Innate and Adaptive Immune Responses against Bordetella pertussis and Pseudomonas aeruginosa in a Murine Model of Mucosal Vaccination against Respiratory Infection


 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Growth
2.2. Vaccine Preparation
2.3. Vaccination with B. pertussis and P. aeruginosa Whole Cell Vaccines, Bacterial Challenge, and Euthanasia
2.4. Detection of Bacterial Load
2.5. Histology
2.6. Hematology
2.7. Flow Cytometry Analysis
2.8. Cytokine Analysis
2.9. Serology
2.10. Statistics
2.11. Animal Care and Use
3. Results
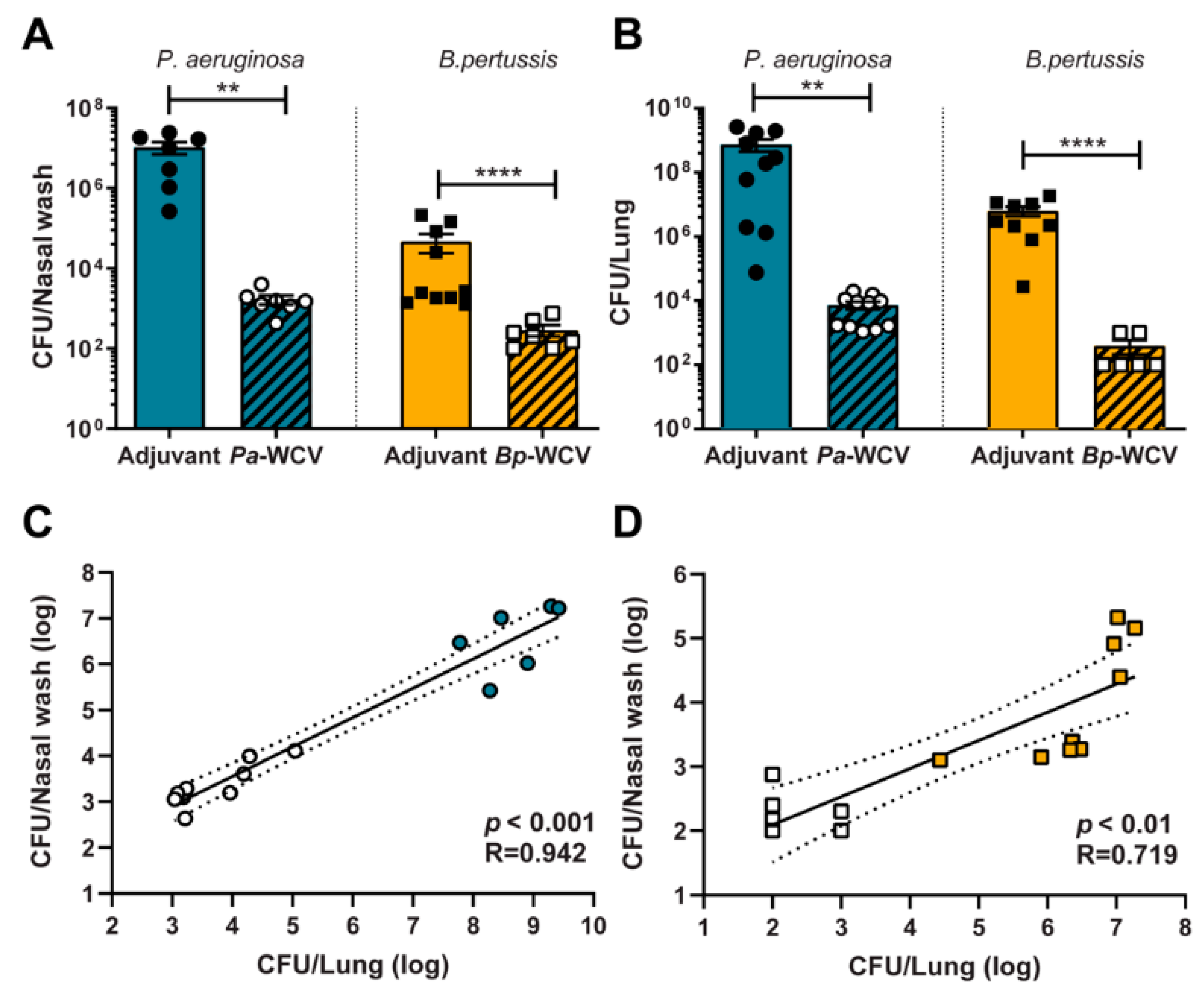
3.1. Whole Cell Intranasal Vaccination Against B. pertussis and P. aeruginosa Reduces Bacterial Colonization Following Challenge
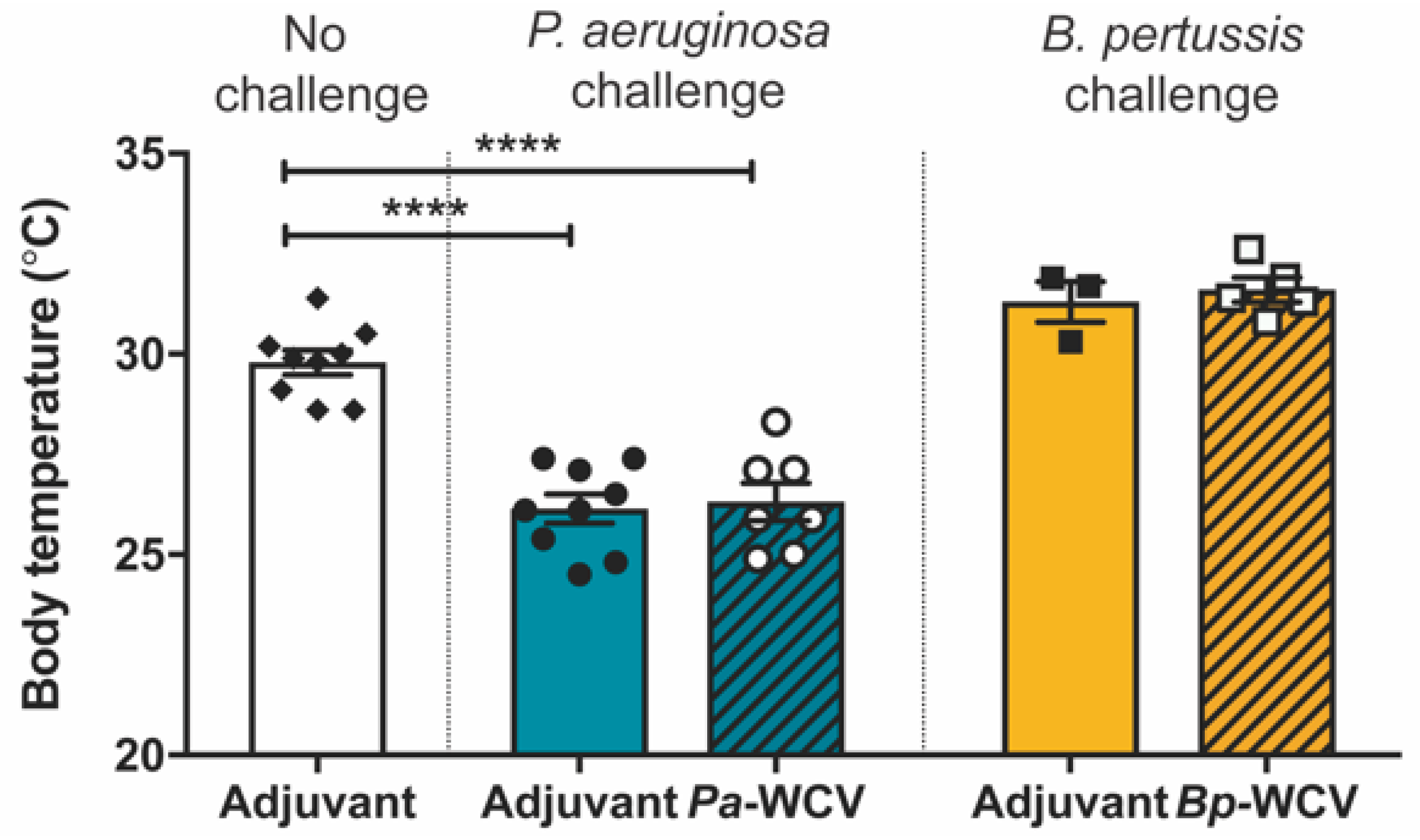
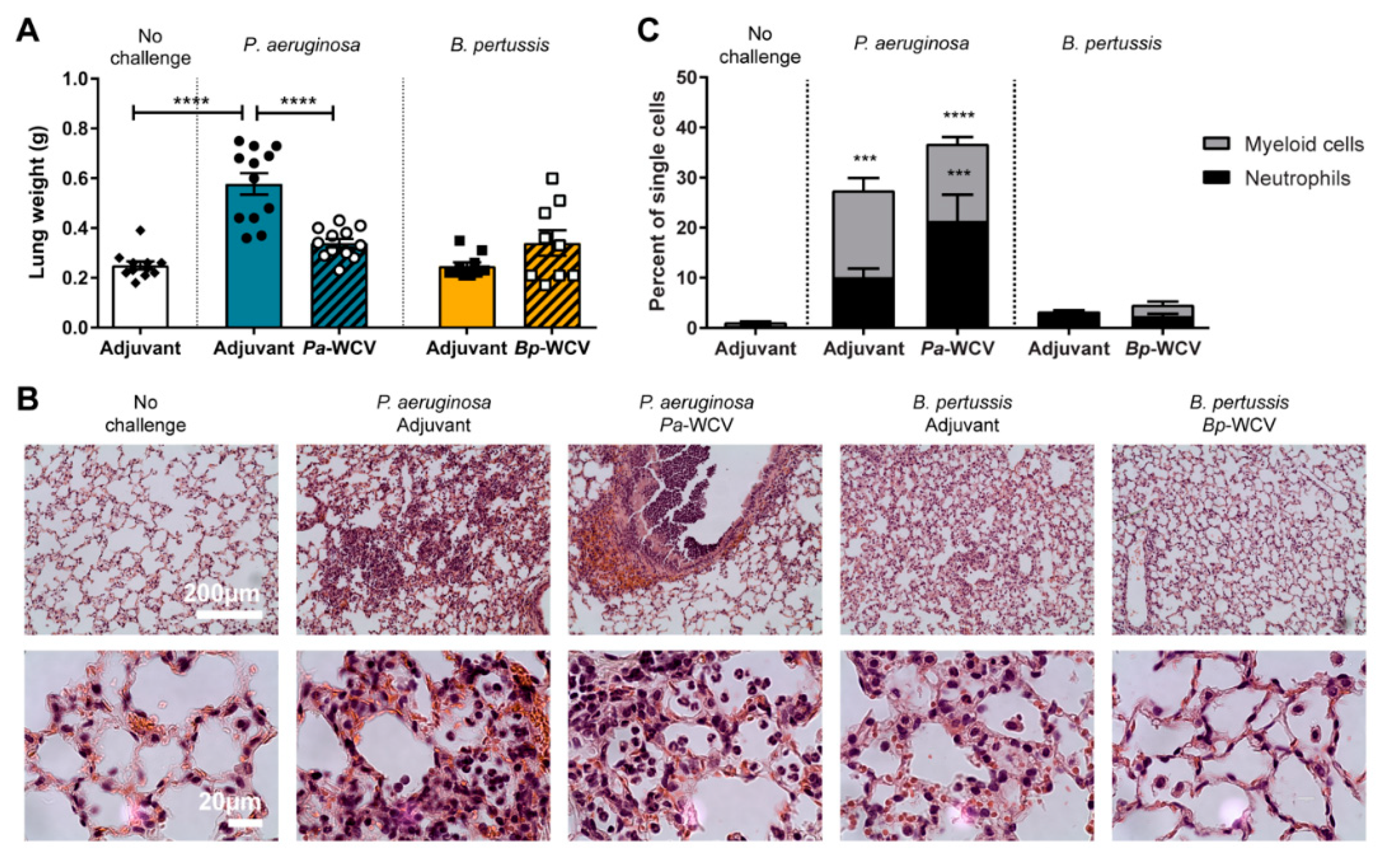
3.2. Innate Immune Response in the Lung Following Vaccination and Challenge
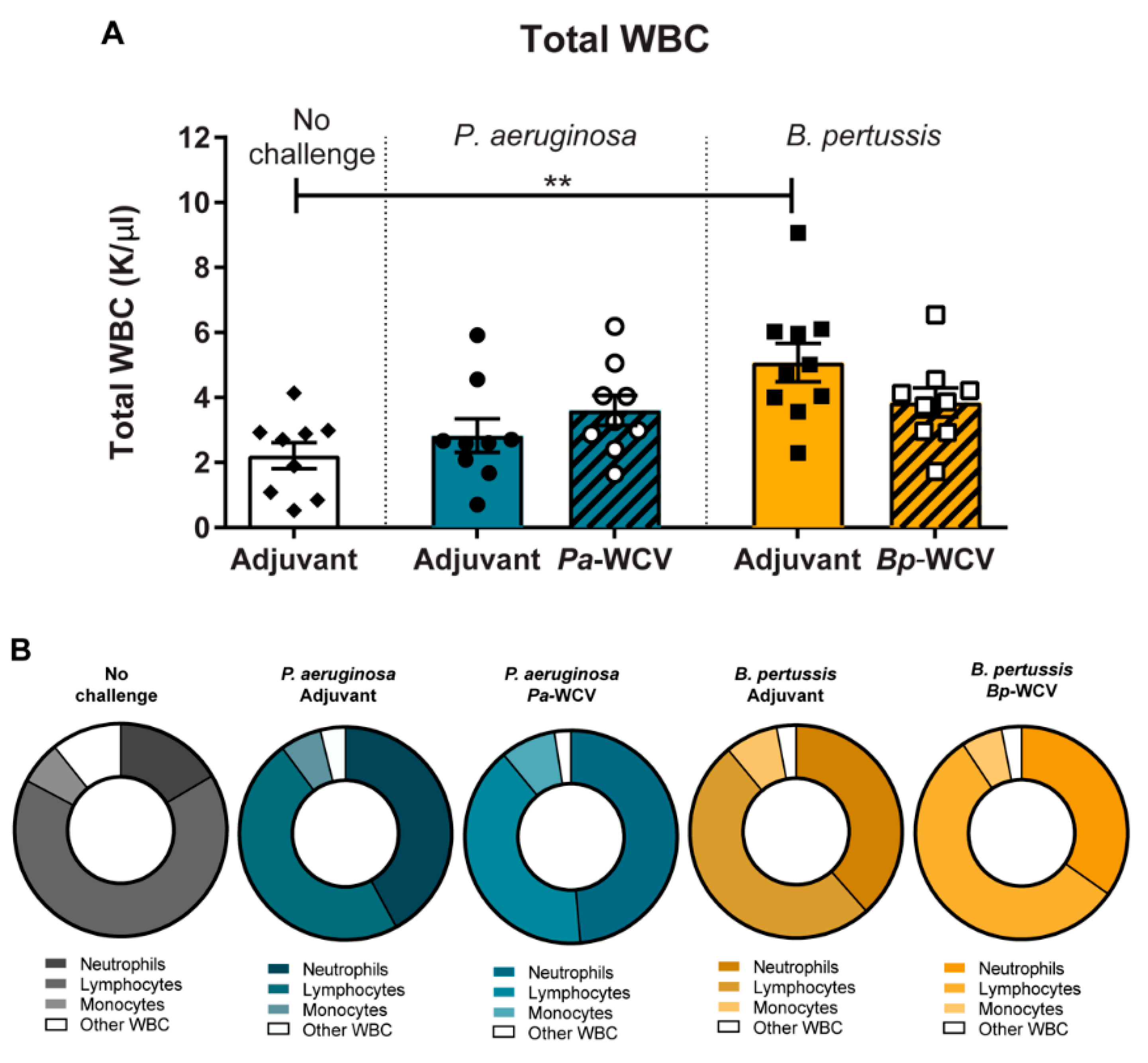
3.3. Leukocytosis Occurs During B. pertussis but Not P. aeruginosa Infection
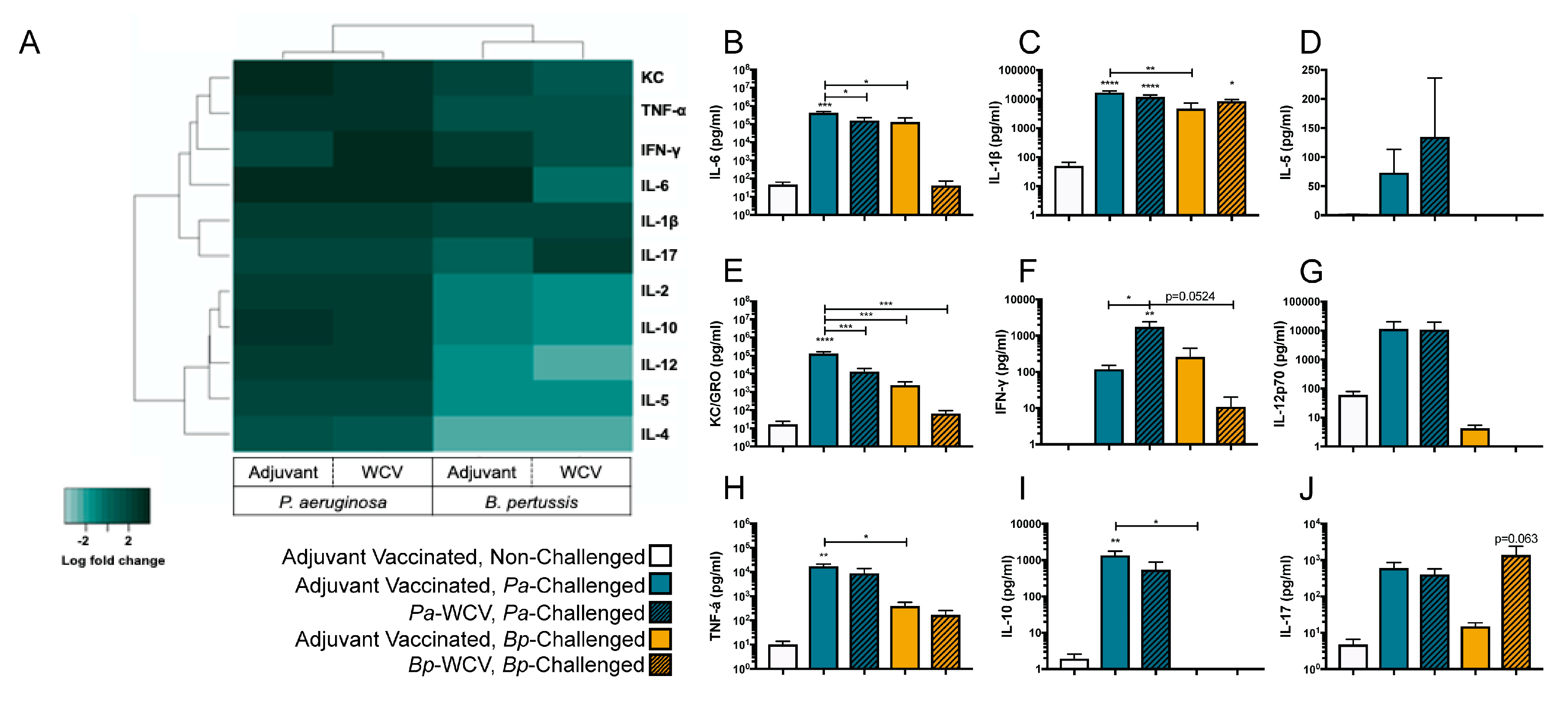
3.4. Vaccination Plays a Role in Modulating the Cytokine Response to B. pertussis but Not P. aeruginosa Infection
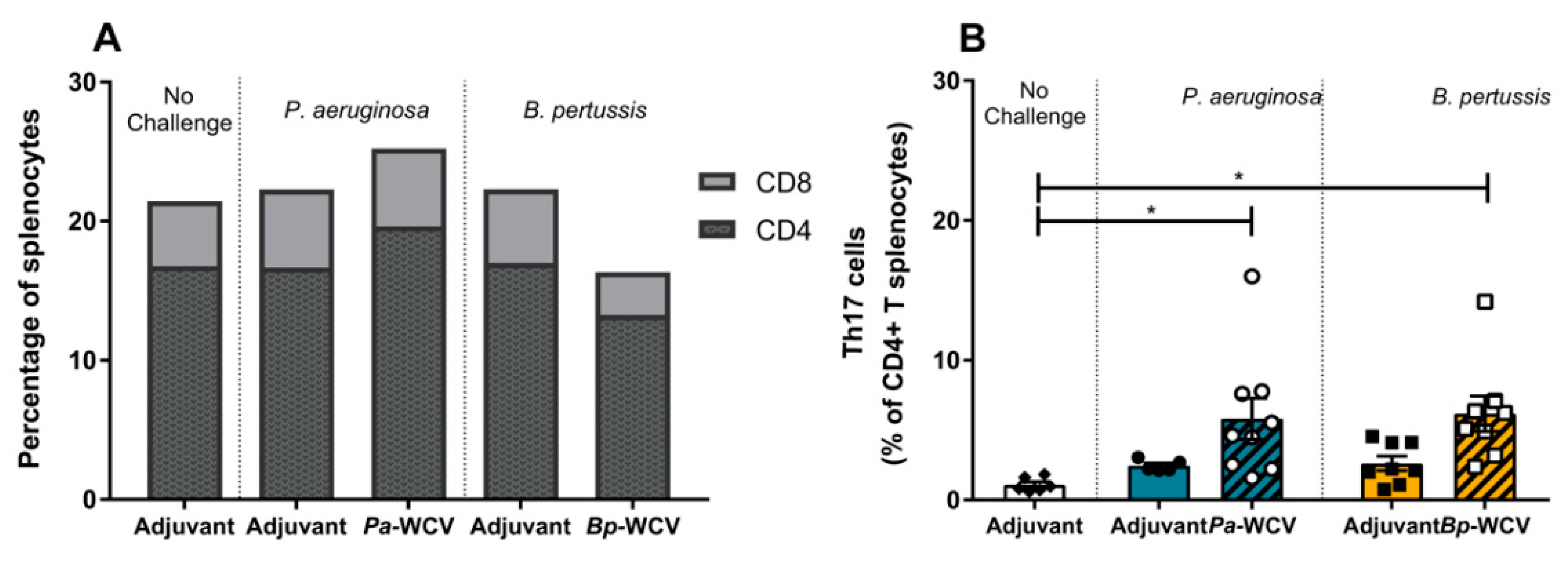
3.5. Whole Cell Vaccination Increases CD4+ Th17+ Cells in Spleen One Week Post Infection
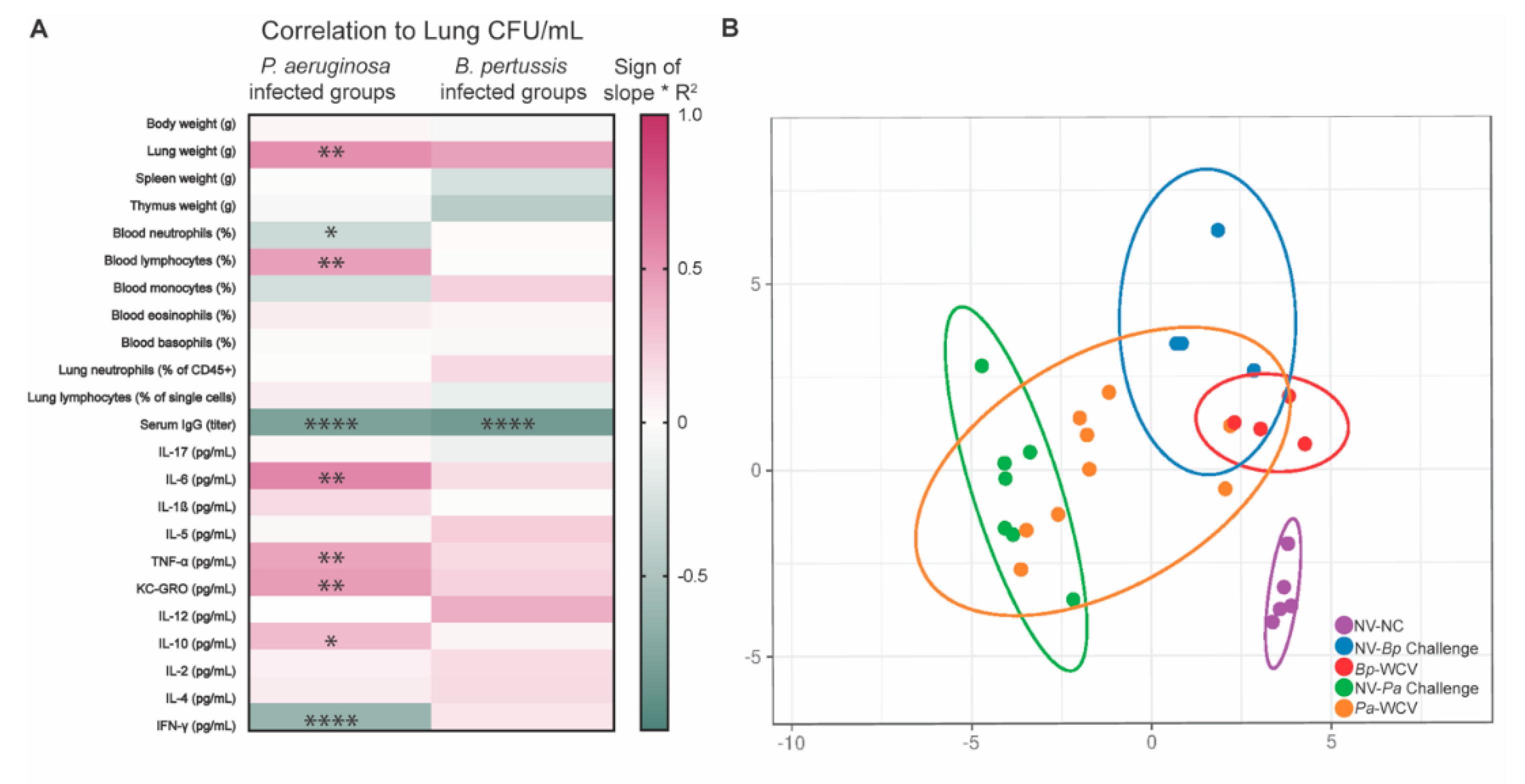
3.6. Correlates of Protection for B. pertussis and P. aeruginosa, Using Intranasal Whole Cell Vaccination as Protective Model
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- European Respiratory Society. The Global Impact of Respiratory Disease- Second Edition; Forum of International Respiratory Societies: Lausanne, Switzerland, 2017; ISBN 9781849840873. [Google Scholar]
- Pitts, S.R.; Niska, R.W.; Xu, J.; Burt, C.W. National Hospital Ambulatory Medical Care Survey: 2006 emergency department summary. Natl. Health Stat. Rep. 2008, 6, 1–38. [Google Scholar]
- Kochanek, K.D.; Murphy, S.L.; Xu, J.; Arias, E. Deaths: Final data for 2017. Natl. Vital Stat. Rep. 2019, 68, 1–18. [Google Scholar] [PubMed]
- Weiner, L.M.; Webb, A.K.; Limbago, B.; Dudeck, M.A.; Patel, J.; Kallen, A.J.; Edwards, J.R.; Sievert, D.M. Antimicrobial-Resistant Pathogens Associated with Healthcare- Associated Infections: Summary of Data Reported to the National Healthcare Safety Network at the Centers for Disease Control and Prevention, 2011–2014. Physiol. Behav. 2017, 176, 139–148. [Google Scholar] [CrossRef] [Green Version]
- Plotkin, S.A.; Orenstein, W.A.; Offit, P.A. Plotkin’s Vaccines; Elsevier: Amsterdam, The Netherlands, 2017; ISBN 9780323357616. [Google Scholar]
- Boucher, H.W.; Talbot, G.H.; Bradley, J.S.; Edwards, J.E.; Gilbert, D.; Rice, L.B.; Scheld, M.; Spellberg, B.; Bartlett, J. Bad Bugs, No Drugs: No ESKAPE! An Update from the Infectious Diseases Society of America. Clin. Infect. Dis. 2009, 48, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Yoon, S.S.; Hennigan, R.F.; Hilliard, G.M.; Ochsner, U.A.; Parvatiyar, K.; Kamani, M.C.; Allen, H.L.; DeKievit, T.R.; Gardner, P.R.; Schwab, U.; et al. Pseudomonas aeruginosa anaerobic respiration in biofilms: Relationships to cystic fibrosis pathogenesis. Dev. Cell 2002, 3, 593–603. [Google Scholar] [CrossRef] [Green Version]
- Gharabaghi, M.A.; Abdollahi, S.M.M.; Safavi, E.; Abtahi, S.H. Community acquired Pseudomonas pneumonia in an immune competent host. BMJ Case Rep. 2012, 1, 2–4. [Google Scholar] [CrossRef] [Green Version]
- Arancibia, F.; Bauer, T.T.; Ewig, S.; Mensa, J.; Gonzalez, J.; Niederman, M.S.; Torres, A. Community-Acquired Pneumonia Due to Gram-Negative Bacteria and Pseudomonas aeruginosa. Am. Med. Assoc. 2014, 162, 1849–1858. [Google Scholar] [CrossRef] [Green Version]
- Priebe, G.P.; Goldberg, J.B. Vaccines for Pseudomonas aeruginosa: A long and winding road. Expert Rev. Vaccines 2014, 13, 507–519. [Google Scholar] [CrossRef] [Green Version]
- Cystic Fibrosis Foundation. Cystic Fibrosis Foundation Patient Registry 2014 Annual Report. Cyst. Fibros. Found. 2014, 25. [Google Scholar]
- Cystic Fibrosis Foundation. Cystic Fibrosis Patient Registry 2018 Annual Data Report; Bethesda: Rockville, MD, USA, 2018. [Google Scholar]
- David, S.C.; Lau, J.; Singleton, E.V.; Babb, R.; Davies, J.; Hirst, T.R.; McColl, S.R.; Paton, J.C.; Alsharifi, M. The effect of gamma-irradiation conditions on the immunogenicity of whole-inactivated Influenza A virus vaccine. Vaccine 2017, 35, 1071–1079. [Google Scholar] [CrossRef] [PubMed]
- Jwa, M.Y.; Jeong, S.; Ko, E.B.; Kim, A.R.; Kim, H.Y.; Kim, S.K.; Seo, H.S.; Yun, C.-H.; Han, S.H. Gamma-irradiation of Streptococcus pneumoniae for the use as an immunogenic whole cell vaccine. J. Microbiol. 2018, 56, 579–585. [Google Scholar] [CrossRef] [PubMed]
- Shahrudin, S.; Chen, C.; David, S.C.; Singleton, E.V.; Davies, J.; Kirkwood, C.D.; Hirst, T.R.; Beard, M.; Alsharifi, M. Gamma-irradiated rotavirus: A possible whole virus inactivated vaccine. PLoS ONE 2018, 13, e0198182. [Google Scholar] [CrossRef] [Green Version]
- Baicus, A. History of polio vaccination. World J. Virol. 2012, 1, 108. [Google Scholar] [CrossRef]
- Milligan, R.; Paul, M.; Richardson, M.; Neuberger, A. Vaccines for preventing typhoid fever. Cochrane Database Syst. Rev. 2018, 5, CD001261. [Google Scholar] [CrossRef] [PubMed]
- Higgs, R.; Higgins, S.C.; Ross, P.J.; Mills, K.H.G. Immunity to the respiratory pathogen Bordetella pertussis. Mucosal Immunol. 2012, 5, 485–500. [Google Scholar] [CrossRef] [Green Version]
- Sato, Y.; Sato, H. Development of acellular pertussis vaccines. Biologicals 1999, 27, 61–69. [Google Scholar] [CrossRef]
- Klein, N.P.; Bartlett, J.; Rowhani-Rahbar, A.; Fireman, B.; Baxter, R. Waning protection after fifth dose of acellular pertussis vaccine in children. N. Engl. J. Med. 2012, 367, 1012–1019. [Google Scholar] [CrossRef]
- Varney, M.E.; Boehm, D.T.; DeRoos, K.; Nowak, E.S.; Wong, T.Y.; Sen-Kilic, E.; Bradford, S.D.; Elkins, C.; Epperly, M.S.; Witt, W.T.; et al. Bordetella pertussis whole cell immunization, unlike acellular immunization, mimics naïve infection by driving hematopoietic stem and progenitor cell expansion in mice. Front. Immunol. 2018, 9, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Smallridge, W.E.; Rolin, O.Y.; Jacobs, N.T.; Harvill, E.T. Different effects of whole-cell and acellular vaccines on Bordetella transmission. J. Infect. Dis. 2014, 209, 1981–1988. [Google Scholar] [CrossRef]
- Allen, A.C.; Wilk, M.M.; Misiak, A.; Borkner, L.; Murphy, D.; Mills, K.H.G. Sustained protective immunity against Bordetella pertussis nasal colonization by intranasal immunization with a vaccine-adjuvant combination that induces IL-17-secreting T RM cells. Mucosal Immunol. 2018, 11, 1763–1776. [Google Scholar] [CrossRef] [Green Version]
- Cauchi, S.; Locht, C. Non-specific Effects of Live Attenuated Pertussis Vaccine Against Heterologous Infectious and Inflammatory Diseases. Front. Immunol. 2018, 9, 2872. [Google Scholar] [PubMed] [Green Version]
- Riese, P.; Sakthivel, P.; Trittel, S.; Guzmán, C.A. Intranasal formulations: Promising strategy to deliver vaccines. Expert Opin. Drug Deliv. 2014, 11, 1619–1634. [Google Scholar]
- Del Giudice, G.; Pizza, M.; Rappuoli, R. Mucosal Delivery of Vaccines. Methods 1999, 19, 148–155. [Google Scholar]
- Wu, W.; Huang, J.; Duan, B.; Traficante, D.C.; Hong, H.; Risech, M.; Lory, S.; Priebe, G.P. Th17-stimulating protein vaccines confer protection against Pseudomonas aeruginosa pneumonia. Am. J. Respir. Crit. Care Med. 2012, 186, 420–427. [Google Scholar] [PubMed] [Green Version]
- Belyakov, I.M.; Ahlers, J.D. What Role Does the Route of Immunization Play in the Generation of Protective Immunity against Mucosal Pathogens? J. Immunol. 2009, 183, 6883–6892. [Google Scholar]
- Boehm, D.T.; Wolf, M.A.; Hall, J.M.; Wong, T.Y.; Sen-Kilic, E.; Basinger, H.D.; Dziadowicz, S.A.; de la Pazc Gutierrez, M.; Blackwood, C.B.; Bradford, S.D.; et al. Intranasal acellular pertussis vaccine provides mucosal immunity and protects mice from Bordetella pertussis. NPJ Vaccines 2019, 4, 1–12. [Google Scholar]
- Warfel, J.M.; Merkel, T.J. Bordetella pertussis infection induces a mucosal IL-17 response and long-lived Th17 and Th1 immune memory cells in nonhuman primates. Mucosal Immunol. 2013, 6, 787–796. [Google Scholar]
- Zhang, R.; Edgar, K.J. Properties, chemistry, and applications of the bioactive polysaccharide curdlan. Biomacromolecules 2014, 15, 1079–1096. [Google Scholar]
- Higashi, T.; Hashimoto, K.; Takagi, R.; Mizuno, Y.; Okazaki, Y.; Tanaka, Y.; Matsushita, S. Curdlan induces DC-mediated Th17 polarization via Jaggedl activation in human dendritic cells. Allergol. Int. 2010, 59, 161–166. [Google Scholar]
- Bordet, J.; Gengou, O. Le Microbe de la Coqueluche. Ann. l’Institut. Pasteur. 1906, 20, 731–741. [Google Scholar]
- Stainer, D.W.; Scholte, M.J. A simple chemically defined medium for the production of phase I Bordetella pertussis. J. Gen. Microbiol. 1970, 63, 211–220. [Google Scholar] [CrossRef] [Green Version]
- Warn, P.A.; Brampton, M.W.; Sharp, A.; Morrissey, G.; Steel, N.; Denning, D.W.; Priest, T. Infrared body temperature measurement of mice as an early predictor of death in experimental fungal infections. Lab. Anim. 2003, 37, 126–131. [Google Scholar] [CrossRef]
- Mei, J.; Riedel, N.; Grittner, U.; Endres, M.; Banneke, S.; Emmrich, J.V. Body temperature measurement in mice during acute illness: Implantable temperature transponder versus surface infrared thermometry. Sci. Rep. 2018, 8, 1–10. [Google Scholar] [CrossRef]
- Ryu, J.I.; Wui, S.R.; Ko, A.; Do, H.T.T.; Lee, Y.J.; Kim, H.J.; Rhee, I.; Park, S.A.; Kim, K.S.; Cho, Y.J.; et al. Increased immunogenicity and protective efficacy of a P. aeruginosa vaccine in mice using an alum and de-O-acylated lipooligosaccharide adjuvant system. J. Microbiol. Biotechnol. 2017, 27, 1539–1548. [Google Scholar]
- Ryu, J.I.; Park, S.A.; Wui, S.R.; Ko, A.; Han, J.E.; Choi, J.A.; Song, M.K.; Kim, K.S.; Cho, Y.J.; Lee, N.G. A De-O-acylated Lipooligosaccharide-Based Adjuvant System Promotes Antibody and Th1-Type Immune Responses to H1N1 Pandemic Influenza Vaccine in Mice. Biomed. Res. Int. 2016, 2016. [Google Scholar] [CrossRef] [Green Version]
- Sen-Kilic, E.; Blackwood, C.B.; Boehm, D.T.; Witt, W.T.; Malkowski, A.C.; Bevere, J.R.; Wong, T.Y.; Hall, J.M.; Bradford, S.D.; Varney, M.E.; et al. Intranasal peptide-based FpvA-KLH conjugate vaccine protects mice from Pseudomonas aeruginosa acute murine pneumonia. Front. Immunol. 2019, 10, 1–17. [Google Scholar] [CrossRef]
- Babicki, S.; Arndt, D.; Marcu, A.; Liang, Y.; Grant, J.R.; Maciejewski, A.; Wishart, D.S. Heatmapper: Web-enabled heat mapping for all. Nucleic. Acids. Res. 2016, 44, W147–W153. [Google Scholar] [CrossRef]
- Metsalu, T.; Vilo, J. ClustVis: A web tool for visualizing clustering of multivariate data using Principal Component Analysis and heatmap. Nucleic. Acids. Res. 2015, 43, W566–W570. [Google Scholar] [CrossRef]
- Kipnis, E.; Sawa, T.; Wiener-Kronish, J. Targeting mechanisms of Pseudomonas aeruginosa pathogenesis. Med. Mal. Infect. 2006, 36, 78–91. [Google Scholar] [CrossRef]
- Mak, T.W.; Saunders, M.E.; Jett, B.D. Primer to The Immune Response, 2nd ed.; Elsevier: Amsterdam, The Netherlands, 2014; ISBN 9780123852458. [Google Scholar]
- Scanlon, K.M.; Snyder, Y.G.; Skerry, C.; Carbonetti, N.H. Fatal pertussis in the neonatal mouse model is associated with pertussis toxin-mediated pathology beyond the airways. Infect. Immun. 2017, 85. [Google Scholar] [CrossRef] [Green Version]
- Pierce, C.; Klein, N.; Peters, M. Is leukocytosis a predictor of mortality in severe pertussis infection? Intensive Care Med. 2000, 26, 1512–1514. [Google Scholar] [CrossRef]
- Locht, C.; Coutte, L.; Mielcarek, N. The ins and outs of pertussis toxin. FEBS J. 2011, 278, 4668–4682. [Google Scholar] [CrossRef]
- Raeven, R.H.; Brummelman, J.; Pennings, J.L.; Nijst, O.E.; Kuipers, B.; Blok, L.E.; Helm, K.; van Riet, E.; Jiskoot, W.; van Els, C.A.; et al. Molecular signatures of the evolving immune response in mice following a Bordetella pertussis infection. PLoS ONE 2014, 9, e104548. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ross, P.J.; Sutton, C.E.; Higgins, S.; Allen, A.C.; Walsh, K.; Misiak, A.; Lavelle, E.C.; McLoughlin, R.M.; Mills, K.H.G. Relative Contribution of Th1 and Th17 Cells in Adaptive Immunity to Bordetella pertussis: Towards the Rational Design of an Improved Acellular Pertussis Vaccine. Infect. Immun. 1998, 66, 594–602. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giese, M. Introduction to molecular vaccinology; Springer: Berlin, Germany, 2016; ISBN 9783319258324. [Google Scholar]
- Center for Disease Control and Prevention (CDC) Pneumococcal Disease. In Epidemiology and Prevention of Vaccine-Preventable Diseases; US Department of Health and Human Services: Washington, DC, USA, 2015; pp. 279–296.
- Kroger, A.; Hamborsky, J. Centers for Disease Control and Prevention, Public Health Foundation. Epidemiology and Prevention of Vaccine-Preventable Diseases; Centers for Disease Control and Prevention: Atlanta, GA, USA, 2015.
- Hervé, C.; Laupèze, B.; Del Giudice, G.; Didierlaurent, A.M.; Da Silva, F.T. The how’s and what’s of vaccine reactogenicity. NPJ Vaccines 2019, 4. [Google Scholar] [CrossRef] [Green Version]
- Lambert, L.C. Pertussis vaccine trials in the 1990s. J. Infect. Dis. 2014, 209 (Suppl. 1), S4–S9. [Google Scholar] [CrossRef] [Green Version]
- Haghighi, F.; Shahcheraghi, F.; Abbasi, E.; Eshraghi, S.S.; Zeraati, H.; Javad Mousavi, S.A.; Asgarian-Omran, H.; Douraghi, M.; Shokri, F. Genetic profile variation in vaccine strains and clinical isolates of Bordetella pertussis recovered from Iranian patients. Avicenna J. Med. Biotechnol. 2014, 6, 178–184. [Google Scholar] [PubMed]
- Weigand, M.R.; Peng, Y.; Loparev, V.; Batra, D.; Bowden, K.E.; Burroughs, M.; Cassiday, P.K.; Davis, J.K.; Johnson, T.; Juieng, P.; et al. The History of Bordetella pertussis Genome Evolution Includes Structural Rearrangement. J. Bacteriol. 2017, 199. [Google Scholar] [CrossRef] [Green Version]
- Wieland, C.W.; Siegmund, B.; Senaldi, G.; Vasil, M.L.; Dinarello, C.A.; Fantuzzi, G. Pulmonary inflammation induced by Pseudomonas aeruginosa lipopolysaccharide, phospholipase C, and exotoxin A: Role of interferon regulatory factor 1. Infect. Immun. 2002, 70, 1352–1358. [Google Scholar] [CrossRef] [Green Version]
- Wahlström, E.; Ollerstam, A.; Sundius, L.; Zhang, H. Use of Lung Weight as Biomarker for Assessment of Lung Toxicity in Rat Inhalation Studies. Toxicol. Pathol. 2013, 41, 902–912. [Google Scholar] [CrossRef] [Green Version]
- Sharplin, J.; Franko, A.J. Pulmonary oedema during the latent period after irradiation of murine lung. Br. J. Cancer 1986, 53, 336–339. [Google Scholar]
- Johansen, H.; Gøtzsche, P. Vaccines for preventing infection with Pseudomonas aeruginosa in cystic fibrosis (Review). Cochrane. Libr. 2015. [Google Scholar]
- Kang, C.; Kim, S.; Kim, H.; Park, S.; Choe, Y.; Oh, M.; Kim, E.; Choe, K. Pseudomonas aeruginosa Bacteremia: Risk Factors for Mortality and Influence of Delayed Receipt of Effective Antimicrobial Therapy on Clinical Outcome. Clin. Infect. Dis. 2003, 37, 745–751. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.S.; Park, B.K.; Koo Kim, S.; Han, S.B.; Lee, J.W.; Lee, D.G.; Chung, N.G.; Cho, B.; Jeong, D.C.; Kang, J.H. Clinical characteristics and outcomes of Pseudomonas aeruginosa bacteremia in febrile neutropenic children and adolescents with the impact of antibiotic resistance: A retrospective study. BMC Infect. Dis. 2017, 17, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Donowitz, G.R.; Maki, D.G.; Crnich, C.J.; Pappas, P.G.; Rolston, K.V. Infections in the neutropenic patient--new views of an old problem. Hematol. Am. Soc. Hematol. Educ. Program. 2001, 113–139. [Google Scholar] [CrossRef] [Green Version]
- Koh, A.Y.; Priebe, G.P.; Pier, G.B. Virulence of Pseudomonas aeruginosa in a murine model of gastrointestinal colonization and dissemination in neutropenia. Infect. Immun. 2005, 73, 2262–2272. [Google Scholar] [CrossRef] [Green Version]
- Koh, A.Y.; Priebe, G.P.; Ray, C.; Van Rooijen, N.; Pier, G.B. Inescapable need for neutrophils as mediators of cellular innate immunity to acute Pseudomonas aeruginosa pneumonia. Infect. Immun. 2009, 77, 5300–5310. [Google Scholar] [CrossRef] [Green Version]
- Chahin, A.; Opal, S.M.; Zorzopulos, J.; Jobes, D.V.; Migdady, Y.; Yamamoto, M.; Parejo, N.; Palardy, J.E.; Horn, D.L. The Novel Immunotherapeutic Oligodeoxynucleotide IMT504 Protects Neutropenic Animals from Fatal Pseudomonas aeruginosa Bacteremia and Sepsis. Antimicrob. Agents Chemother. 2015, 59, 1225–1229. [Google Scholar] [CrossRef] [Green Version]
- Rada, B. Interactions between Neutrophils and Pseudomonas aeruginosa in Cystic Fibrosis. Pathogens 2017, 6, 10. [Google Scholar] [CrossRef]
- Becker, K.A.; Li, X.; Seitz, A.; Steinmann, J.; Koch, A.; Schuchman, E.; Kamler, M.; Edwards, M.J.; Caldwell, C.C.; Gulbins, E. Neutrophils Kill Reactive Oxygen Species-Resistant Pseudomonas aeruginosa by Sphingosine. Cell. Physiol. Biochem. 2017, 43, 1603–1616. [Google Scholar] [CrossRef]
- Parks, Q.M.; Young, R.L.; Poch, K.R.; Malcolm, K.C.; Vasil, M.L.; Nick, J.A. Neutrophil enhancement of Pseudomonas aeruginosa biofilm development: Human F-actin and DNA as targets for therapy. J. Med. Microbiol. 2009, 58, 492–502. [Google Scholar] [CrossRef]
- Carbonetti, N.H. Pertussis leukocytosis: Mechanisms, clinical relevance and treatment. Pathog. Dis. 2016, 74, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Donoso, A.; León, J.; Ramírez, M.; Rojas, G.; Oberpaur, B. pertussis and fatal pulmonary hypertension: A discouraged entity. Scand. J. Infect. Dis. 2005, 37, 145–148. [Google Scholar] [CrossRef] [PubMed]
- Tisoncik, J.R.; Korth, M.J.; Simmons, C.P.; Farrar, J.; Martin, T.R.; Katze, M.G. Into the Eye of the Cytokine Storm. Microbiol. Mol. Biol. Rev. 2012, 76, 16–32. [Google Scholar] [CrossRef] [Green Version]
- Kurahashi, K.; Kajikawa, O.; Sawa, T.; Ohara, M.; Gropper, M.A.; Frank, D.W.; Martin, T.R.; Wiener-Kronish, J.P. Pathogenesis of septic shock in Pseudomonas aeruginosa pneumonia. J. Clin. Investig. 1999, 104, 743–750. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cole, N.; Krockenberger, M.; Bao, S.; Beagley, K.W.; Husband, A.J.; Willcox, M. Effects of exogenous interleukin-6 during Pseudomonas aeruginosa corneal infection. Infect. Immun. 2001, 69, 4116–4119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cusumano, V.; Tufano, M.A.; Mancuso, G.; Carbone, M.; Rossano, F.; Fera, M.T.; Ciliberti, F.A.; Ruocco, E.; Merendino, R.A.; Teti, G. Porins of Pseudomonas aeruginosa induce release of tumor necrosis factor alpha and interleukin-6 by human leukocytes. Infect. Immun. 1997, 65, 1683–1687. [Google Scholar] [CrossRef] [Green Version]
- Son, D.S.; Parl, A.K.; Rice, V.M.; Khabele, D. Keratinocyte chemoattractant (KC)/human growth-regulated oncogene (GRO) chemokines and pro-inflammatory chemokine networks in mouse and human ovarian epithelial cancer cells. Cancer Biol. Ther. 2007, 6, 1308–1318. [Google Scholar] [CrossRef] [Green Version]
- Zygmunt, B.M.; Rharbaoui, F.; Groebe, L.; Guzman, C.A. Intranasal Immunization Promotes Th17 Immune Responses. J. Immunol. 2009, 183, 6933–6938. [Google Scholar] [CrossRef] [Green Version]
- Christensen, D.; Mortensen, R.; Rosenkrands, I.; Dietrich, J.; Andersen, P. Vaccine-induced Th17 cells are established as resident memory cells in the lung and promote local IgA responses. Mucosal Immunol. 2017, 10, 260–270. [Google Scholar] [CrossRef] [Green Version]
- Hou, W.; Wu, Y.; Sun, S.; Shi, M.; Sun, Y.; Yang, C.; Pei, G.; Gu, Y.; Zhong, C.; Sun, B. Pertussis Toxin Enhances Th1 Responses by Stimulation of Dendritic Cells. J. Immunol. 2003, 170, 1728–1736. [Google Scholar] [CrossRef] [PubMed]
- Byrne, P.; McGuirk, P.; Todryk, S.; Mills, K.H.G. Depletion of NK cells results in disseminating lethal infection with Bordetella pertussis associated with a reduction of antigen-specific Th1 and enhancement of Th2, but not Tr1 cells. Eur. J. Immunol. 2004, 34, 2579–2588. [Google Scholar] [CrossRef]
- Mahon, B.P.; Sheahan, B.J.; Griffin, F.; Murphy, G.; Mills, K.H. Atypical disease after Bordetella pertussis respiratory infection of mice with targeted disruptions of interferon-gamma receptor or immunoglobulin mu chain genes. J. Exp. Med. 1997, 186, 1843–1851. [Google Scholar] [CrossRef]
- Mills, K.H.; Barnard, A.; Watkins, J.; Redhead, K. Cell-mediated immunity to Bordetella pertussis: Role of Th1 cells in bacterial clearance in a murine respiratory infection model. Infect. Immun. 1993, 61, 399–410. [Google Scholar] [CrossRef] [Green Version]
- Ryan, M.; Murphy, G.; Gothefors, L.; Nilsson, L.; Storsaeter, J.; Mills, K.H. Bordetella pertussis respiratory infection in children is associated with preferential activation of type 1 T helper cells. J. Infect. Dis. 1997, 175, 1246–1250. [Google Scholar] [CrossRef] [Green Version]
- Mascart, F.; Verscheure, V.; Malfroot, A.; Hainaut, M.; Piérard, D.; Temerman, S.; Peltier, A.; Debrie, A.-S.; Levy, J.; Del Giudice, G.; et al. Bordetella pertussis infection in 2-month-old infants promotes type 1 T cell responses. J. Immunol. 2003, 170, 1504–1509. [Google Scholar] [CrossRef] [Green Version]
- Dunne, A.; Ross, P.J.; Pospisilova, E.; Masin, J.; Meaney, A.; Sutton, C.E.; Iwakura, Y.; Tschopp, J.; Sebo, P.; Mills, K.H. Inflammasome activation by adenylate cyclase toxin directs Th17 responses and protection against Bordetella pertussis. J. Immunol. 2010, 185, 1711–1719. [Google Scholar] [CrossRef] [Green Version]
- Misiak, A.; Wilk, M.M.; Raverdeau, M.; Mills, K.H. IL-17-Producing Innate and Pathogen-Specific Tissue Resident Memory gammadelta T Cells Expand in the Lungs of Bordetella pertussis-Infected Mice. J. Immunol. 2017, 198, 363–374. [Google Scholar] [CrossRef] [Green Version]
- Gregg, K.A.; Harberts, E.; Gardner, F.M.; Pelletier, M.R.; Cayatte, C.; Yu, L.; McCarthy, M.P.; Marshall, J.D.; Ernst, R.K. Rationally designed TLR4 ligands for vaccine adjuvant discovery. MBio 2017, 8, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schülke, S.; Flaczyk, A.; Vogel, L.; Gaudenzio, N.; Angers, I.; Löschner, B.; Wolfheimer, S.; Spreitzer, I.; Qureshi, S.; Tsai, M.; et al. MPLA shows attenuated pro-inflammatory properties and diminished capacity to activate mast cells in comparison with LPS. Allergy. Eur. J. Allergy. Clin. Immunol. 2015, 70, 1259–1268. [Google Scholar] [CrossRef]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Blackwood, C.B.; Sen-Kilic, E.; Boehm, D.T.; Hall, J.M.; Varney, M.E.; Wong, T.Y.; Bradford, S.D.; Bevere, J.R.; Witt, W.T.; Damron, F.H.; et al. Innate and Adaptive Immune Responses against Bordetella pertussis and Pseudomonas aeruginosa in a Murine Model of Mucosal Vaccination against Respiratory Infection. Vaccines 2020, 8, 647. https://doi.org/10.3390/vaccines8040647
Blackwood CB, Sen-Kilic E, Boehm DT, Hall JM, Varney ME, Wong TY, Bradford SD, Bevere JR, Witt WT, Damron FH, et al. Innate and Adaptive Immune Responses against Bordetella pertussis and Pseudomonas aeruginosa in a Murine Model of Mucosal Vaccination against Respiratory Infection. Vaccines. 2020; 8(4):647. https://doi.org/10.3390/vaccines8040647
Chicago/Turabian StyleBlackwood, Catherine B., Emel Sen-Kilic, Dylan T. Boehm, Jesse M. Hall, Melinda E. Varney, Ting Y. Wong, Shelby D. Bradford, Justin R. Bevere, William T. Witt, F. Heath Damron, and et al. 2020. "Innate and Adaptive Immune Responses against Bordetella pertussis and Pseudomonas aeruginosa in a Murine Model of Mucosal Vaccination against Respiratory Infection" Vaccines 8, no. 4: 647. https://doi.org/10.3390/vaccines8040647
APA StyleBlackwood, C. B., Sen-Kilic, E., Boehm, D. T., Hall, J. M., Varney, M. E., Wong, T. Y., Bradford, S. D., Bevere, J. R., Witt, W. T., Damron, F. H., & Barbier, M. (2020). Innate and Adaptive Immune Responses against Bordetella pertussis and Pseudomonas aeruginosa in a Murine Model of Mucosal Vaccination against Respiratory Infection. Vaccines, 8(4), 647. https://doi.org/10.3390/vaccines8040647