIa | - | 2017 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, H.C.; Lee, H.K. Vaccines against Genital Herpes: Where Are We? Vaccines 2020, 8, 420. https://doi.org/10.3390/vaccines8030420
Kim HC, Lee HK. Vaccines against Genital Herpes: Where Are We? Vaccines. 2020; 8(3):420. https://doi.org/10.3390/vaccines8030420
Chicago/Turabian StyleKim, Hyeon Cheol, and Heung Kyu Lee. 2020. "Vaccines against Genital Herpes: Where Are We?" Vaccines 8, no. 3: 420. https://doi.org/10.3390/vaccines8030420
APA StyleKim, H. C., & Lee, H. K. (2020). Vaccines against Genital Herpes: Where Are We? Vaccines, 8(3), 420. https://doi.org/10.3390/vaccines8030420