Immune-Enhancing Activity of Aqueous Extracts from Artemisia rupestris L. via MAPK and NF-kB Pathways of TLR4/TLR2 Downstream in Dendritic Cells

 and
and 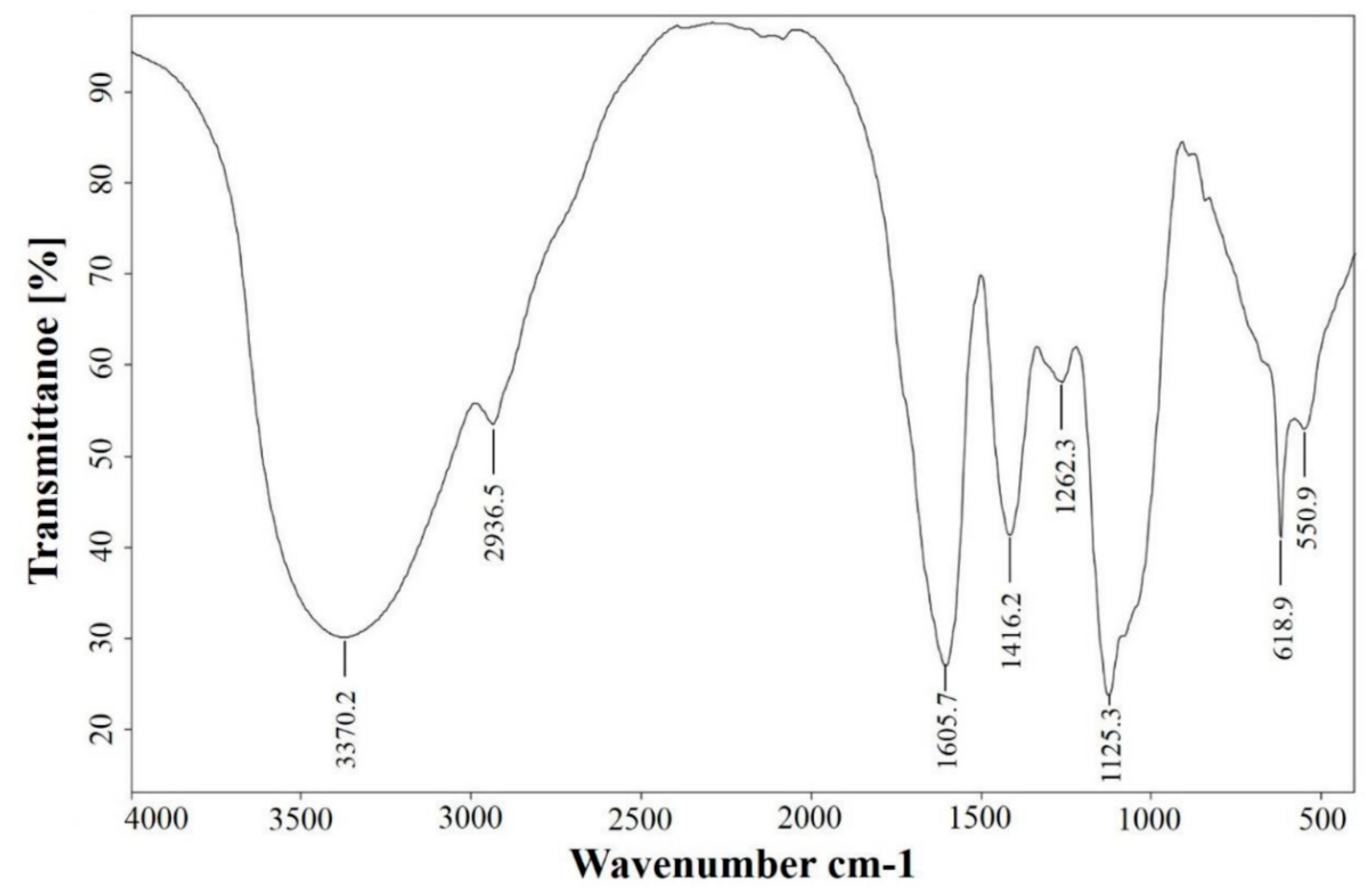{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials and Reagents
2.2. Infrared Spectroscopic Analysis
2.3. Animals and Generation of DCs
2.4. Cell Viability Assays
2.5. Morphological Observations
2.6. Flow Cytometric Detection of Apoptotic Cells
2.7. Flow Cytometric Phenotype Analysis of DCs
2.8. ELISA Analysis of Cytokines
2.9. Endocytosis Assays
2.10. Mixed Lymphocyte Reaction
2.11. Blocking of TLR Pathways with Monoclonal Antibodies
2.12. Western Blotting Analysis
2.13. Data Analysis
3. Results
3.1. FT-IR Spectral Features of AEAR
3.2. Effects of AEAR on Cytotoxicity
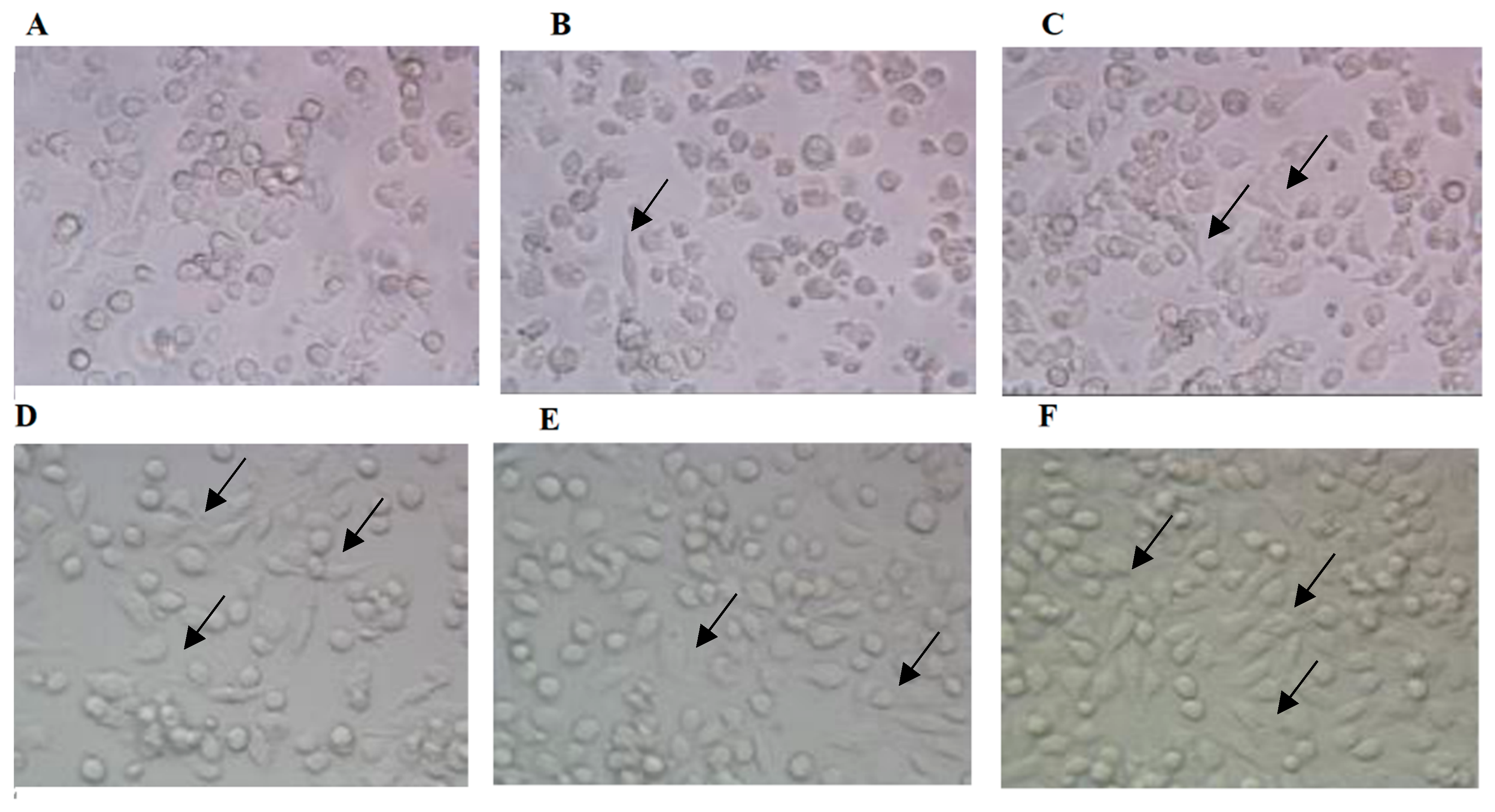
3.3. Effects of AEAR on Morphological Changes of Cells
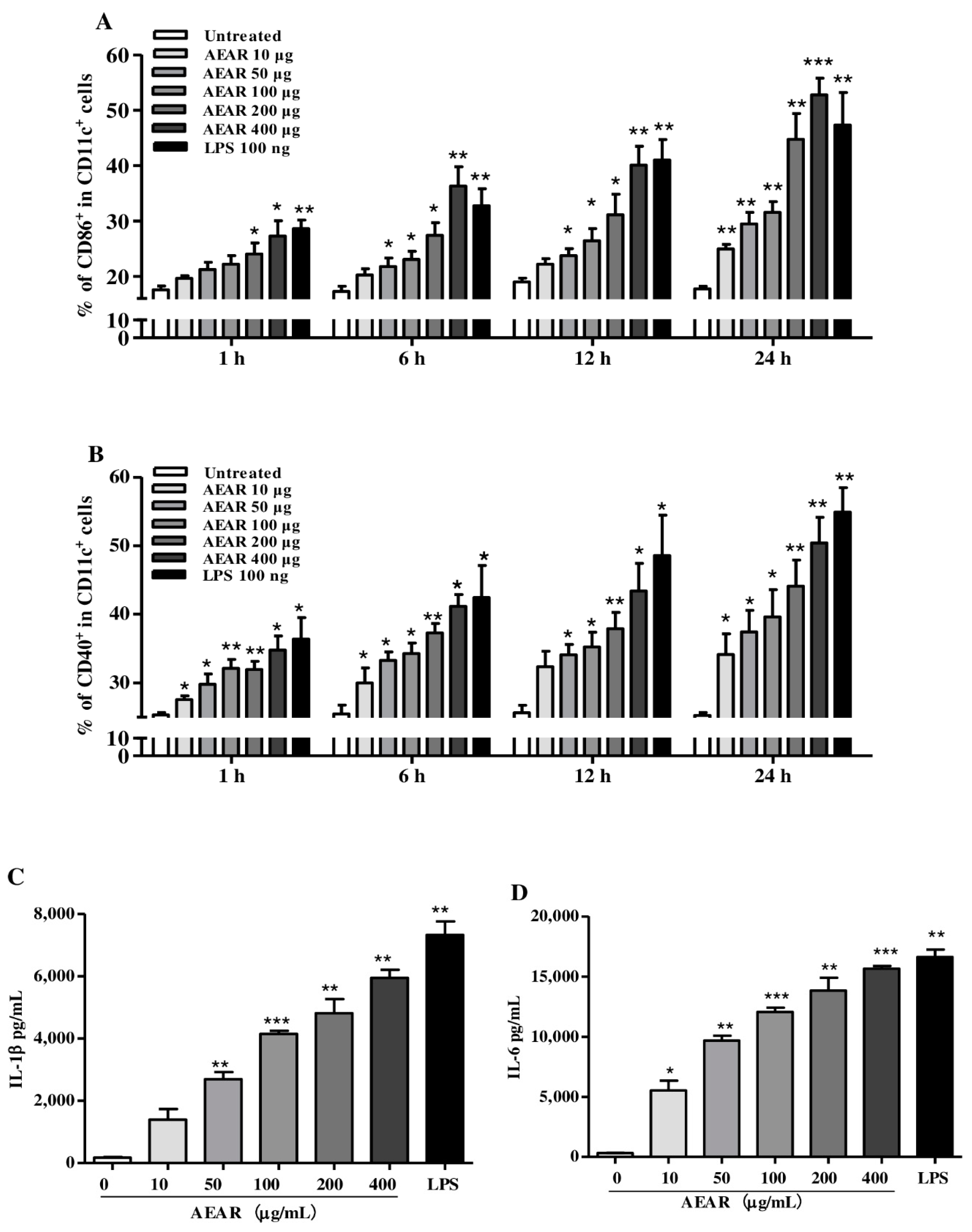
3.4. AEAR Induced DCs’ Activation
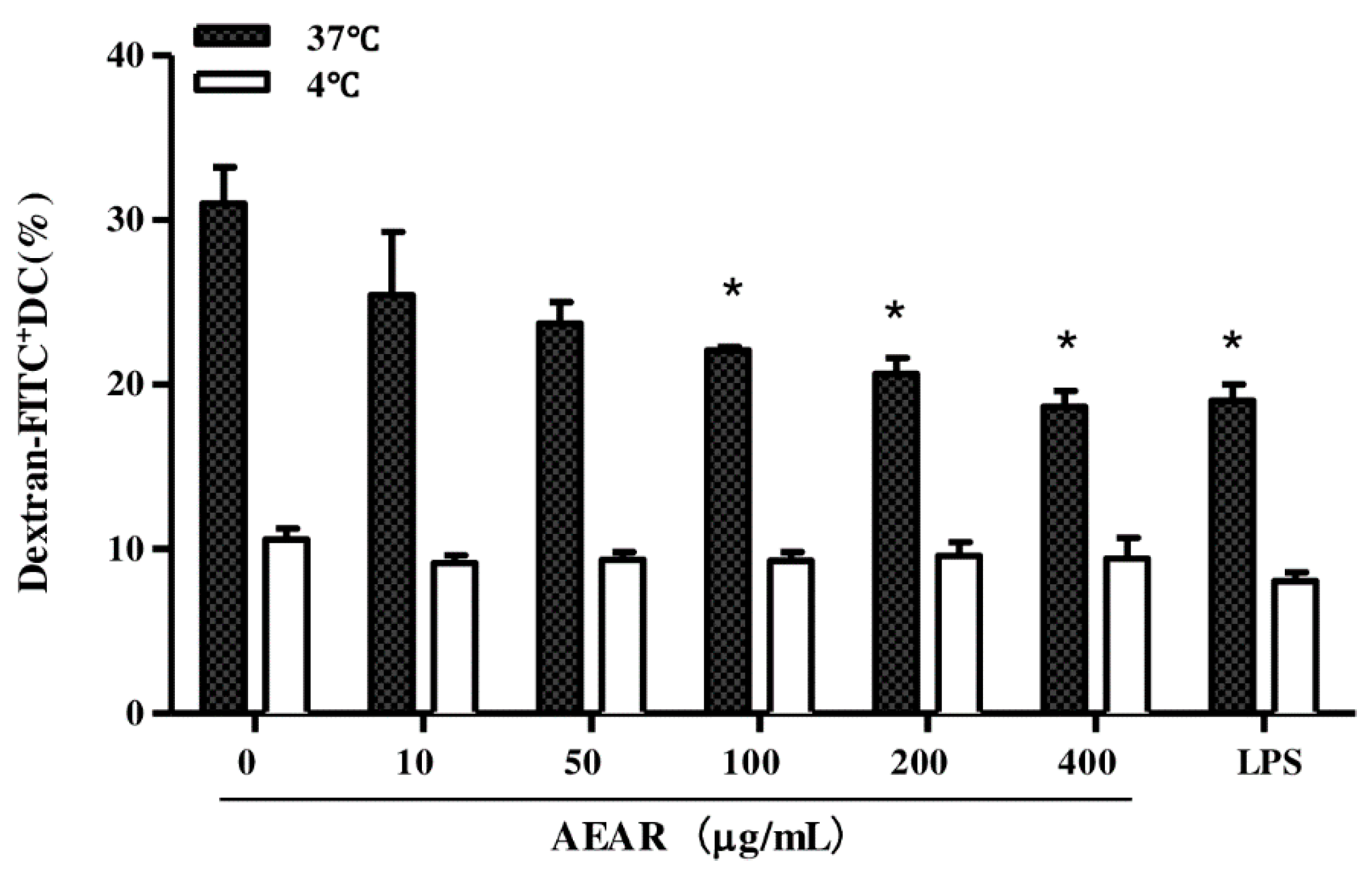
3.5. Effects of AEAR on Endocytosis Activity of DCs
3.6. AEAR-Treated DCs Enhanced Splenocyte Proliferation
3.7. TLR2 Mediated DCs’ Activation by AEAR
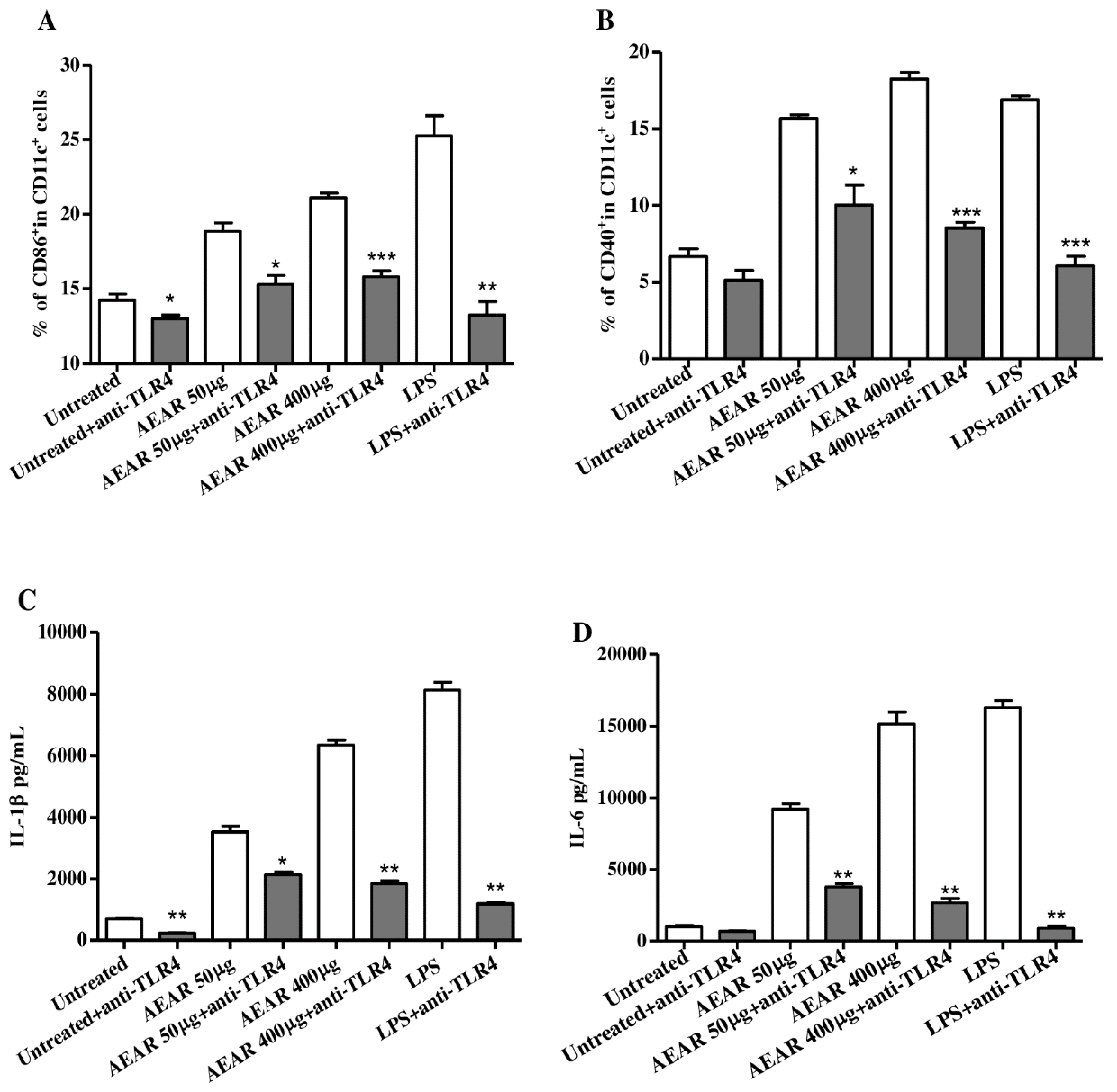
3.8. TLR4 Mediated DCs Activation by AEAR
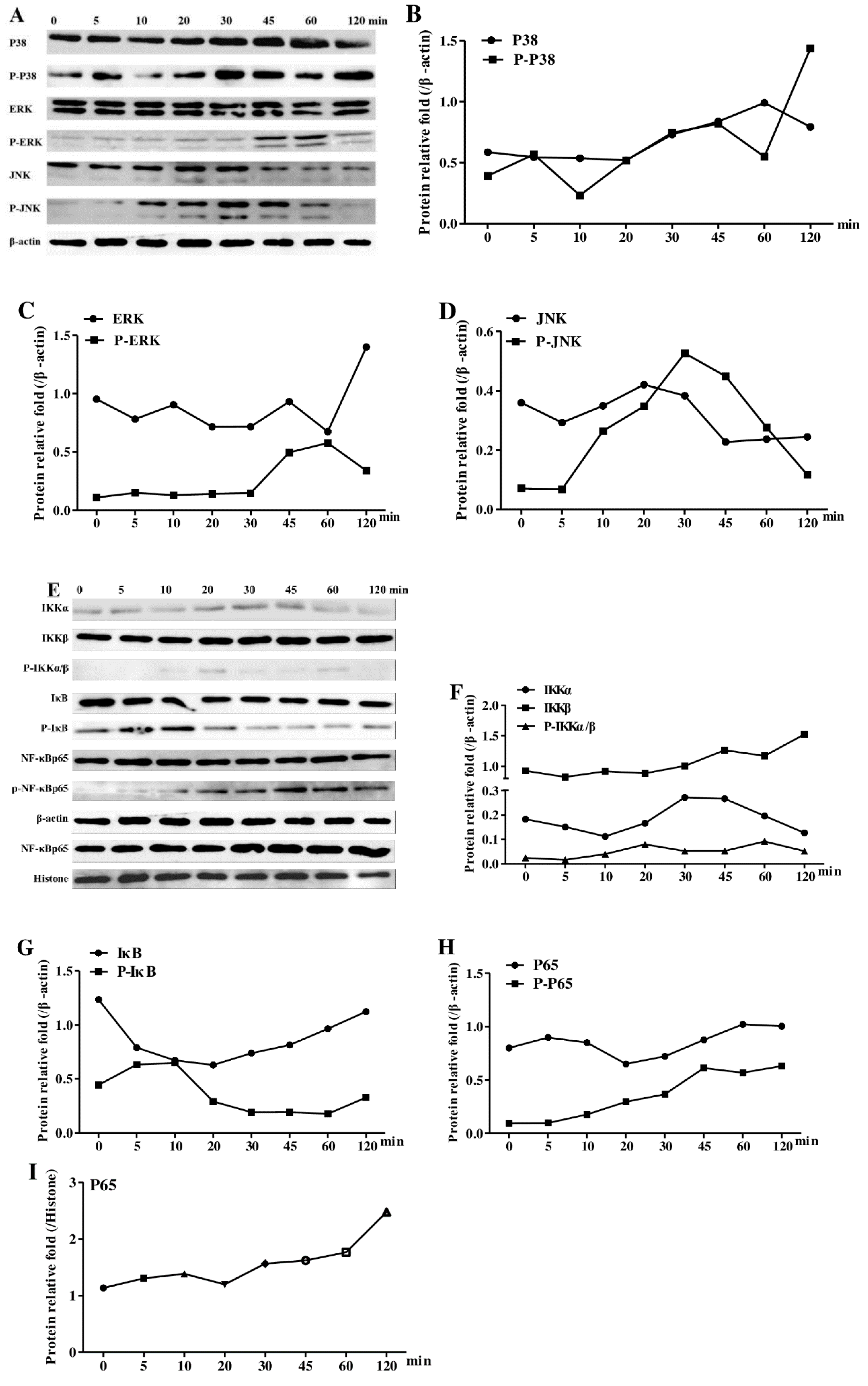
3.9. Effects of AEAR on the Activation of NF-kB and MAPKs in DCs
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
Ethics Statements
References
- Harandi, A.M. Systems analysis of human vaccine adjuvants. Semin. Immunol. 2018, 39, 30–34. [Google Scholar] [CrossRef] [PubMed]
- Tomar, J.; Patil, H.P.; Bracho, G.; Tonnis, W.F.; Frijlink, H.W.; Petrovsky, N.; Hinrichs, W.L.; Vanbever, R.; Huckriede, A.; Hinrichs, W.L.J. Advax augments B and T cell responses upon influenza vaccination via the respiratory tract and enables complete protection of mice against lethal influenza virus challenge. J. Control. Release 2018, 288, 199–211. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Zhu, X.; Ma, J.; Zhang, M.; Wu, H. Structural Elucidation of a Novel Pectin-Polysaccharide from the Petal of Saussurea laniceps and the Mechanism of its Anti-HBV Activity. Carbohydr. Polym. 2019, 223, 115077. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Xing, Q.Q.; Xu, J.Y.; Ding, D.; Zhao, X. Astragalus polysaccharide modulates ER stress response in an OVA-LPS induced murine model of severe asthma. Int. J. Biol. Macromol. 2016, 93, 995–1006. [Google Scholar] [CrossRef]
- Liu, K.; Yin, Y.; Zhang, J.; Zai, X.; Li, R.; Ma, H.; Xu, J.; Shan, J.; Chen, W. Polysaccharide PCP-I isolated from Poria cocos enhances the immunogenicity and protection of an anthrax protective antigen-based vaccine. Hum. Vaccines Immunother. 2019, 1–9. [Google Scholar] [CrossRef]
- Ullah, S.; Khalil, A.A.; Shaukat, F.; Song, Y. Sources, Extraction and Biomedical Properties of Polysaccharides. Foods 2019, 8, 304. [Google Scholar] [CrossRef] [Green Version]
- Seidi, F.; Jenjob, R.; Phakkeeree, T.; Crespy, D. Saccharides, oligosaccharides, and polysaccharides nanoparticles for biomedical applications. J. Control. Release 2018, 284, 188–212. [Google Scholar] [CrossRef]
- Ganesan, K.; Xu, B. Anti-Diabetic Effects and Mechanisms of Dietary Polysaccharides. Molecules 2019, 24, 2556. [Google Scholar] [CrossRef] [Green Version]
- Gupta, N.; Jangid, A.K.; Pooja, D.; Kulhari, H. Inulin: A novel and stretchy polysaccharide tool for biomedical and nutritional applications. Int. J. Biol. Macromol. 2019, 132, 852–863. [Google Scholar] [CrossRef]
- Jin, M.; Zhao, K.; Huang, Q.; Shang, P. Structural features and biological activities of the polysaccharides from Astragalus membranaceus. Int. J. Biol. Macromol. 2014, 64, 257–266. [Google Scholar] [CrossRef]
- Masci, A.; Carradori, S.; Casadei, M.A.; Paolicelli, P.; Petralito, S.; Ragno, R.; Cesa, S. Lycium barbarum polysaccharides: Extraction, purification, structural characterisation and evidence about hypoglycaemic and hypolipidaemic effects. A review. Food Chem. 2018, 254, 377–389. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Ji, T.; Song, W.; Wang, A.; Su, Y. Chemical constituents from herb of Artemisia rupestris. Chin. Tradit. Herb. Drugs 2008, 39, 1125. [Google Scholar]
- Zhang, A.; Wang, D.; Li, J.; Gao, F.; Fan, X. The effect of aqueous extract of Xinjiang Artemisia rupestris L. (an influenza virus vaccine adjuvant) on enhancing immune responses and reducing antigen dose required for immunity. PLoS ONE 2017, 12, e0183720. [Google Scholar] [CrossRef] [PubMed]
- Juteau, F.; Masotti, V.; Bessiere, J.M.; Dherbomez, M.; Viano, J. Antibacterial and antioxidant activities of Artemisia annua essential oil. Fitoterapia 2002, 73, 532–535. [Google Scholar] [CrossRef]
- Zhang, A.L.; Yang, Y.; Wang, Y.; Zhao, G.; Yang, X.M.; Wang, D.Y.; Wang, B. Adjuvant-active aqueous extracts from Artemisia rupestris L. improve immune responses through TLR4 signaling pathway. Vaccine 2017, 35, 1037–1045. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Cao, H.; Li, J.; Zhao, B.; Wang, Y.; Zhang, A.; Huang, J. Adjuvanticity of aqueous extracts of Artemisia rupestris L. for inactivated foot-and-mouth disease vaccine in mice. Res. Vet. Sci. 2019, 124, 191–199. [Google Scholar] [CrossRef] [PubMed]
- Banchereau, J.; Steinman, R.M. Dendritic cells and the control of immunity. Nature 1998, 392, 245–252. [Google Scholar] [CrossRef]
- Akira, S. Toll receptor families: Structure and function. Semin. Immunol. 2004, 16, 1–2. [Google Scholar] [CrossRef]
- Medzhitov, R. Recognition of microorganisms and activation of the immune response. Nature 2007, 449, 819–826. [Google Scholar] [CrossRef]
- Janeway, C.A., Jr.; Medzhitov, R. Innate immune recognition. Annu. Rev. Immunol. 2002, 20, 197–216. [Google Scholar] [CrossRef] [Green Version]
- Kawai, T.; Akira, S. Signaling to NF-κB by Toll-like receptors. Trends Mol. Med. 2007, 13, 460–469. [Google Scholar] [CrossRef] [PubMed]
- Lei, W.; Browning, J.D., Jr.; Eichen, P.A.; Lu, C.H.; Mossine, V.V.; Rottinghaus, G.E.; Folk, W.R.; Sun, G.Y.; Lubahn, D.B.; Fritsche, K.L. Immuno-stimulatory activity of a polysaccharide-enriched fraction of Sutherlandia frutescens occurs by the toll-like receptor-4 signaling pathway. J. Ethnopharmacol. 2015, 172, 247–253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peri, F.; Piazza, M. Therapeutic targeting of innate immunity with Toll-like receptor 4 (TLR4) antagonists. Biotechnol. Adv. 2012, 30, 251–260. [Google Scholar] [CrossRef] [PubMed]
- Wetzler, L.M. The role of Toll-like receptor 2 in microbial disease and immunity. Vaccine 2003, 21 (Suppl. 2), S55–S60. [Google Scholar] [CrossRef]
- Guo, X.P.; Tian, C.R.; Gao, C.Y.; Meng, Y.W. Study on removal process of proteins from crude Sphallerocarpus gracilis polysaccharides and its scavenging capability to nitrite. Sci. Technol. Food Ind. 2011, 32, 274–276. [Google Scholar] [CrossRef] [Green Version]
- Sheu, S.C.; Lyu, Y.; Lee, M.S.; Cheng, J.H. Immunomodulatory effects of polysaccharides isolated from Hericium erinaceus on dendritic cells. Process Biochem. 2013, 48, 1402–1408. [Google Scholar] [CrossRef]
- Huang, D.F.; Nie, S.P.; Jiang, L.M.; Xie, M.Y. A novel polysaccharide from the seeds of Plantago asiatica L. induces dendritic cells maturation through toll-like receptor 4. Int. Immunopharmacol. 2014, 18, 236–243. [Google Scholar] [CrossRef]
- Engering, A.J.; Cella, M.; Fluitsma, D.M.; Hoefsmit, E.C.; Lanzavecchia, A.; Pieters, J. Mannose receptor mediated antigen uptake and presentation in human dendritic cells. Adv. Exp. Med. Biol. 1997, 417, 183–187. [Google Scholar] [CrossRef]
- Bonam, S.R.; Partidos, C.D.; Halmuthur, S.K.M.; Muller, S. An Overview of Novel Adjuvants Designed for Improving Vaccine Efficacy. Trends Pharmacol. Sci. 2017, 38, 771–793. [Google Scholar] [CrossRef]
- Tom, J.K.; Albin, T.J.; Manna, S.; Moser, B.A.; Steinhardt, R.C.; Esser-Kahn, A.P. Applications of Immunomodulatory Immune Synergies to Adjuvant Discovery and Vaccine Development. Trends Biotechnol. 2019, 37, 373–388. [Google Scholar] [CrossRef]
- Tian, H.; Liu, Z.; Pu, Y.; Bao, Y. Immunomodulatory effects exerted by Poria Cocos polysaccharides via TLR4/TRAF6/NF-kappaB signaling in vitro and in vivo. Biomed. Pharmacother. 2019, 112, 108709. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.; Fang, S.; Cui, X.; Guan, R.; Wang, Y.; Shi, F.; Hu, S. Signaling pathway underlying splenocytes activation by polysaccharides from Atractylodis macrocephalae Koidz. Mol. Immunol. 2019, 111, 19–26. [Google Scholar] [CrossRef] [PubMed]
- Beini, S.; Shuang, Y.; Dongying, Z.; Sihan, G.; Xiaohua, W.; Kai, Z. Polysaccharides as vaccine adjuvants. Vaccine 2018, 36, 5226–5234. [Google Scholar] [CrossRef]
- Barclay, T.G.; Day, C.M.; Petrovsky, N.; Garg, S. Review of polysaccharide particle-based functional drug delivery. Carbohydr. Polym. 2019, 221, 94–112. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Liu, D.; Fang, L.; Zhao, X.; Zhou, A.; Xie, J. A galactomannoglucan derived from Agaricus brasiliensis: Purification, characterization and macrophage activation via MAPK and IkappaB/NFkappaB pathways. Food Chem. 2018, 239, 603–611. [Google Scholar] [CrossRef]
- Coquerelle, C.; Moser, M. DC subsets in positive and negative regulation of immunity. Immunol. Rev. 2010, 234, 317–334. [Google Scholar] [CrossRef]
- Lanzavecchia, A.; Sallusto, F. Regulation of T cell immunity by dendritic cells. Cell 2001, 106, 263–266. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Li, H.; Li, M.; Niu, S.; Wang, J.; Shao, H.; Li, T.; Wang, H. Salvia miltiorrhiza polysaccharide activates T Lymphocytes of cancer patients through activation of TLRs mediated -MAPK and -NF-kappaB signaling pathways. J. Ethnopharmacol. 2017, 200, 165–173. [Google Scholar] [CrossRef]
- Wang, H.; Yu, Q.; Nie, S.P.; Xiang, Q.D.; Zhao, M.M.; Liu, S.Y.; Xie, M.Y.; Wang, S.Q. Polysaccharide purified from Ganoderma atrum induced activation and maturation of murine myeloid-derived dendritic cells. Food Chem. Toxicol. 2017, 108, 478–485. [Google Scholar] [CrossRef]
- Zhang, W.; Ma, W.; Zhang, J.; Song, X.; Sun, W.; Fan, Y. The immunoregulatory activities of astragalus polysaccharide liposome on macrophages and dendritic cells. Int. J. Biol. Macromol. 2017, 105, 852–861. [Google Scholar] [CrossRef]
- Park, S.-H.; Kim, M.S.; Lim, H.X.; Cho, D.; Kim, T.S. IL-33-matured dendritic cells promote Th17 cell responses via IL-1β and IL-6. Cytokine 2017, 99, 106–113. [Google Scholar] [CrossRef] [PubMed]
- Minato, K.I.; Laan, L.C.; Ohara, A.; van Die, I. Pleurotus citrinopileatus polysaccharide induces activation of human dendritic cells through multiple pathways. Int. Immunopharmacol. 2016, 40, 156–163. [Google Scholar] [CrossRef] [PubMed]
- Lai, C.; Yang, L.; Lin, W. Type II arabinogalactan from Anoectochilus formosanus induced dendritic cell maturation through TLR2 and TLR4. Phytomed. Int. J. Phytother. Phytopharm. 2015, 22, 1207–1214. [Google Scholar] [CrossRef]
- Kawasaki, T.; Kawai, T. Toll-like receptor signaling pathways. Front. Immunol. 2014, 5, 461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Premkumar, V.; Dey, M.; Dorn, R.; Raskin, I. MyD88-dependent and independent pathways of Toll-Like Receptors are engaged in biological activity of Triptolide in ligand-stimulated macrophages. BMC Chem. Biol. 2010, 10, 3. [Google Scholar] [CrossRef] [Green Version]









© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, Y.; Wang, D.; Li, Q.; He, J.; Wang, B.; Li, J.; Zhang, A. Immune-Enhancing Activity of Aqueous Extracts from Artemisia rupestris L. via MAPK and NF-kB Pathways of TLR4/TLR2 Downstream in Dendritic Cells. Vaccines 2020, 8, 525. https://doi.org/10.3390/vaccines8030525
Yang Y, Wang D, Li Q, He J, Wang B, Li J, Zhang A. Immune-Enhancing Activity of Aqueous Extracts from Artemisia rupestris L. via MAPK and NF-kB Pathways of TLR4/TLR2 Downstream in Dendritic Cells. Vaccines. 2020; 8(3):525. https://doi.org/10.3390/vaccines8030525
Chicago/Turabian StyleYang, Yu, DanYang Wang, QuanXiao Li, Jiang He, Bin Wang, Jinyao Li, and Ailian Zhang. 2020. "Immune-Enhancing Activity of Aqueous Extracts from Artemisia rupestris L. via MAPK and NF-kB Pathways of TLR4/TLR2 Downstream in Dendritic Cells" Vaccines 8, no. 3: 525. https://doi.org/10.3390/vaccines8030525
APA StyleYang, Y., Wang, D., Li, Q., He, J., Wang, B., Li, J., & Zhang, A. (2020). Immune-Enhancing Activity of Aqueous Extracts from Artemisia rupestris L. via MAPK and NF-kB Pathways of TLR4/TLR2 Downstream in Dendritic Cells. Vaccines, 8(3), 525. https://doi.org/10.3390/vaccines8030525