Proteomic as an Exploratory Approach to Develop Vaccines Against Tick-Borne Diseases Using Lyme Borreliosis as a Test Case

 ,
,
Abstract
:1. Introduction
2. Material and Methods
2.1. Mouse Infection and Bacterial Strains
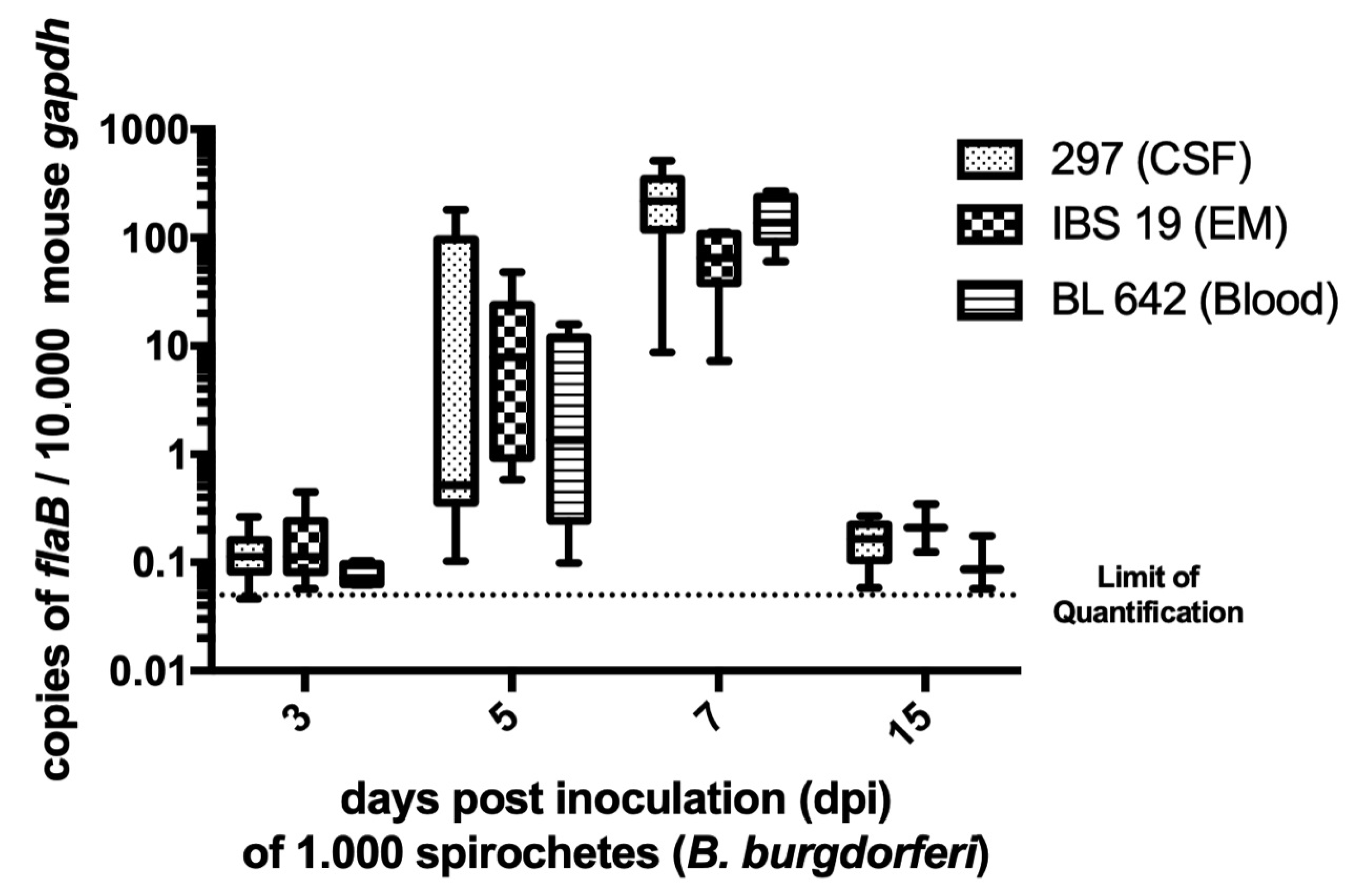
2.2. Quantification of B. burgdorferi DNA by PCR in Mouse Skin
2.3. Proteomics
2.4. In Silico Evaluation of Protein Polymorphism Among Borrelia burgdorferi Coding DNA Sequences
2.5. Ethics
2.6. Statistical Analyses
3. Results
3.1. Efficacy of Infection and Kinetics of Bacterial Multiplication in the Mouse Skin for the Three B. burgdorferi ss Strains
3.2. Detection of Borrelia Proteins on Day 7 by Proteomics
3.3. Peptidic Polymorphism of the Selected Coding DNA Sequences Among B. burgdorferi ss Genomes
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Steere, A.; Strle, F.; Wormser, G.; Hu, L.; Branda, J.; Hovius, J.; Li, X.; Mead, P. Lyme borreliosis. Nat. Rev. Dis. Prim. 2016, 2, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Gomes-Solecki, M.; Arnaboldi, P.M.; Backenson, P.B.; Benach, J.L.; Cooper, C.L.; Dattwyler, R.J.; Diuk-Wasser, M.; Fikrig, E.; Hovius, J.W.; Laegreid, W.; et al. Protective immunity and new vaccines for lyme disease. Clin. Infect. Dis. 2019. [Google Scholar] [CrossRef] [PubMed]
- Ohnishi, J.; Piesman, J.; de Silva, A. Antigenic and genetic heterogeneity of Borrelia burgdorferi populations transmitted by ticks. Proc. Nati. Acad. Sci. USA 2001, 98, 670–675. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tilly, K.; Bestor, A.; Jewett, M.W.; Rosa, P. Rapid clearance of Lyme disease spirochetes lacking OspC from skin. Infect. Immun. 2007, 75, 1517–1519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Earnhart, C.; Marconi, R. An octavalent lyme disease vaccine induces antibodies that recognize all incorporated OspC type-specific sequences. Hum. Vaccin. 2007, 3, 281–289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pal, U.; Li, X.; Wang, T.; Montgomery, R.; Ramamoorthi, N.; Desilva, A.; Bao, F.; Yang, X.; Pypaert, M.; Pradhan, D.; et al. TROSPA, an Ixodes scapularis receptor for Borrelia burgdorferi. Cell 2004, 119, 457–468. [Google Scholar] [CrossRef] [Green Version]
- Schaible, U.; Wallich, R.; Kramer, M.; Gern, L.; Anderson, J.F.; Museteanu, C.; Simon, M. Immune sera to individual Borrelia burgdorferi isolates or recombinant OspA thereof protect SCID mice against infection with homologous strains but only partially or not at all against those of different OspA/OspB genotype. Vaccine 1993, 11, 1049–1054. [Google Scholar] [CrossRef]
- Abbott, A. Lyme disease: Uphill struggle. Nature 2006, 439, 524–525. [Google Scholar] [CrossRef]
- Wressnigg, N.; Barrett, P.; Pöllabauer, E.; O’Rourke, M.; Portsmouth, D.; Schwendinger, M.; Crowe, B.; Livey, I.; Dvorak, T.; Schmitt, B.; et al. A novel multivalent OspA vaccine against Lyme borreliosis is safe and immunogenic in an adult population previously infected with Borrelia burgdorferi sensu lato. Clin. Vaccine Immunol. 2014, 21, 1490–1499. [Google Scholar] [CrossRef]
- Hagman, K.; Yang, X.; Wikel, S.; Schoeler, G.; Caimano, M.; Radolf, J.; Norgard, M. Decorin-binding protein A (DbpA) of Borrelia burgdorferi is not protective when immunized mice are challenged via tick infestation and correlates with the lack of DbpA expression by B. burgdorferi in ticks. Infect. Immun. 2000, 68, 4759–4764. [Google Scholar] [CrossRef] [Green Version]
- Brown, E.L.; Kim, J.H.; Reisenbichler, E.S.; Höök, M. Multicomponent Lyme vaccine: Three is not a crowd. Vaccine 2005, 23, 3687–3696. [Google Scholar] [CrossRef] [PubMed]
- Tsao, J.I.; Wootton, J.T.; Bunikis, J.; Luna, M.G.; Fish, D.; Barbour, A.G. Elimination of Borrelia burgdorferi from vector ticks feeding on OspA-immunized mice. Proc. Nati. Acad. Sci. USA 2004, 89, 5418–5421. [Google Scholar] [CrossRef] [Green Version]
- Gomes-Solecki, M.J.C.; Brisson, D.R.; Dattwyler, R.J. Oral vaccine that breaks the transmission cycle of the Lyme disease spirochete can be delivered via bait. Vaccine 2006, 24, 4440–4449. [Google Scholar] [CrossRef] [PubMed]
- Merino, O.; Almazan, C.; Canales, M.; Villar, M.; Moreno-Cid, J.A.; Galindo, R.C.; de la Fuente, J. Targeting the tick protective antigen subolesin reduces vector infestations and pathogen infection by Anaplasma marginale and Babesia bigemina. Vaccine 2011, 29, 8575–8579. [Google Scholar] [CrossRef] [PubMed]
- Schuijt, T.J.; Hovius, J.W.R.; van Burgel, N.D.; Ramamoorthi, N.; Fikrig, E.; van Dam, A.P. The tick salivary protein Salp15 inhibits the killing of serum-sensitive Borrelia burgdorferi sensu lato isolates. Infect. Immun. 2008, 76, 2888–2894. [Google Scholar] [CrossRef] [Green Version]
- Valenzuela, J.G.; Charlab, R.; Mather, T.N.; Ribeiro, J.M. Purification, cloning, and expression of a novel salivary anticomplement protein from the tick, Ixodes scapularis. JBC 2000, 275, 18717–18723. [Google Scholar] [CrossRef] [Green Version]
- Francischetti, I.M.B.; Valenzuela, J.G.; Andersen, J.F.; Mather, T.N.; Ribeiro, J.M.C. Ixolaris, a novel recombinant tissue factor pathway inhibitor (TFPI) from the salivary gland of the tick, Ixodes scapularis: Identification of factor X and factor Xa as scaffolds for the inhibition of factor VIIa/tissue factor complex. Blood 2002, 99, 3602–3612. [Google Scholar] [CrossRef] [Green Version]
- Kotsyfakis, M.; Sá-Nunes, A.; Francischetti, I.M.B.; Mather, T.N.; Andersen, J.F.; Ribeiro, J.M.C. Antiinflammatory and immunosuppressive activity of sialostatin L, a salivary cystatin from the tick ixodes scapularis. J. Biol. Chem. 2006, 281, 26298–26307. [Google Scholar] [CrossRef] [Green Version]
- Labuda, M.; Trimnell, A.R.; Licková, M.; Kazimírová, M.; Davies, G.M.; Lissina, O.; Hails, R.S.; Nuttall, P. An antivector vaccine protects against a lethal vector-borne pathogen. PLoS Pathog. 2006, 2, e27. [Google Scholar] [CrossRef] [Green Version]
- Kunz, C. TBE vaccination and the Austrian experience. Vaccine 2003, 21, S50–S55. [Google Scholar] [CrossRef]
- Bernard, Q.; Jaulhac, B.; Boulanger, N. Smuggling across the border: How arthropod-borne pathogens evade and exploit the host defense system of the skin. J. Investif. Dermatol. 2014, 134, 1211–1219. [Google Scholar] [CrossRef] [Green Version]
- Schnell, G.; Boeuf, A.; Westermann, B.; Jaulhac, B.; Carapito, C.; Boulanger, N.; Ehret-Sabatier, L. Discovery and targeted proteomics on cutaneous biopsies: A promising work toward an early diagnosis of Lyme disease. Mol. Cell. Proteom. 2015, 14, 1254–1264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kern, A.; Schnell, G.; Bernard, Q.; Bœuf, A.; Jaulhac, B.; Collin, E.; Barthel, C.; Ehret-Sabatier, L.; Boulanger, N. Heterogeneity of borrelia burgdorferi sensu stricto population and its involvement in borrelia pathogenicity: Study on murine model with specific emphasis on the skin interface. PLoS ONE 2015, 10, e0133195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nebbak, A.; Dahmana, H.; Almeras, L.; Raoult, D.; Boulanger, N.; Jaulhac, B.; Mediannikov, O.; Parola, P. Co-infection of bacteria and protozoan parasites in Ixodes ricinus nymphs collected in the Alsace region, France. Ticks Tick. Borne. Dis. 2019, 10, 101241. [Google Scholar] [CrossRef] [PubMed]
- Gissot, M.; Hovasse, A.; Chaloin, L.; Schaeffer-Reiss, C.; Van Dorsselaer, A.; Tomavo, S. An evolutionary conserved zinc finger protein is involved in Toxoplasma gondii mRNA nuclear export. Cell. Microbiol. 2017, 19, e12644. [Google Scholar] [CrossRef] [PubMed]
- Grillon, A.; Westermann, B.; Cantero, P.; Jaulhac, B.; Voordouw, M.; Kapps, D.; Collin, E.; Barthel, C.; Ehret-Sabatier, L.; Boulanger, N. Identification of Borrelia protein candidates in mouse skin for potential diagnosis of disseminated Lyme borreliosis. Sci. Rep. 2017, 7, 16719. [Google Scholar] [CrossRef] [Green Version]
- Di, L.; Pagan, P.E.; Packer, D.; Martin, C.L.; Akther, S.; Ramrattan, G.; Mongodin, E.F.; Fraser, C.M.; Schutzer, S.E.; Luft, B.J.; et al. BorreliaBase: A phylogeny-centered browser of Borrelia genomes. BMC Bioinform. 2014, 15, 233. [Google Scholar] [CrossRef] [Green Version]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Embers, M.E.; Narasimhan, S. Vaccination against Lyme disease: Past, present, and future. Front. Cell. Infect. Microbiol. 2013, 3. [Google Scholar] [CrossRef] [Green Version]
- Rogovskyy, A.S.; Bankhead, T. Variable VlsE is critical for host reinfection by the lyme disease spirochete. PLoS ONE 2013, 8, e61226. [Google Scholar] [CrossRef] [Green Version]
- Kraiczy, P.; Stevenson, B. Complement regulator-acquiring surface proteins of Borrelia burgdorferi: Structure, function and regulation of gene expression. Ticks Tick. Borne. Dis. 2013, 4, 26–34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elsner, R.A.; Hastey, C.J.; Olsen, K.J.; Baumgarth, N. Suppression of long-lived humoral immunity following borrelia burgdorferi infection. PLoS Pathog. 2015, 11, e1004976. [Google Scholar] [CrossRef] [PubMed]
- Shaffer, L. Inner workings: Lyme disease vaccines face familiar challenges, both societal and scientific. Proc. Natl. Acad. Sci. USA 2019, 116, 19214–19217. [Google Scholar] [CrossRef] [Green Version]
- Comstedt, P.; Hanner, M.; Schüler, W.; Meinke, A.; Lundberg, U. Design and development of a novel vaccine for protection against Lyme borreliosis. PLoS ONE 2014, 9, e113294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Comstedt, P.; Schüler, W.; Meinke, A.; Lundberg, U. The novel Lyme borreliosis vaccine VLA15 shows broad protection against Borrelia species expressing six different OspA serotypes. PLoS ONE 2017, 12, e0184357. [Google Scholar] [CrossRef] [PubMed]
- Barthold, S.; de Souza, M.; Janotka, J.; Smith, A.; Persing, D. Chronic Lyme borreliosis in the laboratory mouse. Am. J. Pathol. 1993, 143, 959–971. [Google Scholar]
- Margos, G.; Vollmer, S.A.; Ogden, N.H.; Fish, D. Population genetics, taxonomy, phylogeny and evolution of Borrelia burgdorferi sensu lato. Infect. Genet. Evol. 2011, 11, 1545–1563. [Google Scholar] [CrossRef] [Green Version]
- Aguero-Rosenfeld, M.E.; Nowakowski, J.; McKenna, D.F.; Carbonaro, C.A.; Wormser, G.P. Serodiagnosis in early Lyme disease. J. Clin. Microbiol. 1993, 31, 3090–3095. [Google Scholar] [CrossRef] [Green Version]
- Sultan, S.Z.; Manne, A.; Stewart, P.E.; Bestor, A.; Rosa, P.A.; Charon, N.W.; Motaleb, M.A. Motility is crucial for the infectious life cycle of borrelia burgdorferi. Infect. Immun. 2013, 81, 2012–2021. [Google Scholar] [CrossRef] [Green Version]
- Fikrig, E.; Barthold, S.W.; Marcantonio, N.; Deponte, K.; Kantor, F.S.; Flavell, R.A. Roles of OspA, OspB, and flagellin in protective immunity to Lyme borreliosis in laboratory mice. Infect. Immun. 1992, 60, 657–661. [Google Scholar] [CrossRef] [Green Version]
- Cui, B.; Liu, X.; Fang, Y.; Zhou, P.; Zhang, Y.; Wang, Y. Flagellin as a vaccine adjuvant. Expert Rev. Vaccines 2018, 17, 335–349. [Google Scholar] [CrossRef] [PubMed]
- Meunier, M.; Guyard-Nicodème, M.; Vigouroux, E.; Poezevara, T.; Béven, V.; Quesne, S.; Amelot, M.; Parra, A.; Chemaly, M.; Dory, D. A DNA prime/protein boost vaccine protocol developed against Campylobacter jejuni for poultry. Vaccine 2018, 36, 2119–2125. [Google Scholar] [CrossRef] [PubMed]
- Bruxelle, J.-F.; Mizrahi, A.; Hoÿs, S.; Collignon, A.; Janoir, C.; Péchiné, S. Clostridium difficile flagellin FliC: Evaluation as adjuvant and use in a mucosal vaccine against Clostridium difficile. PLoS ONE 2017, 12, e0187212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tilly, K.; Krum, J.G.; Bestor, A.; Jewett, M.W.; Grimm, D.; Bueschel, D.; Byram, R.; Dorward, D.; Vanraden, M.J.; Stewart, P.; et al. Borrelia burgdorferi OspC protein required exclusively in a crucial early stage of mammalian infection. Infect. Immun. 2006, 74, 3554–3564. [Google Scholar] [CrossRef] [Green Version]
- Brisson, D.; Baxamusa, N.; Schwartz, I.; Wormser, G.P. Biodiversity of Borrelia burgdorferi strains in tissues of Lyme disease patients. PLoS ONE 2011, 6, e22926. [Google Scholar] [CrossRef] [Green Version]
- Toledo, A.; Coleman, J.; Kuhlow, C.; Crowley, J.; Benach, J. The enolase of Borrelia burgdorferi is a plasminogen receptor released in outer membrane vesicles. Infect Immun. 2012, 80, 359–368. [Google Scholar] [CrossRef] [Green Version]
- Cappello, P.; Principe, M.; Bulfamante, S.; Novelli, F. Alpha-Enolase (ENO1), a potential target in novel immunotherapies. Front. Biosci. 2017, 22, 944–959. [Google Scholar] [CrossRef]
- Nogueira, S.V.; Smith, A.A.; Qin, J.-H.; Pal, U. A surface enolase participates in Borrelia burgdorferi-plasminogen interaction and contributes to pathogen survival within feeding ticks. Infect. Immun. 2012, 80, 82–90. [Google Scholar] [CrossRef] [Green Version]
- Nowalk, A.J.; Nolder, C.; Clifton, D.R.; Carroll, J.A. Comparative proteome analysis of subcellular fractions from Borrelia burgdorferi by NEPHGE and IPG. Proteomics 2006, 6, 2121–2134. [Google Scholar] [CrossRef]
- Lee, S.-H.; Lee, J.-H.; Park, H.-S.; Jang, W.-J.; Koh, S.-E.; Yang, Y.-M.; Kim, B.-J.; Kook, Y.-H.; Park, K.-H. Differentiation of Borrelia burgdorferi sensu lato through groEL gene analysis. FEMS Microbiol. Lett. 2003, 222, 51–57. [Google Scholar] [CrossRef]
- Jungblut, P.; Grabher, G.; Stöffler, G. Comprehensive detection of immunorelevant Borrelia garinii antigens by two-dimensional electrophoresis. Electrophoresis 1999, 20, 3611–3622. [Google Scholar] [CrossRef]
- Carrasco, S.; Yang, Y.; Troxell, B.; Yang, X.; Pal, U.; Yang, X. Borrelia burgdorferi elongation factor EF-Tu is an immunogenic protein during Lyme borreliosis. Emerg. Microbes Infect. 2015, 4, e54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de la Fuente, J.; Kocan, K.; Almazan, C.; Blouin, E. Targeting the tick-pathogen interface for novel control strategies. Front Biosci. 2008, 13, 6947–6956. [Google Scholar] [CrossRef] [PubMed]
- Caljon, G.; Van Reet, N.; De Trez, C.; Vermeersch, M.; Pérez-Morga, D.; Van Den Abbeele, J. The Dermis as a delivery site of trypanosoma brucei for tsetse flies. PLoS Pathog. 2016, 12, e1005. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ménard, R.; Tavares, J.; Cockburn, I.; Markus, M.; Zavala, F.; Amino, R. Looking under the skin: The first steps in malarial infection and immunity. Nat. Rev. Microbiol. 2013, 11, 701–712. [Google Scholar] [CrossRef] [PubMed]
- Kuleš, J.; Horvatić, A.; Guillemin, N.; Galan, A.; Mrljak, V.; Bhide, M. New approaches and omics tools for mining of vaccine candidates against vector-borne diseases. Mol. Biosyst. 2016, 12, 2680–2694. [Google Scholar]
- Moxon, R.; Reche, P.A.; Rappuoli, R. Editorial: Reverse vaccinology. Front. Immunol. 2019, 10, 2776. [Google Scholar] [CrossRef] [Green Version]
- Gesslbauer, B.; Poljak, A.; Handwerker, C.; Schüler, W.; Schwendenwein, D.; Weber, C.; Lundberg, U.; Meinke, A.; Kungl, A.J. Comparative membrane proteome analysis of three Borrelia species. Proteomics 2012, 12, 845–858. [Google Scholar] [CrossRef]
- Bencurova, E.; Gupta, S.K.; Oskoueian, E.; Bhide, M.; Dandekar, T. Omics and bioinformatics applied to vaccine development against Borrelia. Mol. Omi. 2018, 14, 330–340. [Google Scholar] [CrossRef]
- Iyer, R.; Caimano, M.J.; Luthra, A.; Axline, D.; Corona, A.; Iacobas, D.A.; Radolf, J.D.; Schwartz, I. Stage-specific global alterations in the transcriptomes of Lyme disease spirochetes during tick feeding and following mammalian host adaptation. Mol. Microbiol. 2015, 95. [Google Scholar] [CrossRef] [Green Version]
- Caimano, M.J.; Groshong, A.M.; Belperron, A.; Mao, J.; Hawley, K.L.; Luthra, A.; Graham, D.E.; Earnhart, C.G.; Marconi, R.T.; Bockenstedt, L.K.; et al. The RpoS gatekeeper in Borrelia burgdorferi: An invariant regulatory scheme that promotes spirochete persistence in reservoir hosts and niche diversity. Front. Microbiol. 2019, 10, 1923. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.E.; Fischbach, M.A.; Belkaid, Y. Skin microbiota-host interactions. Nature 2018, 553, 427–436. [Google Scholar] [CrossRef] [PubMed]
- Buckley, P.R.; Alden, K.; Coccia, M.; Chalon, A.; Collignon, C.; Temmerman, S.T.; Didierlaurent, A.M.; van der Most, R.; Timmis, J.; Andersen, C.A.; et al. Application of Modeling Approaches to Explore Vaccine Adjuvant Mode-of-Action. Front. Immunol. 2019, 10, 2150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Donadei, A.; Kraan, H.; Ophorst, O.; Flynn, O.; O’Mahony, C.; Soema, P.C.; Moore, A.C. Skin delivery of trivalent Sabin inactivated poliovirus vaccine using dissolvable microneedle patches induces neutralizing antibodies. J. Control. Release 2019, 311–312, 96–103. [Google Scholar] [CrossRef] [PubMed]
- Rouphael, N.G.; Paine, M.; Mosley, R.; Henry, S.; McAllister, D.V.; Kalluri, H.; Pewin, W.; Frew, P.M.; Yu, T.; Thornburg, N.J.; et al. The safety, immunogenicity, and acceptability of inactivated influenza vaccine delivered by microneedle patch (TIV-MNP 2015): A randomised, partly blinded, placebo-controlled, phase 1 trial. Lancet 2017, 390, 649–658. [Google Scholar] [CrossRef]


{kind=link}
| Days Post-Inoculation (dpi) | Strains | Positive PCR Amplification | |
|---|---|---|---|
| Positive Reactions/No. of Tested Animals | Rate of Positivity (CI 95%) | ||
| 3 dpi | 297 | 14/19 | 74% (49–91) |
| IBS 19 | 6/11 | 55% (23–83) | |
| BL 642 | 4/11 | 36% (11–69) | |
| 5 dpi | 297 | 16/17 | 94% (71–100) |
| IBS 19 | 8/8 | 100% (63–100) | |
| BL 642 | 6/7 | 86% (42–100) | |
| 7 dpi | 297 | 15/15 | 100% (78–100) |
| IBS 19 | 10/10 | 100% (69–100) | |
| BL 642 | 6/6 | 100% (54–100) | |
| 15 dpi | 297 | 6/13 | 46% (19–75) |
| IBS 19 | 3/7 | 43% (10–82) | |
| BL 642 | 3/7 | 43% (10–82) | |
| Protein Name | Accession Number (Borrelia burgdorferi B31) | Accession Number (Borrelia burgdorferi Pko) | Infection via Syringe Inoculation Borrelia burgdorferi Strain (Spirochete Density Flagellin/104gapdh) | Infection via Tick Bite (Spirochete Density Flagellin/104gapdh) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 297c4 (192) | 297c4 (106) | 297n (346) | IBS19 (85) | IBS19 (136) | IBS19 (166) | BL642 (157) | BL642 (227) | BL642 (268) | B. Afzelii NE4049 (66) | Field-Collected Ticks (534) | |||
| Flagellin | gi|120230 | gi|111114970 | 7 | 8 | 7 | 2 | 2 | 3 | 3 | 2 | 4 | 8 | |
| Osp C | gi|3914248 | gi|111074137 | 3 | 2 | 3 | 1 | 2 | 2 | 2 | 1 | 1 | 4 | 1 |
| GAPDH | gi|238828321 gi|3915702 | 1 | 1 | 1 | 1 | 1 | |||||||
| Glycosaminoglycan binding Protein (Bgp) | gi|15594933 | 1 | 1 | 1 | |||||||||
| GroEL | gi|229553917 | gi|123145654 | 1 | 1 | 2 | ||||||||
| p66 | gi|15594948 | 1 | |||||||||||
| Fructose-bisphosphate aldolase | gi|6686370 | gi|111115272 | 1 | ||||||||||
| DbpA | gi|17373807 | 1 | |||||||||||
| DbpB | gi|327507700 | 2 | |||||||||||
| Elongation factor 4 | gi|15594434 gi|6016495 | 1 | |||||||||||
| Elongation factor Tu | gi|1706598 | gi|123341337 | 1 | 1 | 1 | 2 | |||||||
| Enolase | gi|3913583 | 1 | |||||||||||
| Lipoprotein BbA36 | gi|365823350 | 2 | |||||||||||
| L-lactate dehydrogenase | gi|15594433 gi|17367476 | 1 | |||||||||||
| hypothetical protein BB_0363 | gi|15594708 | 1 | |||||||||||
| hypothetical protein BB_0563 | gi|365992369 | 1 | 1 | ||||||||||
| hypothetical protein BB_F14 | gi|365823340 | 1 | 1 | ||||||||||
| hypothetical protein BB_J48 | gi|364556751 | 1 | |||||||||||
| hypothetical protein BAPKO_4515 | gi|117621815 | 1 | |||||||||||
| hypothetical protein BAPKO_0028 | gi|111114851 | 1 | |||||||||||
| hypothetical protein BAPKO_0593 | gi|111115391 | 1 | |||||||||||
| hypothetical protein BAPKO_2500 | gi|117621647 | 1 | |||||||||||
| 50S ribosomal protein L7/L12 | gi|123046997 | 1 | |||||||||||
| 30S ribosomal protein S16 | gi|123145645 | 1 | 1 | ||||||||||
| Neutrophil activating protein | gi|111115523 | 2 | |||||||||||
| Phosphoglyceromutase | gi|123145651 | 1 | |||||||||||
| Flagellar filament outer layer protein | gi|111115501 | 1 | |||||||||||
| ATP-dependent protease | gi|111115078 | 1 | 1 | ||||||||||
| Periplasmic oligopeptide-binding protein | gi|111115153 | 1 | |||||||||||
| Chemotaxis protein methyltransferase | gi|111114863 | 1 | |||||||||||
| Number of Borrelia proteins | 9 | 6 | 7 | 2 | 2 | 2 | 3 | 6 | 5 | 7 | 15 | ||
| Number of mouse proteins | 1831 | 1791 | 1824 | 1486 | 1617 | 1550 | 1702 | 1351 | 1426 | 5916 | 5926 | ||
| Protein (Length) | Protein Accession Number (B31) | Gene Accession Number (B31) | CDS Location | Peptidic Variability Among the CDS of 15 B. burgdorferi Sensu Stricto Strains |
|---|---|---|---|---|
| FlaB (336 aa) | SwissProt P11089 | Genbank BB_0147 | chromosome | 100% S./100% Id.: 15/15 strains |
| GAPDH (335 aa) | SwissProt P46795 | Genbank BB_0057 | chromosome | 100% S./100% Id.: 14/15 strains 100% S./99% Id.: 1/15 strain |
| GroEL (545 aa) | SwissProt P0C923 | Genbank BB_0649 | chromosome | 100% S./100% Id.: 15/15 strains 1 supplementary aa in 1 strain |
| Enolase (433 aa) | SwissProt O51312 | Genbank BB_0337 | chromosome | 100% S./100% Id.: 12/15 strains 100% S./99% Id.: 2/15 strains Truncated peptidic CDS in the 29805 strain (from 219th to 273th aa) |
| Lipoprotein BbA36 (212 aa) | TrEmbl O50929 | Genbank BB_A36 | lp54 plasmid(13/15 strains) | 100% S./100% Id.: 2/13 strains 100% S./99% Id.: 2/13 strains 97–99% S./97–99% Id.: 9/13 strains Not found in 2 WGS without sequenced lp54 plasmid 8 first aa not annotated in the 12 strains other than B31 1 supplementary Asparagin between the 80th and 81th residue of the reference sequence in 6/13 strains |
| Outer surface protein C (210 aa) | SwissProt Q07337 | Genbank BB_B19 | cp26 plasmid(13/15 strains) | 100% S./100% Id.: 1/13 strains 82–88% S./74–81% Id.: 12/13 strains Not found in 2 WGS without sequenced cp26 plasmid |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Talagrand-Reboul, E.; Westermann, B.; Raess, M.A.; Schnell, G.; Cantero, P.; Barthel, C.; Ehret-Sabatier, L.; Jaulhac, B.; Boulanger, N. Proteomic as an Exploratory Approach to Develop Vaccines Against Tick-Borne Diseases Using Lyme Borreliosis as a Test Case. Vaccines 2020, 8, 463. https://doi.org/10.3390/vaccines8030463
Talagrand-Reboul E, Westermann B, Raess MA, Schnell G, Cantero P, Barthel C, Ehret-Sabatier L, Jaulhac B, Boulanger N. Proteomic as an Exploratory Approach to Develop Vaccines Against Tick-Borne Diseases Using Lyme Borreliosis as a Test Case. Vaccines. 2020; 8(3):463. https://doi.org/10.3390/vaccines8030463
Chicago/Turabian StyleTalagrand-Reboul, Emilie, Benoit Westermann, Matthieu A. Raess, Gilles Schnell, Paola Cantero, Cathy Barthel, Laurence Ehret-Sabatier, Benoit Jaulhac, and Nathalie Boulanger. 2020. "Proteomic as an Exploratory Approach to Develop Vaccines Against Tick-Borne Diseases Using Lyme Borreliosis as a Test Case" Vaccines 8, no. 3: 463. https://doi.org/10.3390/vaccines8030463
APA StyleTalagrand-Reboul, E., Westermann, B., Raess, M. A., Schnell, G., Cantero, P., Barthel, C., Ehret-Sabatier, L., Jaulhac, B., & Boulanger, N. (2020). Proteomic as an Exploratory Approach to Develop Vaccines Against Tick-Borne Diseases Using Lyme Borreliosis as a Test Case. Vaccines, 8(3), 463. https://doi.org/10.3390/vaccines8030463