The Combination of Molecular Adjuvant CCL35.2 and DNA Vaccine Significantly Enhances the Immune Protection of Carassius auratus gibelio against CyHV-2 Infection



Abstract
:1. Introduction
2. Materials and Methods
2.1. Fish, Virus and Cells
2.2. Preparation of Inactivated Vaccine, DNA Vaccine and Molecular Adjuvant
2.3. Expression of Recombinant pcORF25 and pcCCL35.2 in FHM Cells
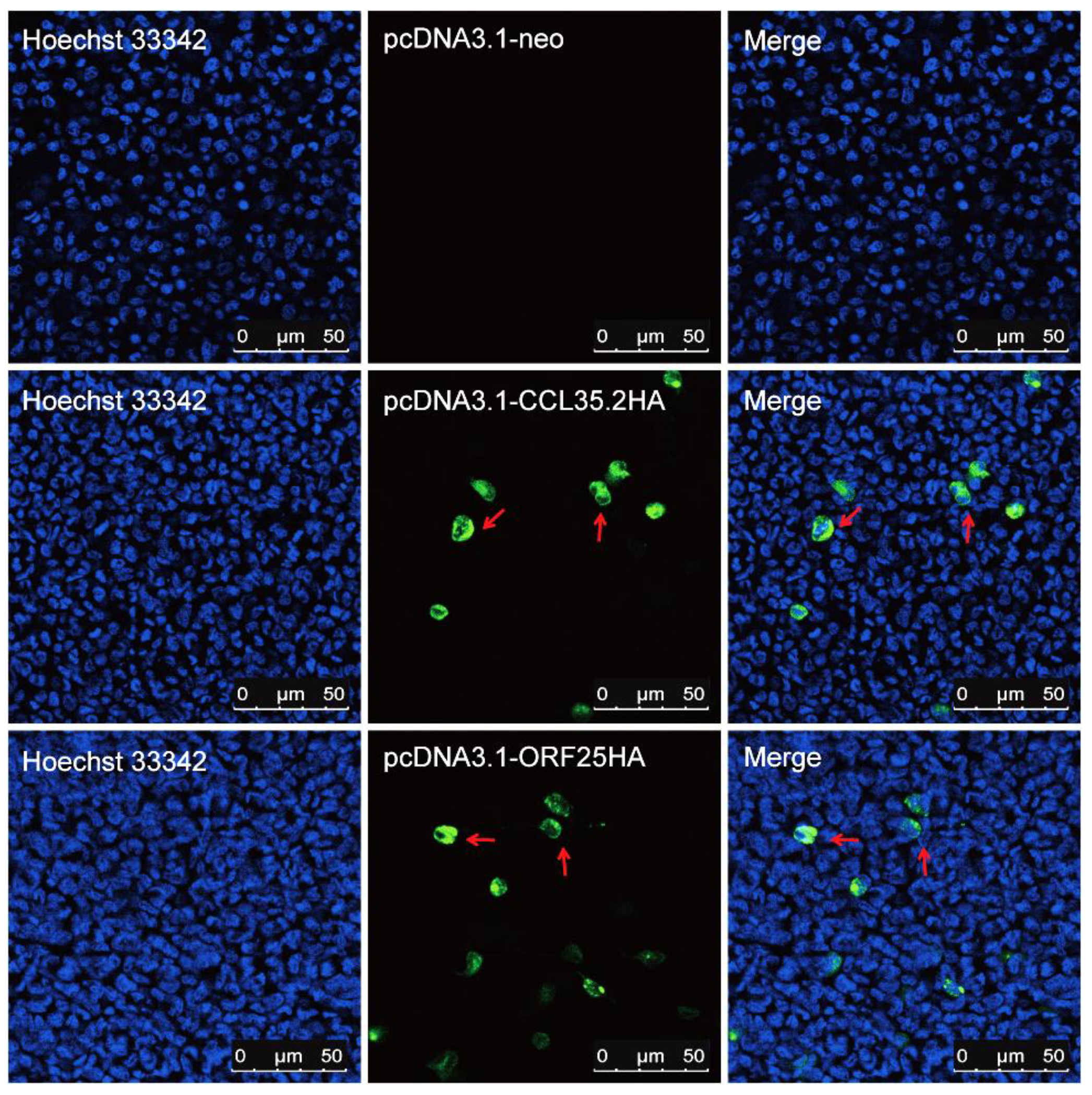
2.4. Expression of Recombinant pcORF25 and pcCCL35.2 in Muscle Tissue
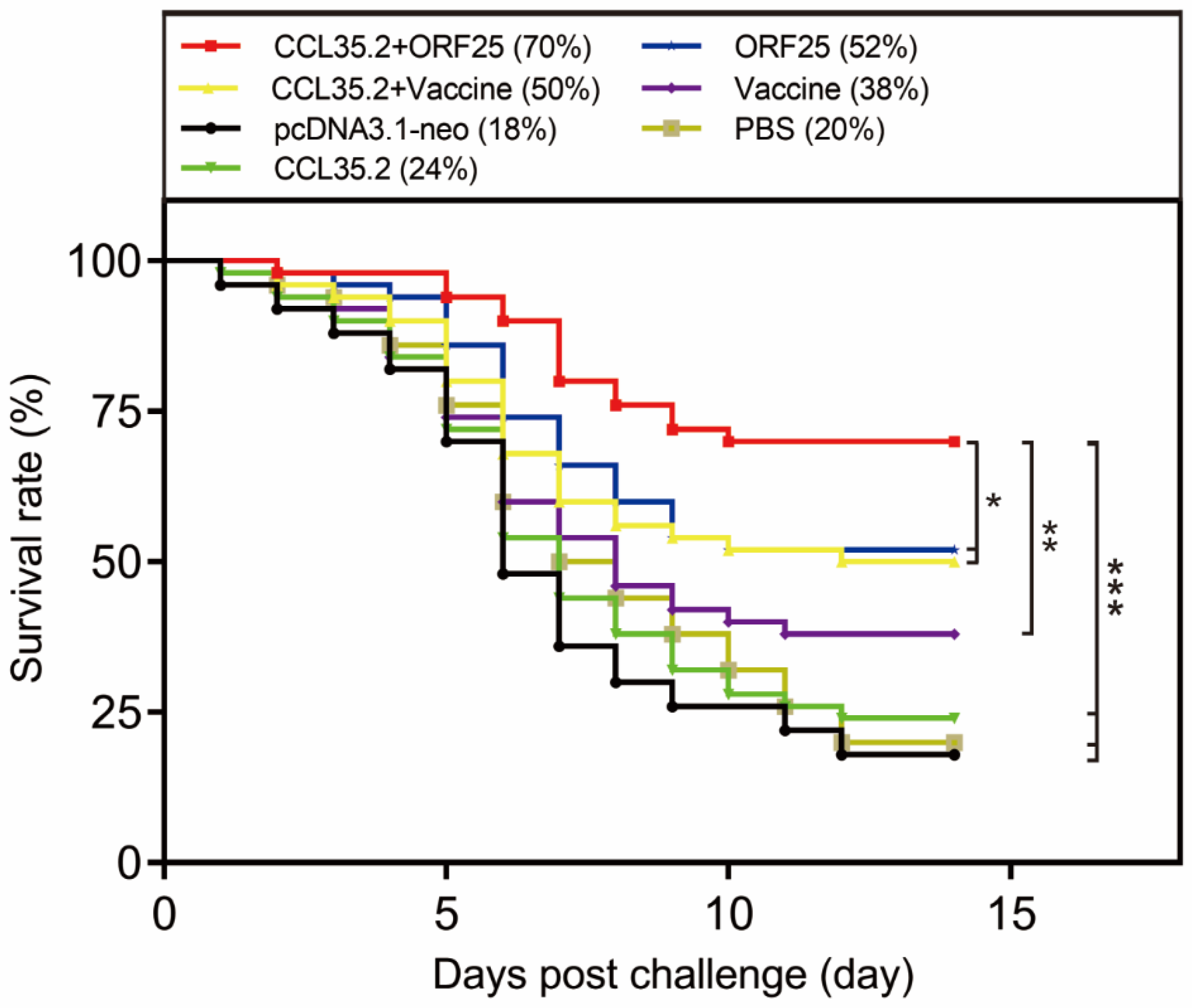
2.5. Vaccination, Challenge, Sampling and Survival Assay
2.6. Serum Antibody Titer by ELISA Assay
2.7. Serum Antibody Neutralization Tests and Biochemistry Index Assays
2.8. Immune Genes and Virus Gene for qRT-PCR
2.9. Histopathological Examination
2.10. Statistical Analysis
3. Results
3.1. Recombinant pcORF25 and pcCCL35.2 Can Express Corresponding Proteins In Vitro and In Vivo
3.2. pcORF25/pcCCL35.2 Mixture Remarkably Improves the Survival Rate of Gibel Carp
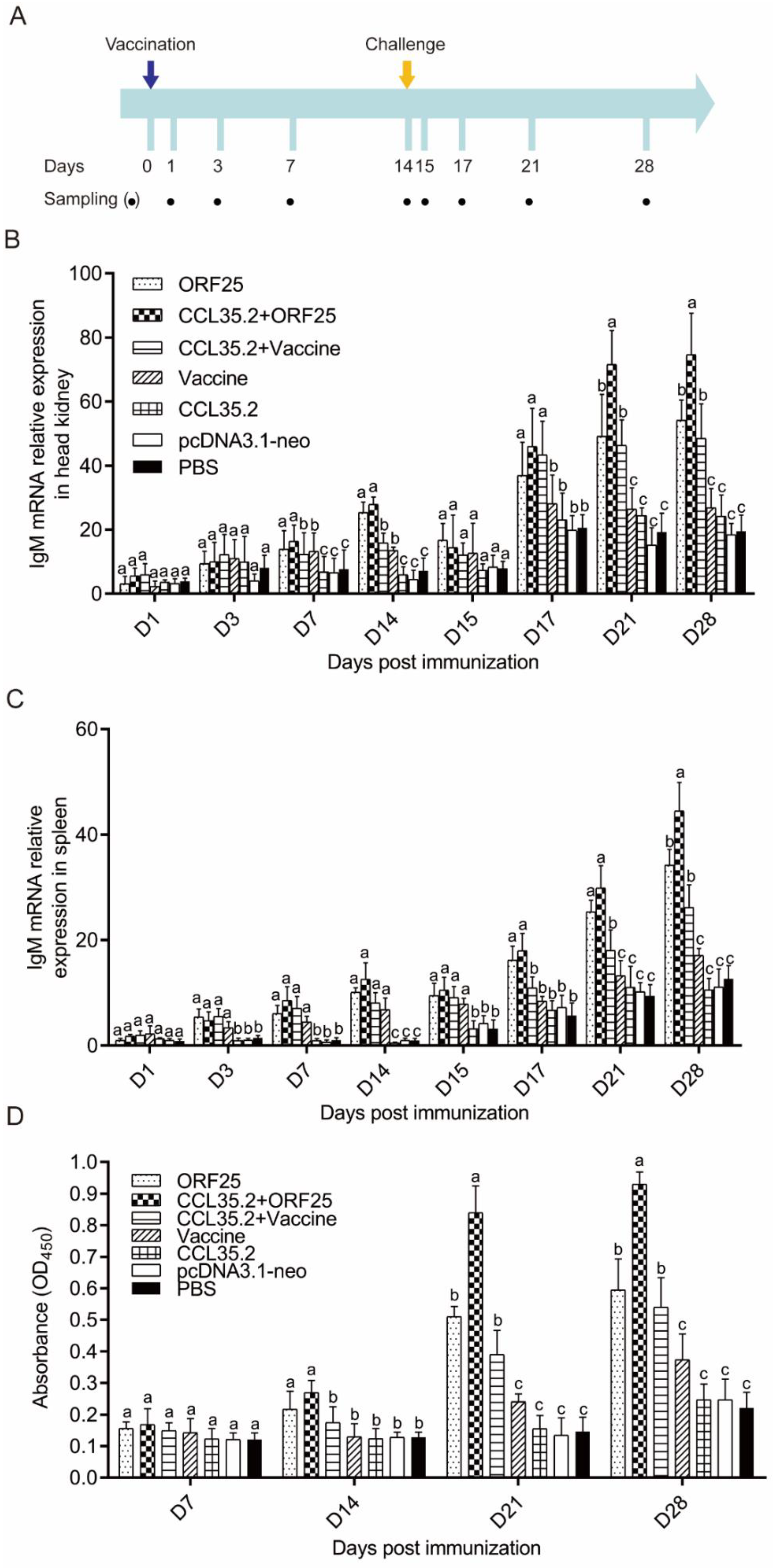
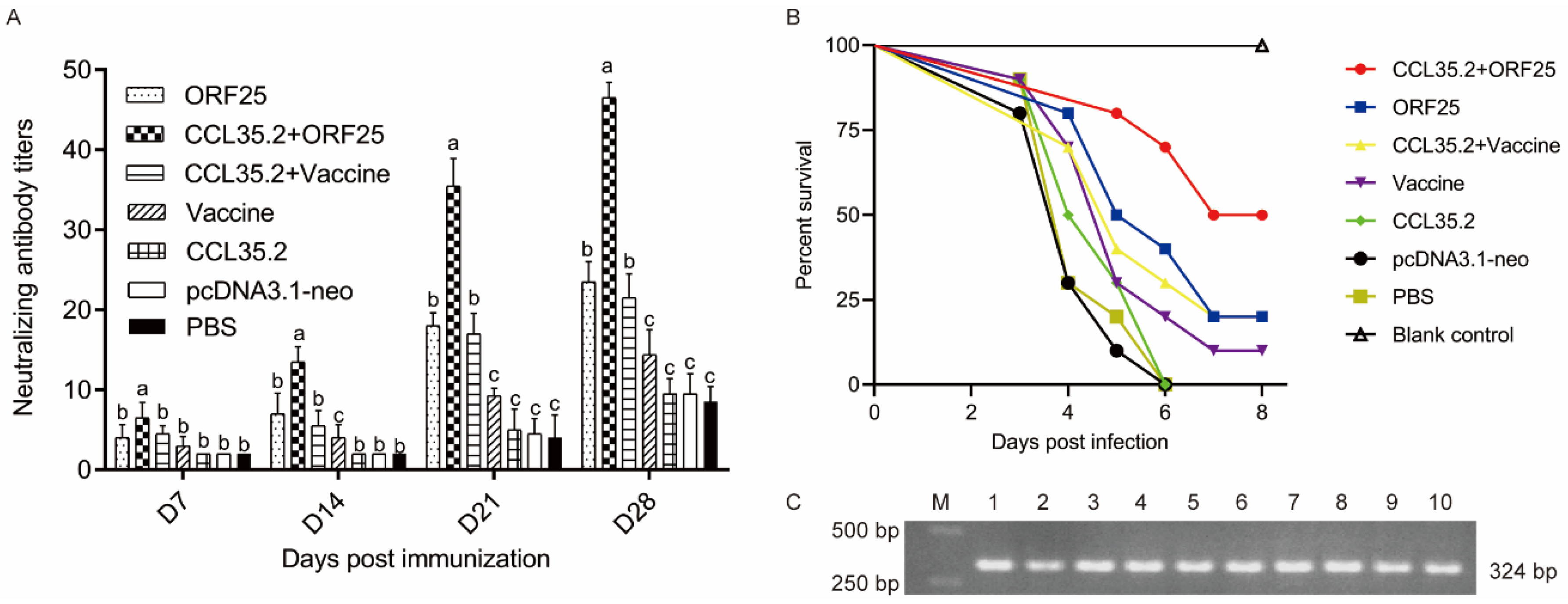
3.3. pcORF25/pcCCL35.2 Significantly Enhances Specific Antibody Levels and Neutralization Capacity
3.4. Improvement of Serum Innate Immune Indices
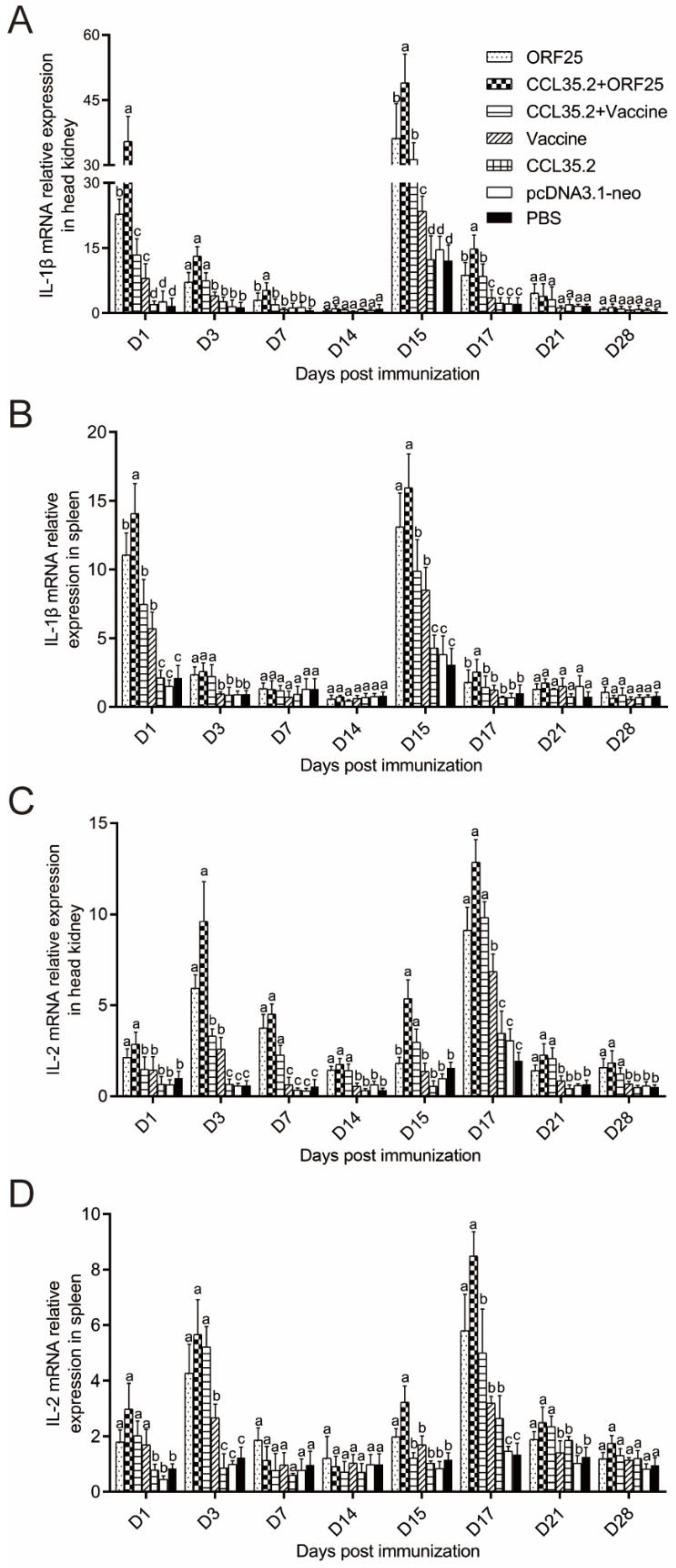
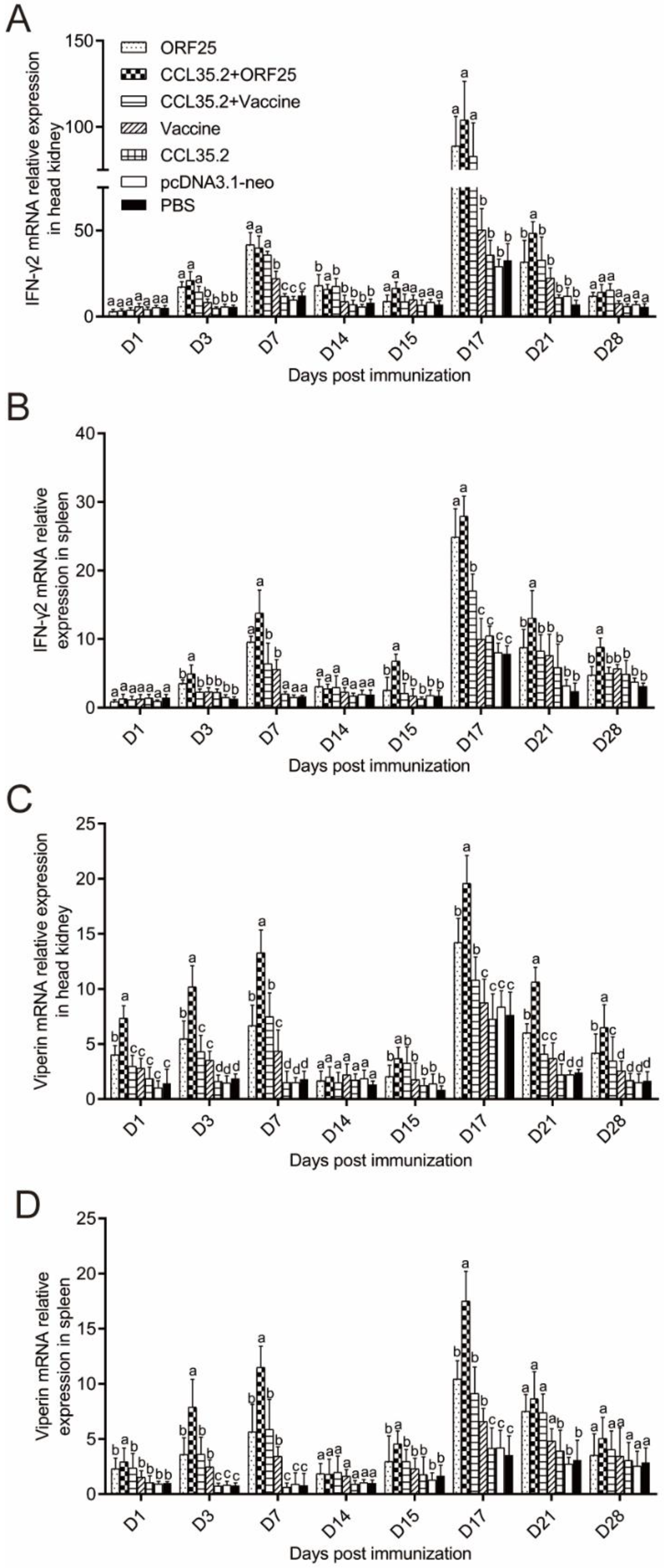
3.5. Immune Gene Expression was Significantly Upregulated by pcORF25/pcCCL35.2 Treatment
3.6. pcORF25/pcCCL35.2 Treatment Effectively Relieves the Tissue Lesions against Virus Infection
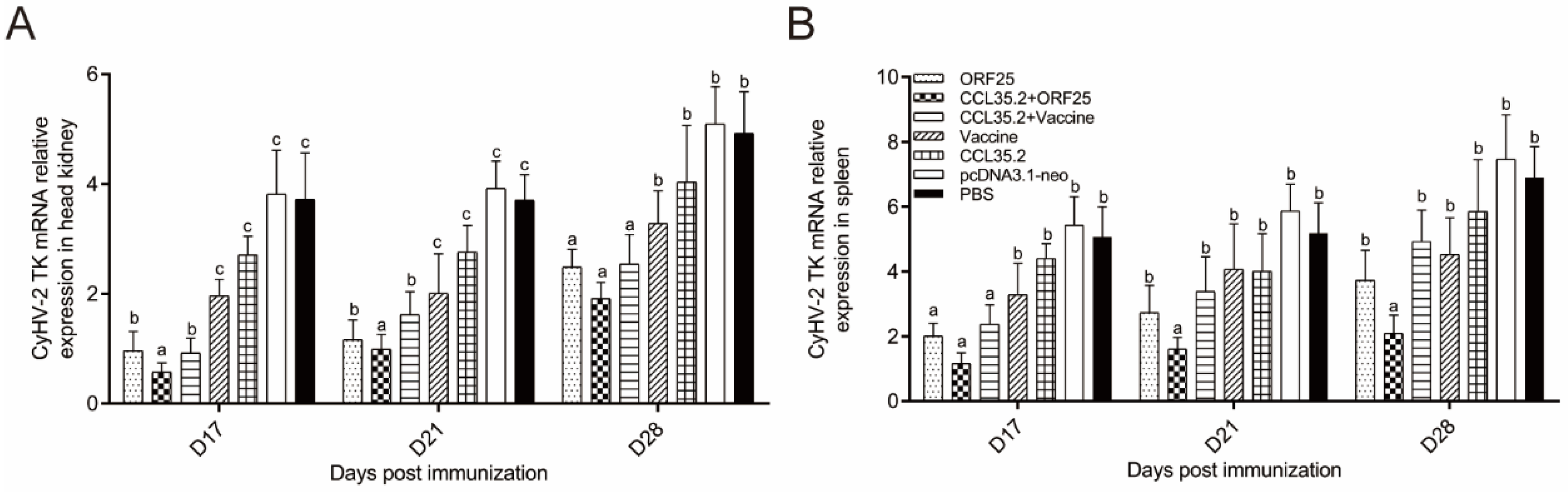
3.7. Virus Replication Was Suppressed in CCL35.2/ORF25 Group
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Gui, J.; Zhou, L. Genetic basis and breeding application of clonal diversity and dual reproduction modes in polyploid Carassius auratus gibelio. Sci. China Life Sci. 2010, 53, 409–415. [Google Scholar] [CrossRef] [PubMed]
- Goodwin, A.E.; Sadler, J.; Merry, G.E.; Marecaux, E.N. Herpesviral haematopoietic necrosis virus (CyHV-2) infection: Case studies from commercial goldfish farms. J. Fish Dis. 2009, 32, 271–278. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Zeng, L.; Zhang, H.; Zhou, Y.; Ma, J.; Fan, Y. Cyprinid herpesvirus 2 infection emerged in cultured gibel carp, Carassius auratus gibelio in China. Vet. Microbiol. 2013, 166, 138–144. [Google Scholar] [CrossRef] [PubMed]
- Wei, C.; Iida, H.; Chuah, Q.; Tanaka, M.; Kato, G.; Sano, M. Persistence of cyprinid herpesvirus 2 in asymptomatic goldfish Carassius auratus (L.) that survived an experimental infection. J. Fish Dis. 2019, 42, 913–921. [Google Scholar] [CrossRef] [PubMed]
- Yuan, X.; Shen, J.; Pan, X.; Yao, J.; Lyu, S.; Liu, L.; Zhang, H. Screening for protective antigens of Cyprinid herpesvirus 2 and construction of DNA vaccines. J. Virol. Methods 2020, 280, 113877. [Google Scholar] [CrossRef]
- Zhou, Y.; Jiang, N.; Ma, J.; Fan, Y.; Zhang, L.; Xu, J.; Zeng, L. Protective immunity in gibel carp, Carassius gibelio of the truncated proteins of Cyprinid herpesvirus 2 expressed in Pichia pastoris. Fish Shellfish Immunol. 2015, 47, 1024–1031. [Google Scholar] [CrossRef]
- Liu, L.; Gao, S.; Luan, W.; Zhou, J.; Wang, H. Generation and functional evaluation of a DNA vaccine co-expressing Cyprinid herpesvirus-3 envelope protein and carp interleukin-1 beta. Fish Shellfish Immunol. 2018, 80, 223–231. [Google Scholar] [CrossRef]
- Su, H.; Su, J. Cyprinid viral diseases and vaccine development. Fish Shellfish Immunol. 2018, 83, 84–95. [Google Scholar] [CrossRef]
- Suschak, J.J.; Williams, J.A.; Schmaljohn, C.S. Advancements in DNA vaccine vectors, non-mechanical delivery methods, and molecular adjuvants to increase immunogenicity. Hum. Vaccin. Immunother. 2017, 13, 2837–2848. [Google Scholar] [CrossRef] [Green Version]
- Kutzler, M.A.; Weiner, D.B. DNA vaccines: Ready for prime time? Nat. Rev. Genet. 2008, 9, 776–788. [Google Scholar] [CrossRef]
- Pal, R.; Yu, Q.; Wang, S.; Kalyanaraman, V.S.; Nair, B.C.; Hudacik, L.; Whitney, S.; Keen, T.; Hung, C.L.; Hocker, L.; et al. Definitive toxicology and biodistribution study of a polyvalent DNA prime/protein boost human immunodeficiency virus type 1 (HIV-1) vaccine in rabbits. Vaccine 2006, 24, 1225–1234. [Google Scholar] [CrossRef] [PubMed]
- Sheets, R.L.; Stein, J.; Manetz, T.S.; Duffy, C.; Nason, M.; Andrews, C.; Kong, W.; Nabel, G.J.; Gomez, P.L. Biodistribution of DNA plasmid vaccines against HIV-1, Ebola, severe acute respiratory syndrome, or west nile virus is similar, without integration, despite differing plasmid backbones or gene inserts. Toxicol. Sci. 2006, 91, 610–619. [Google Scholar] [CrossRef]
- Chapman, R.; Rybicki, E.P. Use of a novel enhanced DNA vaccine vector for preclinical virus vaccine investigation. Vaccines 2019, 7, 50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Das, S.; Ramakrishnan, K.; Behera, S.K.; Ganesapandian, M.; Xavier, A.S.; Selvarajan, S. Hepatitis B vaccine and immunoglobulin: Key concepts. J. Clin. Transl. Hepatol. 2019, 7, 165–171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garza-Morales, R.; Perez-Trujillo, J.J.; Martinez-Jaramillo, E.; Saucedo-Cardenas, O.; Loera-Arias, M.J.; Garcia-Garcia, A.; Rodriguez-Rocha, H.; Yolcu, E.; Shirwan, H.; Gomez-Gutierrez, J.G.; et al. A DNA vaccine encoding SA-4-1BBL fused to HPV-16 E7 antigen has prophylactic and therapeutic efficacy in a cervical cancer mouse model. Cancers 2019, 11, 96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saade, F.; Petrovsky, N. Technologies for enhanced efficacy of DNA vaccines. Expert Rev. Vaccines 2012, 11, 189–209. [Google Scholar] [CrossRef]
- Xu, H.; Xing, J.; Tang, X.; Sheng, X.; Zhan, W. The effects of CCL3, CCL4, CCL19 and CCL21 as molecular adjuvants on the immune response to VAA DNA vaccine in flounder (Paralichthys olivaceus). Dev. Comp. Immunol. 2020, 103, 103492. [Google Scholar] [CrossRef]
- Miller, M.C.; Mayo, K.H. Chemokines from a Structural Perspective. Int. J. Mol. Sci. 2017, 18, 88. [Google Scholar] [CrossRef] [Green Version]
- Bystry, R.S.; Aluvihare, V.; Welch, K.A.; Kallikourdis, M.; Betz, A.G. B cells and professional APCs recruit regulatory T cells via CCL4. Nat. Immunol. 2001, 2, 1126–1132. [Google Scholar] [CrossRef]
- Nguyen-Hoai, T.; Pham-Duc, M.; Gries, M.; Dorken, B.; Pezzutto, A.; Westermann, J. CCL4 as an adjuvant for DNA vaccination in a Her2/neu mouse tumor model. Cancer Gene Ther. 2016, 23, 162–167. [Google Scholar] [CrossRef]
- Lu, M.; Kim, W.H.; Lillehoj, H.S.; Li, C. Development and characterization of novel mouse monoclonal antibodies against chicken chemokine CC motif ligand 4. Vet. Immunol. Immunopathol. 2020, 227, 110091. [Google Scholar] [CrossRef] [PubMed]
- Lillard, J.W., Jr.; Singh, U.P.; Boyaka, P.N.; Singh, S.; Taub, D.D.; McGhee, J.R. MIP-1alpha and MIP-1beta differentially mediate mucosal and systemic adaptive immunity. Blood 2003, 101, 807–814. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liao, Z.; Wan, Q.; Xiao, X.; Ji, J.; Su, J. A systematic investigation on the composition, evolution and expression characteristics of chemokine superfamily in grass carp Ctenopharyngodon idella. Dev. Comp. Immunol. 2018, 82, 72–82. [Google Scholar] [CrossRef] [PubMed]
- Yan, Y.; Huo, X.; Ai, T.; Su, J. β-glucan and anisodamine can enhance the immersion immune efficacy of inactivated Cyprinid herpesvirus 2 vaccine in Carassius auratus gibelio. Fish Shellfish Immunol. 2020, 98, 285–295. [Google Scholar] [CrossRef]
- Yao, Q.; Qian, P.; Huang, Q.; Cao, Y.; Chen, H. Comparison of immune responses to different foot-and-mouth disease genetically engineered vaccines in guinea pigs. J. Virol. Methods 2008, 147, 143–150. [Google Scholar] [CrossRef]
- Tonheim, T.C.; Bogwald, J.; Dalmo, R.A. What happens to the DNA vaccine in fish? A review of current knowledge. Fish Shellfish Immunol. 2008, 25, 1–18. [Google Scholar] [CrossRef]
- Collins, C.; Lorenzen, N.; Collet, B. DNA vaccination for finfish aquaculture. Fish Shellfish Immunol. 2019, 85, 106–125. [Google Scholar] [CrossRef]
- Pinto, A.R.; Reyes-Sandoval, A.; Ertl, H.C. Chemokines and trance as genetic adjuvants for a DNA vaccine to rabies virus. Cell. Immunol. 2003, 224, 106–113. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, B.; Sun, L. P247 and p523: Two in vivo-expressed megalocytivirus proteins that induce protective immunity and are essential to viral infection. PLoS ONE 2015, 10, e0121282. [Google Scholar] [CrossRef]
- Chang, C.; Gu, J.; Robertsen, B. Protective effect and antibody response of DNA vaccine against salmonid alphavirus 3 (SAV3) in Atlantic salmon. J. Fish Dis. 2017, 40, 1775–1781. [Google Scholar] [CrossRef]
- Ito, T.; Maeno, Y. Effect of booster shot and investigation of vaccination efficacy period against herpesviral haematopoietic necrosis (HVHN) in goldfish Carassius auratus. Vet. Microbiol. 2015, 175, 139–144. [Google Scholar] [CrossRef] [PubMed]
- Zwollo, P.; Cole, S.; Bromage, E.; Kaattari, S. B cell heterogeneity in the teleost kidney: Evidence for a maturation gradient from anterior to posterior kidney. J. Immunol. 2005, 174, 6608–6616. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Wit, J.; Souwer, Y.; Jorritsma, T.; Klaasse Bos, H.; ten Brinke, A.; Neefjes, J.; van Ham, S.M. Antigen-specific B cells reactivate an effective cytotoxic T cell response against phagocytosed Salmonella through cross-presentation. PLoS ONE 2010, 5, e13016. [Google Scholar] [CrossRef] [PubMed]
- Zhu, B.; Zhang, C.; Zhao, Z.; Wang, G. Targeted delivery of mannosylated nanoparticles improve prophylactic efficacy of immersion vaccine against fish viral disease. Vaccines 2020, 8, 87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiu, S.T.; Tsai, R.T.; Hsu, J.P.; Liu, C.; Cheng, W. Dietary sodium alginate administration to enhance the non-specific immune responses, and disease resistance of the juvenile grouper Epinephelus fuscoguttatus. Aquaculture 2008, 277, 66–72. [Google Scholar] [CrossRef]
- Boshra, H.; Li, J.; Sunyer, J.O. Recent advances on the complement system of teleost fish. Fish Shellfish Immunol. 2006, 20, 239–262. [Google Scholar] [CrossRef]
- Zhu, W.; Zhang, Y.; Zhang, J.; Yuan, G.; Liu, X.; Ai, T.; Su, J. Astragalus polysaccharides, chitosan and poly(I:C) obviously enhance inactivated Edwardsiella ictaluri vaccine potency in yellow catfish Pelteobagrus fulvidraco. Fish Shellfish Immunol. 2019, 87, 379–385. [Google Scholar] [CrossRef]
- Garlanda, C.; Dinarello, C.A.; Mantovani, A. The interleukin-1 family: Back to the future. Immunity 2013, 39, 1003–1018. [Google Scholar] [CrossRef] [Green Version]
- Wang, T.; Hu, Y.; Wangkahart, E.; Liu, F.; Wang, A.; Zahran, E.; Maisey, K.R.; Liu, M.; Xu, Q.; Imarai, M.; et al. Interleukin (IL)-2 Is a key regulator of T helper 1 and T helper 2 cytokine expression in fish: Functional characterization of two divergent IL2 paralogs in Salmonids. Front. Immunol. 2018, 9, 1683. [Google Scholar] [CrossRef] [Green Version]
- Yu, T.K.; Caudell, E.G.; Smid, C.; Grimm, E.A. IL-2 activation of NK cells: Involvement of MKK1/2/ERK but not p38 kinase pathway. J. Immunol. 2000, 164, 6244–6251. [Google Scholar] [CrossRef] [Green Version]
- Diaz-Rosales, P.; Bird, S.; Wang, T.; Fujiki, K.; Davidson, W.S.; Zou, J.; Secombes, C.J. Rainbow trout interleukin-2: Cloning, expression and bioactivity analysis. Fish Shellfish Immunol. 2009, 27, 414–422. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Zhang, Y.; Liu, T.; Shi, J.; Sun, F.; Gui, J. Fish viperin exerts a conserved antiviral function through RLR-triggered IFN signaling pathway. Dev. Comp. Immunol. 2014, 47, 140–149. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Zhang, J.; Xiao, Z.; Sun, L. Rock bream (Oplegnathus fasciatus) viperin is a virus-responsive protein that modulates innate immunity and promotes resistance against megalocytivirus infection. Dev. Comp. Immunol. 2014, 45, 35–42. [Google Scholar] [CrossRef] [PubMed]
- Dang, W.; Zhang, M.; Hu, Y.H.; Sun, L. Differential regulation of Sciaenops ocellatus viperin expression by intracellular and extracellular bacterial pathogens. Fish Shellfish Immunol. 2010, 29, 264–270. [Google Scholar] [CrossRef] [PubMed]
- Ding, Z.; Xia, S.; Zhao, Z.; Xia, A.; Shen, M.; Tang, J.; Xue, H.; Geng, X.; Yuan, S. Histopathological characterization and fluorescence in situ hybridization of Cyprinid herpesvirus 2 in cultured Prussian carp, Carassius auratus gibelio in China. J. Virol. Methods 2014, 206, 76–83. [Google Scholar] [CrossRef]
- Wu, T.; Ding, Z.; Ren, M.; An, L.; Xiao, Z.; Liu, P.; Gu, W.; Meng, Q.; Wang, W. The histo—And ultra-pathological studies on a fatal disease of Prussian carp (Carassius gibelio) in mainland China associated with cyprinid herpesvirus 2 (CyHV-2). Aquaculture 2013, 412–413, 8–13. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Dilution | Number of Survivors | |||||
|---|---|---|---|---|---|---|---|
| 2 | 3 | 4 | 5 | 6 | 7 | ||
| CCL35.2/ORF25 | 1:20 | 10 | 10 | 10 | 8 | 5 | 5 |
| ORF25 | 1:20 | 10 | 10 | 8 | 5 | 4 | 2 |
| CCL35.2/Vaccine | 1:20 | 10 | 10 | 7 | 6 | 3 | 2 |
| Vaccine | 1:20 | 10 | 9 | 7 | 3 | 2 | 1 |
| CCL35.2 | 1:20 | 10 | 9 | 5 | 3 | 0 | 0 |
| pcDNA3.1-neo | 1:20 | 10 | 8 | 3 | 1 | 0 | 0 |
| PBS | 1:20 | 10 | 9 | 3 | 2 | 0 | 0 |
| Blank control | 1:20 | 10 | 10 | 10 | 10 | 10 | 10 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huo, X.; Fan, C.; Ai, T.; Su, J. The Combination of Molecular Adjuvant CCL35.2 and DNA Vaccine Significantly Enhances the Immune Protection of Carassius auratus gibelio against CyHV-2 Infection. Vaccines 2020, 8, 567. https://doi.org/10.3390/vaccines8040567
Huo X, Fan C, Ai T, Su J. The Combination of Molecular Adjuvant CCL35.2 and DNA Vaccine Significantly Enhances the Immune Protection of Carassius auratus gibelio against CyHV-2 Infection. Vaccines. 2020; 8(4):567. https://doi.org/10.3390/vaccines8040567
Chicago/Turabian StyleHuo, Xingchen, Chengjian Fan, Taoshan Ai, and Jianguo Su. 2020. "The Combination of Molecular Adjuvant CCL35.2 and DNA Vaccine Significantly Enhances the Immune Protection of Carassius auratus gibelio against CyHV-2 Infection" Vaccines 8, no. 4: 567. https://doi.org/10.3390/vaccines8040567
APA StyleHuo, X., Fan, C., Ai, T., & Su, J. (2020). The Combination of Molecular Adjuvant CCL35.2 and DNA Vaccine Significantly Enhances the Immune Protection of Carassius auratus gibelio against CyHV-2 Infection. Vaccines, 8(4), 567. https://doi.org/10.3390/vaccines8040567