Development of a Safe and Highly Efficient Inactivated Vaccine Candidate against Lumpy Skin Disease Virus


Abstract
:1. Introduction
2. Materials and Methods
2.1. General
2.1.1. Animals
2.1.2. Inactivation of Virus Using Binary Ethylenimine
2.1.3. Clinical Score after Challenge Infection
2.1.4. Sample Collection
2.1.5. Molecular Diagnostics
2.1.6. Virus Isolation
2.1.7. Serological Examination
2.2. Proof-of-Concept Study
2.2.1. Vaccine Preparation
2.2.2. Experimental Design and Challenge Infection
2.3. Vaccine-Efficacy Study
2.3.1. Vaccine Preparation
2.3.2. Experimental Design and Challenge Infection
2.3.3. Safety Scoring
3. Results
3.1. Proof-of-Concept-Study
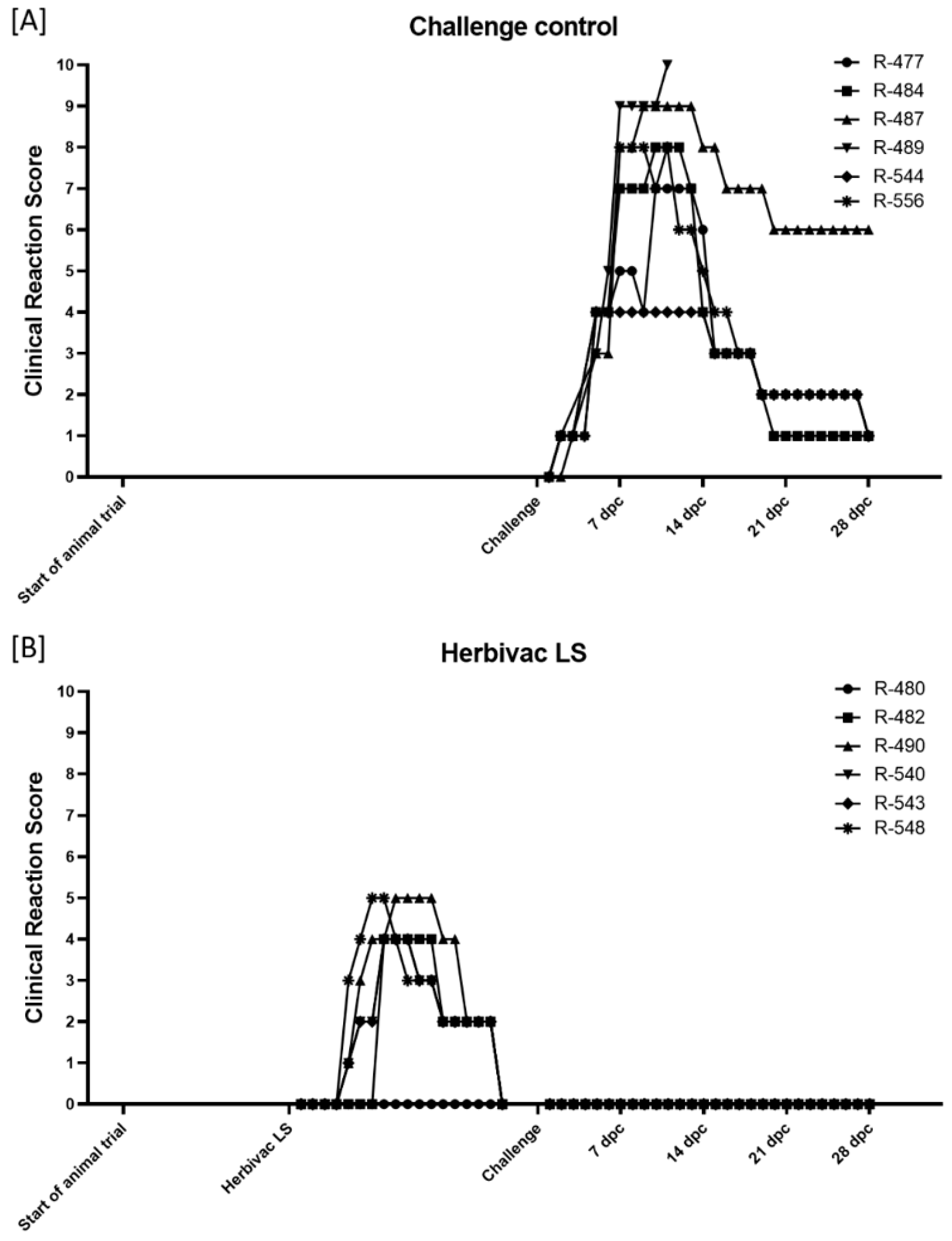
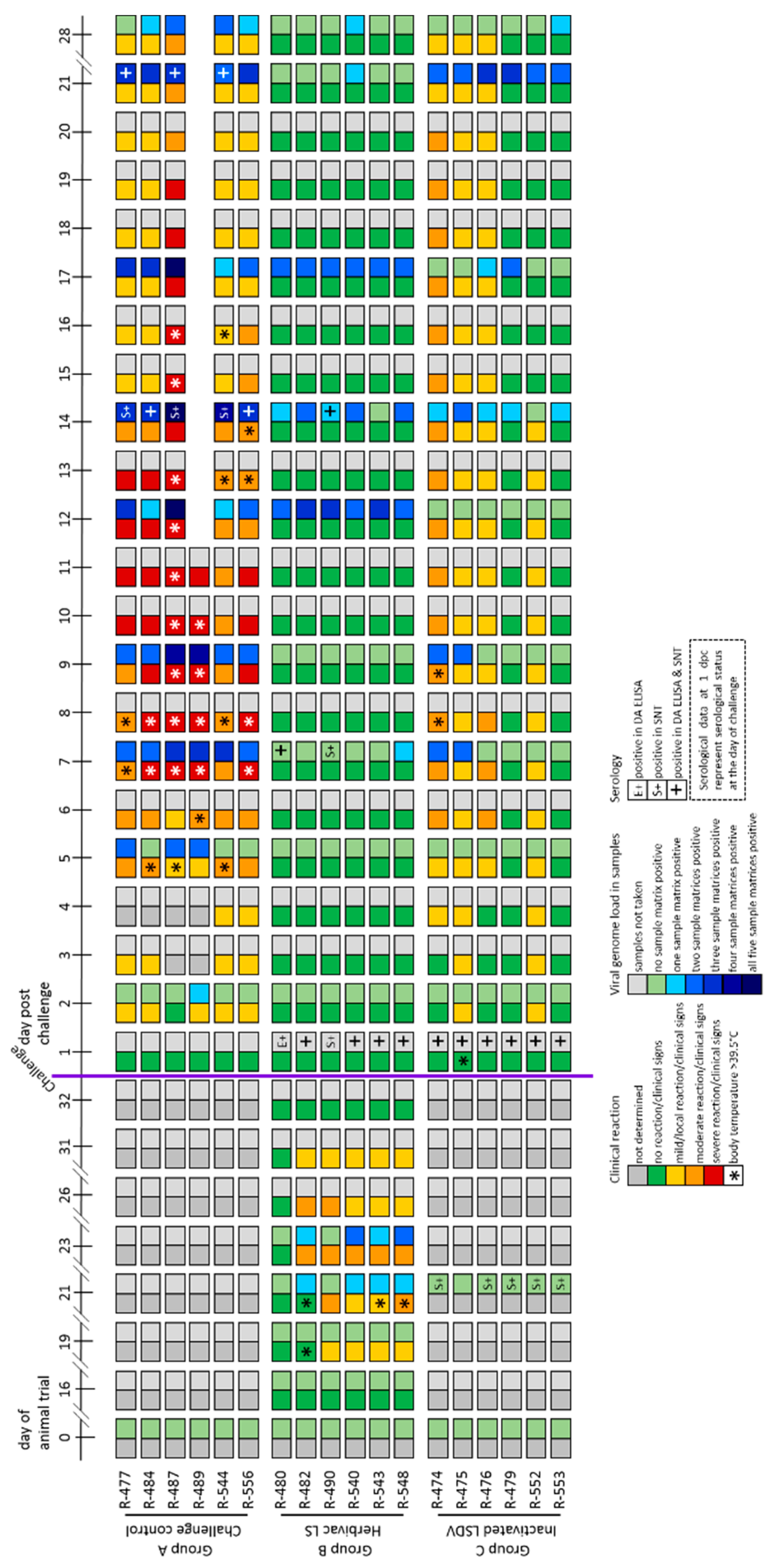
3.1.1. Clinical Reaction
3.1.2. Virus Replication and Shedding
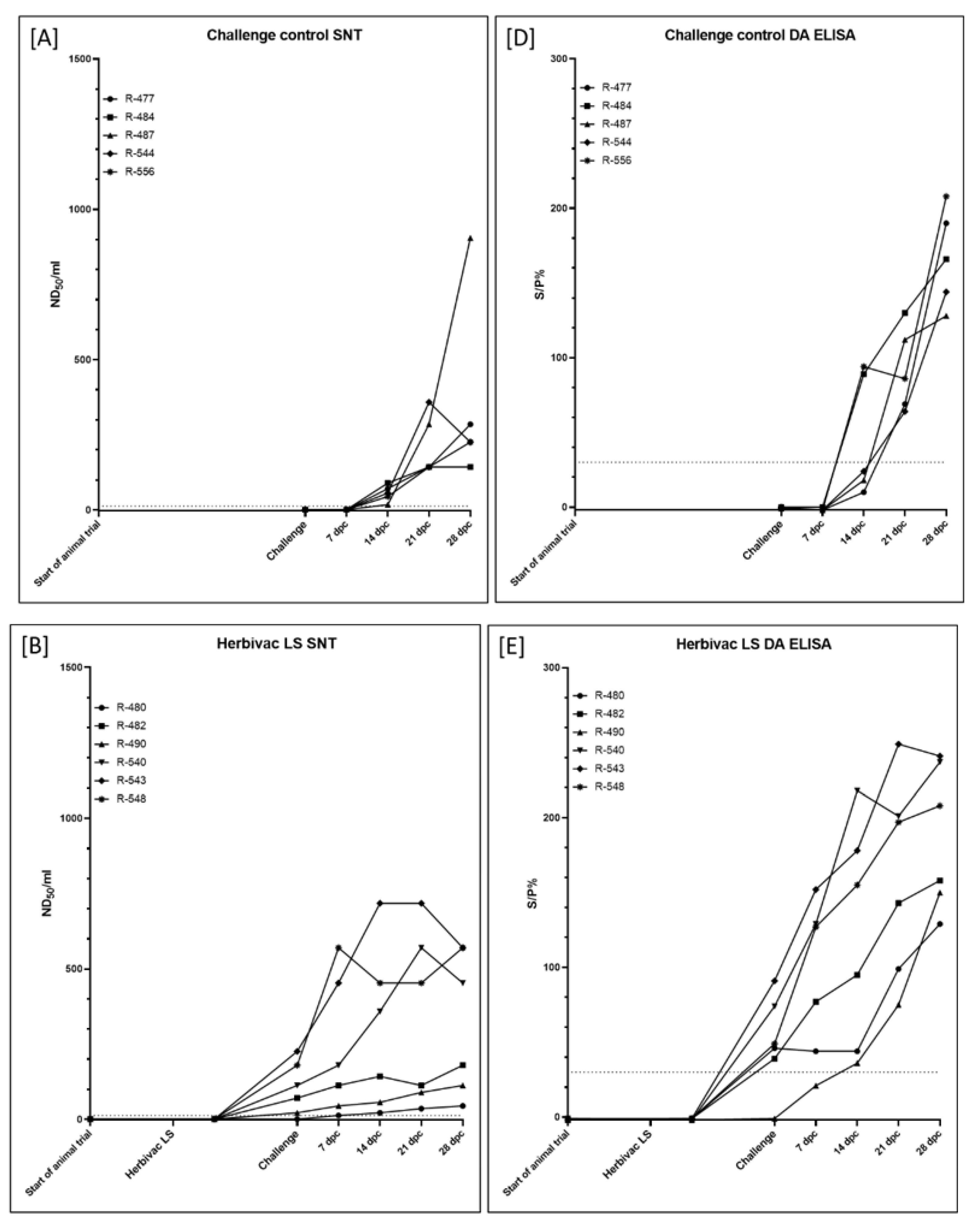
3.1.3. Serological Response
3.2. Vaccine-Efficacy Study
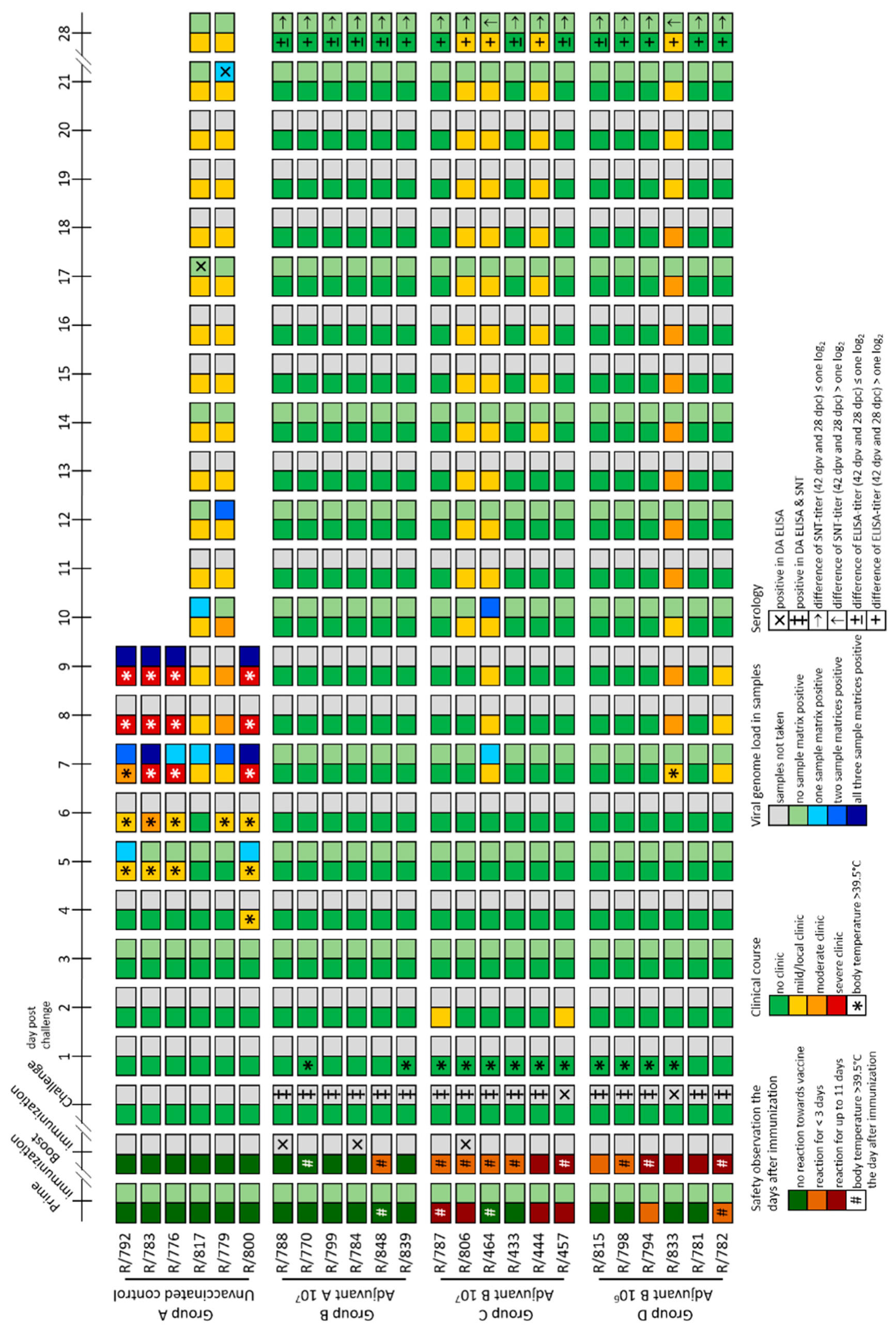
3.2.1. Clinical Reaction and Safety Observation
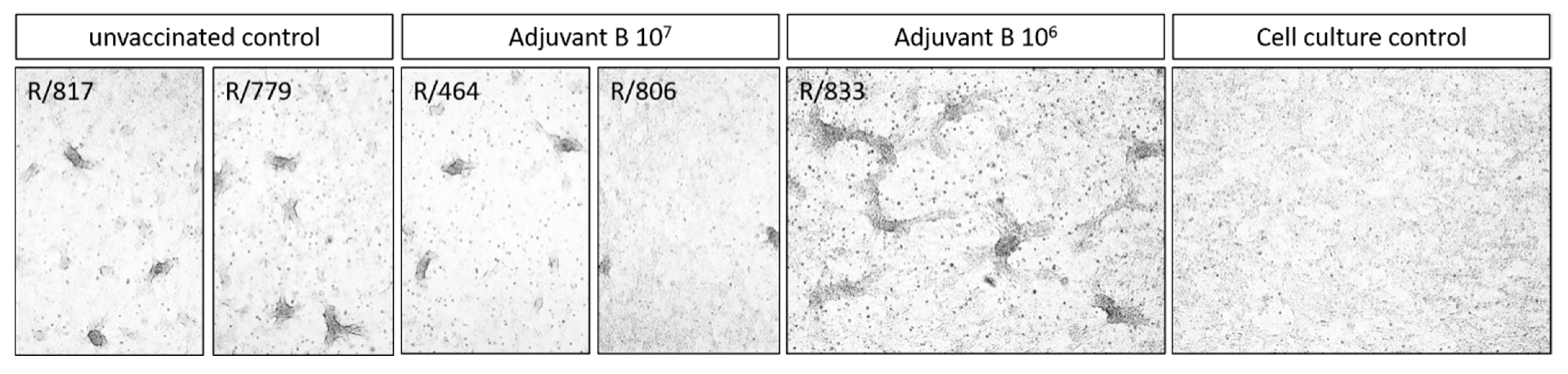
3.2.2. Analyses of Skin Nodes
3.2.3. Virus Replication and Shedding
3.2.4. Viral Genome Load in Certain Organ Samples
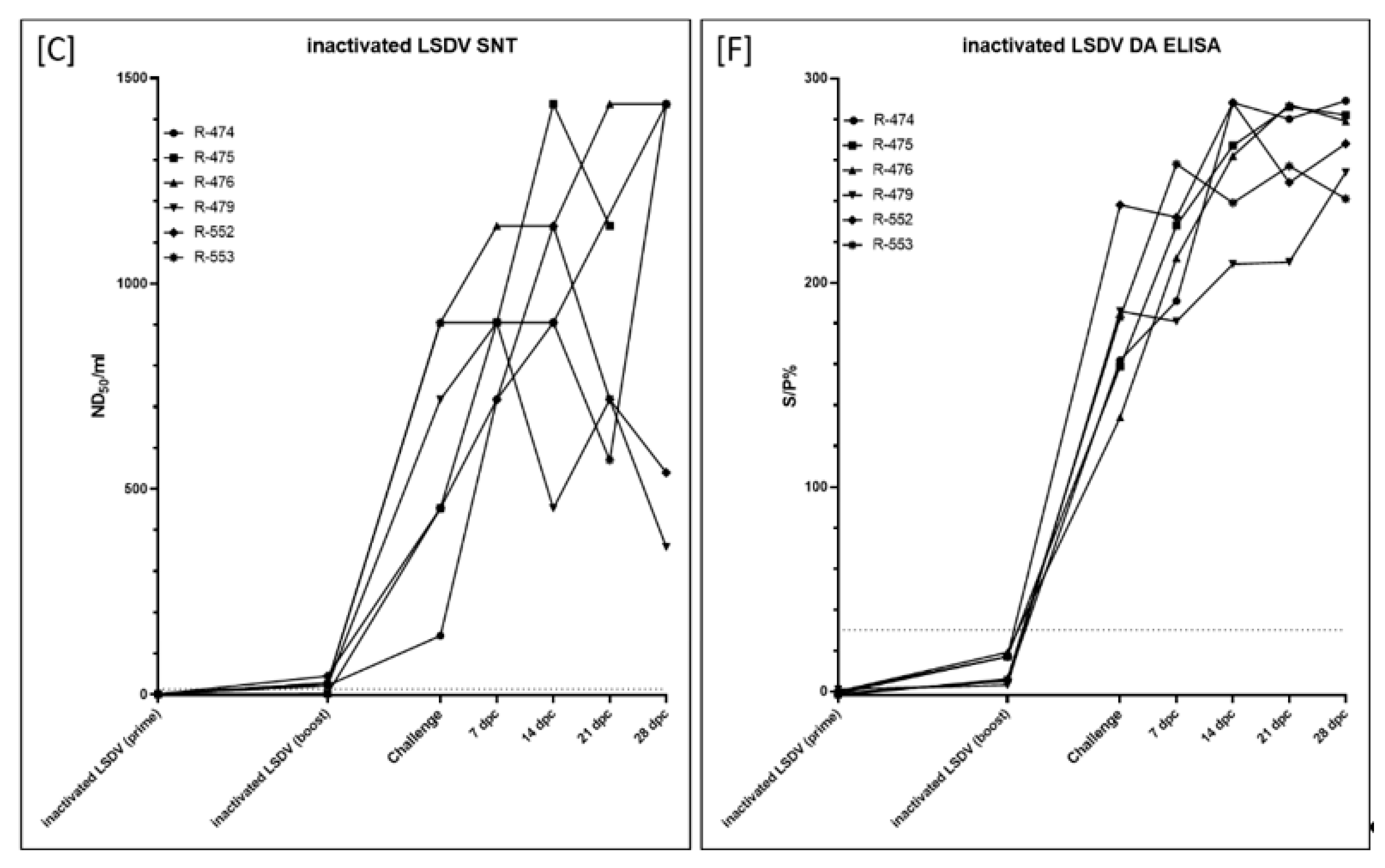
3.2.5. Serological Response
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- King, A.M.Q.; Adams, M.J.; Carstens, E.B.; Lefkowitz, E. Virus Taxonomy-Classification and Nomenclature of Viruses: Ninth Report of the International Committee on Taxonomy of Viruses; Elsevier Academic Press: Amsterdam, The Netherlands, 2012; pp. 291–309. [Google Scholar]
- Carn, V.M. Control of capripoxvirus infections. Vaccine 1993, 11, 1275–1279. [Google Scholar] [CrossRef]
- Babiuk, S.; Bowden, T.R.; Boyle, D.B.; Wallace, D.B.; Kitching, R.P. Capripoxviruses: An emerging worldwide threat to sheep, goats and cattle. Transbound. Emerg. Dis. 2008, 55, 263–272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carn, V.M.; Kitching, R.P. An investigation of possible routes of transmission of lumpy skin disease virus (Neethling). Epidemiol. Infect. 1995, 114, 219–226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mellor, P.S.; Kitching, R.P.; Wilkinson, P.J. Mechanical transmission of capripox virus and African swine fever virus by Stomoxys calcitrans. Res. Vet. Sci. 1987, 43, 109–112. [Google Scholar] [CrossRef]
- Chihota, C.M.; Rennie, L.F.; Kitching, R.P.; Mellor, P.S. Mechanical transmission of lumpy skin disease virus by Aedes aegypti (Diptera: Culicidae). Epidemiol. Infect. 2001, 126, 317–321. [Google Scholar] [CrossRef] [Green Version]
- Chihota, C.M.; Rennie, L.F.; Kitching, R.P.; Mellor, P.S. Attempted mechanical transmission of lumpy skin disease virus by biting insects. Med. Vet. Entomol. 2003, 17, 294–300. [Google Scholar] [CrossRef]
- Tuppurainen, E.S.; Lubinga, J.C.; Stoltsz, W.H.; Troskie, M.; Carpenter, S.T.; Coetzer, J.A.; Venter, E.H.; Oura, C.A. Mechanical transmission of lumpy skin disease virus by Rhipicephalus appendiculatus male ticks. Epidemiol. Infect. 2013, 141, 425–430. [Google Scholar] [CrossRef] [Green Version]
- Tuppurainen, E.S.; Lubinga, J.C.; Stoltsz, W.H.; Troskie, M.; Carpenter, S.T.; Coetzer, J.A.; Venter, E.H.; Oura, C.A. Evidence of vertical transmission of lumpy skin disease virus in Rhipicephalus decoloratus ticks. Ticks Tick Borne Dis. 2013, 4, 329–333. [Google Scholar] [CrossRef] [Green Version]
- Tuppurainen, E.S.; Stoltsz, W.H.; Troskie, M.; Wallace, D.B.; Oura, C.A.; Mellor, P.S.; Coetzer, J.A.; Venter, E.H. A potential role for ixodid (hard) tick vectors in the transmission of lumpy skin disease virus in cattle. Transbound. Emerg. Dis. 2011, 58, 93–104. [Google Scholar] [CrossRef] [Green Version]
- Tuppurainen, E.S.; Venter, E.H.; Coetzer, J.A.; Bell-Sakyi, L. Lumpy skin disease: Attempted propagation in tick cell lines and presence of viral DNA in field ticks collected from naturally-infected cattle. Ticks Tick Borne Dis. 2015, 6, 134–140. [Google Scholar] [CrossRef] [Green Version]
- Haig, D.A. Lumpy skin disease. Bull. Epizoot. Dis. Afr. 1957, 5, 421–430. [Google Scholar]
- Annandale, C.H.; Holm, D.E.; Ebersohn, K.; Venter, E.H. Seminal transmission of lumpy skin disease virus in heifers. Transbound. Emerg. Dis. 2014, 61, 443–448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tuppurainen, E.S.M.; Venter, E.H.; Shisler, J.L.; Gari, G.; Mekonnen, G.A.; Juleff, N.; Lyons, N.A.; De Clercq, K.; Upton, C.; Bowden, T.R.; et al. Review: Capripoxvirus Diseases: Current Status and Opportunities for Control. Transbound. Emerg. Dis. 2017, 64, 729–745. [Google Scholar] [CrossRef] [PubMed]
- Babiuk, S.; Bowden, T.R.; Parkyn, G.; Dalman, B.; Manning, L.; Neufeld, J.; Embury-Hyatt, C.; Copps, J.; Boyle, D.B. Quantification of lumpy skin disease virus following experimental infection in cattle. Transbound. Emerg. Dis. 2008, 55, 299–307. [Google Scholar] [CrossRef] [PubMed]
- Carn, V.M.; Kitching, R.P. The clinical response of cattle experimentally infected with lumpy skin disease (Neethling) virus. Arch. Virol. 1995, 140, 503–513. [Google Scholar] [CrossRef] [PubMed]
- Davies, F.G. Lumpy skin disease, an African capripox virus disease of cattle. Br. Vet. J. 1991, 147, 489–503. [Google Scholar] [CrossRef]
- Spickler, A.R. Lumpy Skin Disease. Available online: http://www.cfsph.iastate.edu/Factsheets/pdfs/lumpy_skin_disease.pdf (accessed on 26 June 2020).
- Sohier, C.; Haegeman, A.; Mostin, L.; De Leeuw, I.; Campe, W.V.; De Vleeschauwer, A.; Tuppurainen, E.S.M.; van den Berg, T.; De Regge, N.; De Clercq, K. Experimental evidence of mechanical lumpy skin disease virus transmission by Stomoxys calcitrans biting flies and Haematopota spp. horseflies. Sci. Rep. 2019, 9, 20076. [Google Scholar] [CrossRef]
- Tuppurainen, E.S.; Oura, C.A. Review: Lumpy skin disease: An emerging threat to Europe, the Middle East and Asia. Transbound. Emerg. Dis. 2012, 59, 40–48. [Google Scholar] [CrossRef]
- Tuppurainen, E.S.; Venter, E.H.; Coetzer, J.A. The detection of lumpy skin disease virus in samples of experimentally infected cattle using different diagnostic techniques. Onderstepoort J. Vet. Res. 2005, 72, 153–164. [Google Scholar] [CrossRef]
- Davies, F.G. Lumpy skin disease of cattle: A growing problem in Africa and the Near East. World Anim Rev. 1991, 68, 37–42. [Google Scholar]
- Weiss, K.E. Lumpy skin disease virus. Virol. Monogr. 1968, 3, 111–131. [Google Scholar]
- Hunter, P.; Wallace, D. Lumpy skin disease in southern Africa: A review of the disease and aspects of control. J. S. Afr Vet. Assoc. 2001, 72, 68–71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- OIE. Manual of Diagnostic Tests and Vaccines for Terrestrial Animals 2019; World Organization for Animal Health, OIE: Paris, France, 2019. [Google Scholar]
- Beard, P.M. Lumpy skin disease: A direct threat to Europe. Vet. Rec 2016, 178, 557–558. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tuppurainen, E.S.; Pearson, C.R.; Bachanek-Bankowska, K.; Knowles, N.J.; Amareen, S.; Frost, L.; Henstock, M.R.; Lamien, C.E.; Diallo, A.; Mertens, P.P. Characterization of sheep pox virus vaccine for cattle against lumpy skin disease virus. Antivir. Res. 2014, 109, 1–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klement, E.; Broglia, A.; Antoniou, S.E.; Tsiamadis, V.; Plevraki, E.; Petrovic, T.; Polacek, V.; Debeljak, Z.; Miteva, A.; Alexandrov, T.; et al. Neethling vaccine proved highly effective in controlling lumpy skin disease epidemics in the Balkans. Prev. Vet. Med. 2018. [Google Scholar] [CrossRef] [PubMed]
- Roth, J.A.; Spickler, A.R. A survey of vaccines produced for OIE list A diseases in OIE member countries. Dev. Biol. 2003, 114, 5–25. [Google Scholar]
- Kitching, R.P. Vaccines for lumpy skin disease, sheep pox and goat pox. Dev. Biol. 2003, 114, 161–167. [Google Scholar]
- Davies, F.G.; Mbugwa, G. The alterations in pathogenicity and immunogenicity of a Kenya sheep and goat pox virus on serial passage in bovine foetal muscle cell cultures. J. Comp. Pathol. 1985, 95, 565–572. [Google Scholar] [CrossRef]
- van Rooyen, P.J.; Munz, E.K.; Weiss, K.E. The optimal conditions for the multiplication of Neethling-type lumpy skin disease virus in embryonated eggs. Onderstepoort J. Vet. Res. 1969, 36, 165–174. [Google Scholar]
- Abutarbush, S.M.; Hananeh, W.M.; Ramadan, W.; Al Sheyab, O.M.; Alnajjar, A.R.; Al Zoubi, I.G.; Knowles, N.J.; Bachanek-Bankowska, K.; Tuppurainen, E.S. Adverse Reactions to Field Vaccination Against Lumpy Skin Disease in Jordan. Transbound. Emerg. Dis. 2016, 63, e213–e219. [Google Scholar] [CrossRef]
- Norian, R.; Afzal Ahangaran, N.; Vashovi, H.R.; Azadmehr, A. Evaluation of Humoral and Cell-mediated Immunity of Two Capripoxvirus Vaccine Strains against Lumpy Skin Disease Virus. Iran. J. Virol. 2016, 10, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Varshovi, H.R.; Norian, R.; Azadmehr, A.; Ahangaran, N.A. Immune response characteristic of Capri pox virus vaccines following emergency vaccination of cattle against lumpy skin disease virus. Iran. J. Vet. Sci Tech. 2018, 9, 33–40. [Google Scholar] [CrossRef]
- Ben-Gera, J.; Klement, E.; Khinich, E.; Stram, Y.; Shpigel, N.Y. Comparison of the efficacy of Neethling lumpy skin disease virus and x10RM65 sheep-pox live attenuated vaccines for the prevention of lumpy skin disease–The results of a randomized controlled field study. Vaccine 2015, 33, 4837–4842. [Google Scholar] [CrossRef] [PubMed]
- Gari, G.; Abie, G.; Gizaw, D.; Wubete, A.; Kidane, M.; Asgedom, H.; Bayissa, B.; Ayelet, G.; Oura, C.A.; Roger, F.; et al. Evaluation of the safety, immunogenicity and efficacy of three capripoxvirus vaccine strains against lumpy skin disease virus. Vaccine 2015, 33, 3256–3261. [Google Scholar] [CrossRef]
- Abdelwahab, M.; Khafagy, H.; Moustafa, A.; Saad, M. Evaluation of Humoral and Cell-mediated Immunity of Lumpy Skin Disease Vaccine Prepared from Local strainin calves and Its Related to Maternal Immunity Evaluation of Humoral and Cell-mediated Immunity of Lumpy Skin Disease Vaccine Prepared from Local in calves and Its. J. Am. Sci. 2016. [Google Scholar] [CrossRef]
- Katsoulos, P.D.; Chaintoutis, S.C.; Dovas, C.I.; Polizopoulou, Z.S.; Brellou, G.D.; Agianniotaki, E.I.; Tasioudi, K.E.; Chondrokouki, E.; Papadopoulos, O.; Karatzias, H.; et al. Investigation on the incidence of adverse reactions, viraemia and haematological changes following field immunization of cattle using a live attenuated vaccine against lumpy skin disease. Transbound. Emerg. Dis. 2018, 65, 174–185. [Google Scholar] [CrossRef]
- Bedekovic, T.; Simic, I.; Kresic, N.; Lojkic, I. Detection of lumpy skin disease virus in skin lesions, blood, nasal swabs and milk following preventive vaccination. Transbound. Emerg. Dis. 2018, 65, 491–496. [Google Scholar] [CrossRef]
- Yeruham, I.; Perl, S.; Nyska, A.; Abraham, A.; Davidson, M.; Haymovitch, M.; Zamir, O.; Grinstein, H. Adverse reactions in cattle to a capripox vaccine. Vet. Rec. 1994, 135, 330–332. [Google Scholar] [CrossRef]
- Morgenstern, M.; Klement, E. The Effect of Vaccination with Live Attenuated Neethling Lumpy Skin Disease Vaccine on Milk Production and Mortality-An Analysis of 77 Dairy Farms in Israel. Vaccines 2020, 8, 324. [Google Scholar] [CrossRef]
- Brenner, J.; Bellaiche, M.; Gross, E.; Elad, D.; Oved, Z.; Haimovitz, M.; Wasserman, A.; Friedgut, O.; Stram, Y.; Bumbarov, V.; et al. Appearance of skin lesions in cattle populations vaccinated against lumpy skin disease: Statutory challenge. Vaccine 2009, 27, 1500–1503. [Google Scholar] [CrossRef]
- Kononov, A.; Byadovskaya, O.; Kononova, S.; Yashin, R.; Zinyakov, N.; Mischenko, V.; Perevozchikova, N.; Sprygin, A. Detection of vaccine-like strains of lumpy skin disease virus in outbreaks in Russia in 2017. Arch. Virol. 2019, 164, 1575–1585. [Google Scholar] [CrossRef] [PubMed]
- Boumart, Z.; Daouam, S.; Belkourati, I.; Rafi, L.; Tuppurainen, E.; Tadlaoui, K.O.; El Harrak, M. Comparative innocuity and efficacy of live and inactivated sheeppox vaccines. BMC Vet. Res. 2016, 12, 133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamdi, J.; Boumart, Z.; Daouam, S.; El Arkam, A.; Bamouh, Z.; Jazouli, M.; Tadlaoui, K.O.; Fihri, O.F.; Gavrilov, B.; El Harrak, M. Development and Evaluation of an Inactivated Lumpy Skin Disease Vaccine for Cattle. Vet. Microbiol. 2020, 245, 108689. [Google Scholar] [CrossRef] [PubMed]
- Kitching, P. Progress towards sheep and goat pox vaccines. Vaccine 1983, 1, 4–9. [Google Scholar] [CrossRef]
- Solyom, F.; Perenlei, L.; Roith, J. Sheep-pox vaccine prepared from formaldehyde inactivated virus adsorbed to aluminium hydroxide gel. Acta Microbiol. Acad. Sci. Hung. 1982, 29, 69–75. [Google Scholar]
- Awad, M.; Michael, A.; Soliman, S.M.; Samir, S.S.; Daoud, A.M. Trials for preparation of inactivated sheep pox vaccine using binary ethyleneimine. Egypt. J. Immunol. 2003, 10, 67–72. [Google Scholar]
- Yadav, M.P.; Pandey, A.B.; Negi, B.S.; Sharma, B.; Shankar, H. Studies on inactivated goat pox vaccine. Indian J. Virol. 1986, 2, 207–221. [Google Scholar]
- Kavitha, L.; Chetty, M. Efficacy of inactivated and live attenuated goatpox vaccines. Indian J. Anim. Sci. 2009, 79, 1018–1019. [Google Scholar]
- Pal, J.K.; Soman, J.P. Further trials on the inactivated goat pox vaccine. Indian J. Virol. 1992, 8, 86–91. [Google Scholar]
- Aarthi, D.; Ananda Rao, K.; Robinson, R.; Srinivasan, V.A. Validation of binary ethyleneimine (BEI) used as an inactivant for foot and mouth disease tissue culture vaccine. Biologicals 2004, 32, 153–156. [Google Scholar] [CrossRef]
- Bahnemann, H.G. Inactivation of viruses in serum with binary ethyleneimine. J. Clin. Microbiol. 1976, 3, 209–210. [Google Scholar]
- Wolff, J.; Krstevski, K.; Beer, M.; Hoffmann, B. Minimum Infective Dose of a Lumpy Skin Disease Virus Field Strain from North Macedonia. Viruses 2020, 12, 768. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, B.; Depner, K.; Schirrmeier, H.; Beer, M. A universal heterologous internal control system for duplex real-time RT-PCR assays used in a detection system for pestiviruses. J. Virol. Methods 2006, 136, 200–209. [Google Scholar] [CrossRef] [PubMed]
- Bowden, T.R.; Babiuk, S.L.; Parkyn, G.R.; Copps, J.S.; Boyle, D.B. Capripoxvirus tissue tropism and shedding: A quantitative study in experimentally infected sheep and goats. Virology 2008, 371, 380–393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dietze, K.; Moritz, T.; Alexandrov, T.; Krstevski, K.; Schlottau, K.; Milovanovic, M.; Hoffmann, D.; Hoffmann, B. Suitability of group-level oral fluid sampling in ruminant populations for lumpy skin disease virus detection. Vet. Microbiol. 2018, 221, 44–48. [Google Scholar] [CrossRef]
- Kärber, G. Beitrag zur kollektiven Behandlung pharmakologischer Reihenversuche. Naunyn-Schmiedebergs Archiv. Exp. Pathol. Pharmakol. 1931, 162, 480–483. [Google Scholar] [CrossRef]
- Spearman, C. The method of ‘right and wrong cases’ (‘constant stimuli’) without gauss’s formulae. Br. J. Psychol. 1908, 2, 227–242. [Google Scholar] [CrossRef]
- Zoetis. Herbivac LS. Available online: https://www.zoetis.co.za/products/beef-and-feedlot/herbivac-ls.aspx (accessed on 9 December 2020).
- Möller, J.; Moritz, T.; Schlottau, K.; Krstevski, K.; Hoffmann, D.; Beer, M.; Hoffmann, B. Experimental lumpy skin disease virus infection of cattle: Comparison of a field strain and a vaccine strain. Arch. Virol. 2019, 164, 2931–2941. [Google Scholar] [CrossRef]
- Bastian, M.; Holsteg, M.; Hanke-Robinson, H.; Duchow, K.; Cussler, K. Bovine Neonatal Pancytopenia: Is this alloimmune syndrome caused by vaccine-induced alloreactive antibodies? Vaccine 2011, 29, 5267–5275. [Google Scholar] [CrossRef]
- MVP-Adjuvants. Information about Polygen. Available online: https://mvpadjuvants.com/wp-content/uploads/2017/10/PHIB-17018-Adjuvants-Bulletin_Polygen.pdf (accessed on 26 June 2020).
- Srivastava, R.N.; Singh, I.P. A study on the role of cellular and humoral factors in immunity to sheep-pox. Indian J. Anim. Sci. 1980, 50, 861–866. [Google Scholar]
- Bachh, A.S.; Ram, G.C.; Hopkins, J.; Bansal, M.P. Observations on cellular response in experimentally sheep-pox-infected lambs. Indian J. Anim. Sci. 1997, 67, 263–266. [Google Scholar]















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
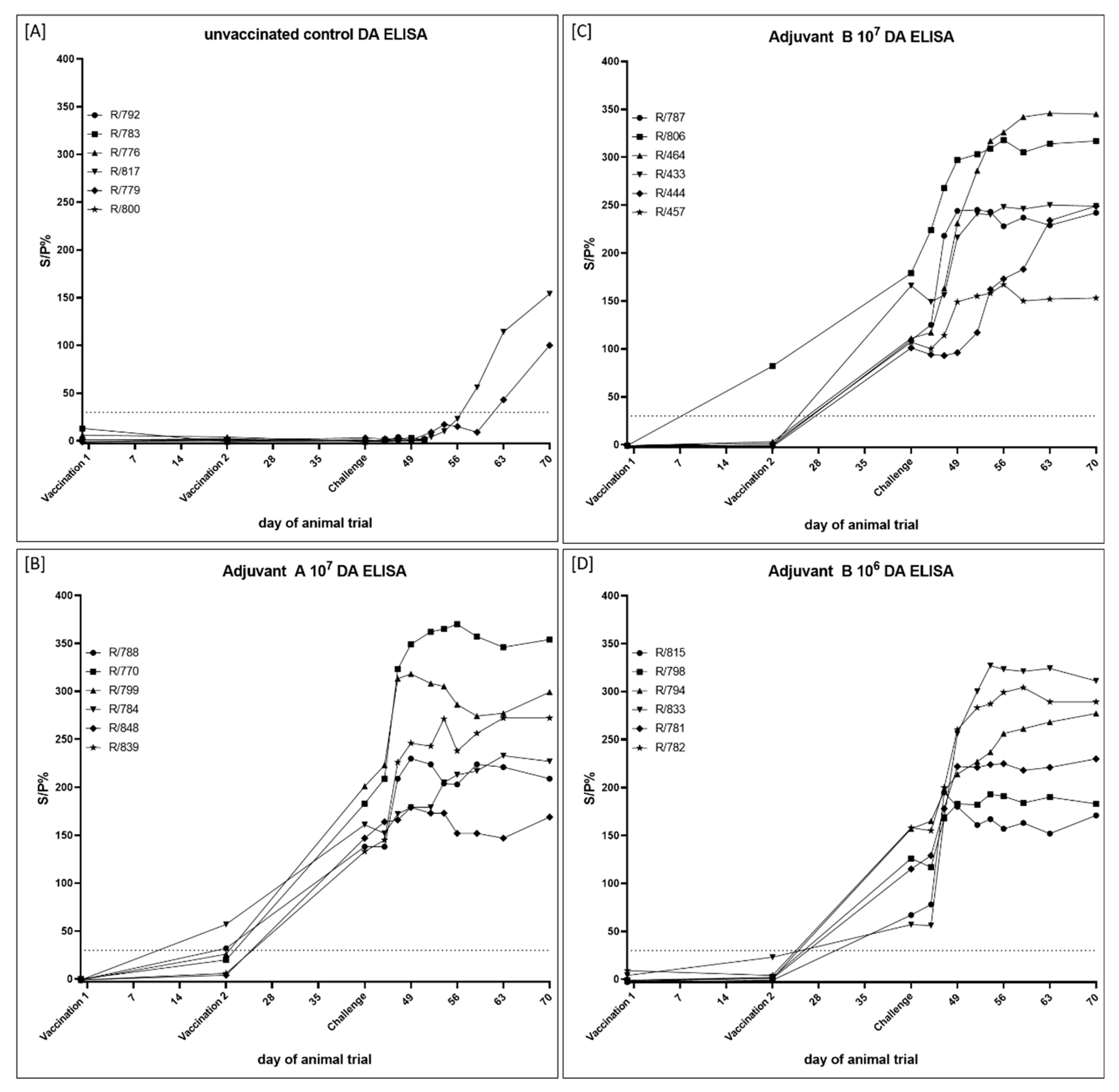{kind=link}
| Component | Adjuvant A 107 (Group 2B) | Adjuvant B 107 (Group 2C) | Adjuvant B 106 (Group 2D) |
|---|---|---|---|
| Inactivated antigen | 10 mL | 10 mL | 1 mL |
| Adjuvant A | 2 mL | - | - |
| Adjuvant B | - | 10 mL | 10 mL |
| TE buffer pH 8.0 | 8 mL | - | - |
| 0.063% PBS | - | - | 9 mL |
| Group | Cattle | Pan-Capripox Real-Time qPCR (Cq) | ||
|---|---|---|---|---|
| 11 dpc | 13 dpc | 17dpc | ||
| Group 2A—unvaccinated control (PBS) | R/779 | 20.2 | n.t. | n.t. |
| 24.6 | n.t. | n.t. | ||
| R/817 | 21.9 | n.t. | n.t. | |
| 20.6 | n.t. | n.t. | ||
| 23.4 | n.t. | n.t. | ||
| Group 2B—Adjuvant A 107 | R/784 | no Cq | n.t. | n.t. |
| Group 2C—Adjuvant B 107 | R/806 | 21.5 | no Cq | n.t. |
| n.t. | 37.3 | n.t. | ||
| R/464 | 18.1 | 17.9 | n.t. | |
| n.t. | 16.4 | n.t. | ||
| R/444 | n.t. | n.t. | 20.9 | |
| n.t. | n.t. | 31.4 | ||
| Group 2D—Adjuvant B 106 | R/833 | 17.8 | 18.1 | n.t. |
| n.t. | 17.2 | n.t. | ||
| Group | Cattle | Cervical Lymph Node | Mediastinal Lymph Node | Mesenterial Lymph Node |
|---|---|---|---|---|
| Group 2A—unvaccinated control (PBS) | R/792 | 21.9 | 27.4 | no Cq |
| R/783 | 27.4 | 23.3 | no Cq | |
| R/776 | 28.4 | 35.2 | no Cq | |
| R/817 | 38.0 | no Cq | no Cq | |
| R/779 | no Cq | no Cq | no Cq | |
| R/800 | 27.6 | 35.2 | no Cq | |
| Group 2B—Adjuvant A 107 | R/788 | no Cq | n.a. | n.a. |
| R/770 | no Cq | n.a. | n.a. | |
| R/799 | no Cq | n.a. | n.a. | |
| R/784 | no Cq | n.a. | n.a. | |
| R/848 | no Cq | n.a. | n.a. | |
| R/839 | no Cq | n.a. | n.a. | |
| Group 2C—Adjuvant B 107 | R/787 | no Cq | n.a. | n.a. |
| R/806 | no Cq | n.a. | n.a. | |
| R/464 | no Cq | n.a. | n.a. | |
| R/433 | no Cq | n.a. | n.a. | |
| R/444 | no Cq | n.a. | n.a. | |
| R/457 | no Cq | n.a. | n.a. | |
| Group 2D—Adjuvant B 106 | R/815 | no Cq | n.a. | n.a. |
| R/798 | no Cq | n.a. | n.a. | |
| R/794 | no Cq | n.a. | n.a. | |
| R/833 | no Cq | n.a. | n.a. | |
| R/781 | no Cq | n.a. | n.a. | |
| R/782 | no Cq | n.a. | n.a. |
| Group | Cattle | DA ELISA | SNT | ||||
|---|---|---|---|---|---|---|---|
| 42 dpv1 | 28 dpc | Comparison | 42 dpv1 | 28 dpc | Comparison | ||
| Group 2B Adjuvant A 107 | R/788 | 1:32 | 1:64 | → | 1:50 | 1:40 | → |
| R/770 | 1:32 | 1:256 | ↑ | 1:40 | 1:16 | → | |
| R/799 | 1:64 | 1:128 | → | 1:80 | 1:80 | → | |
| R/784 | 1:16 | 1:32 | → | 1:40 | 1:50 | → | |
| R/848 | 1:32 | 1:16 | → | 1:20 | 1:13 | → | |
| R/839 | 1:16 | 1:64 | ↑ | 1:32 | 1:20 | → | |
| Group 2C Adjuvant B 107 | R/787 | 1:16 | 1:64 | ↑ | 1:100 | 1:80 | → |
| R/806 | 1:64 | 1:256 | ↑ | 1:200 | 1:256 | → | |
| R/464 | 1:8 | 1:2048 | ↑ | 1:80 | 1:1024 | ↑ | |
| R/433 | 1:32 | 1:64 | → | 1:160 | 1:100 | → | |
| R/444 | 1:8 | 1:64 | ↑ | 1:128 | 1:128 | → | |
| R/457 | 1:8 | 1:16 | → | 1:13 | 1:32 | → | |
| Group 2D Adjuvant B 106 | R/815 | 1:8 | 1:16 | → | 1:32 | 1:20 | → |
| R/798 | 1:16 | 1:64 | ↑ | 1:64 | 1:32 | → | |
| R/794 | 1:32 | 1:128 | ↑ | 1:40 | 1:50 | → | |
| R/833 | 1:2 | 1:1024 | ↑ | 1:13 | 1:400 | ↑ | |
| R/781 | 1:8 | 1:32 | ↑ | 1:100 | 1:32 | → | |
| R/782 | 1:16 | 1:128 | ↑ | 1:50 | 1:160 | → | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wolff, J.; Moritz, T.; Schlottau, K.; Hoffmann, D.; Beer, M.; Hoffmann, B. Development of a Safe and Highly Efficient Inactivated Vaccine Candidate against Lumpy Skin Disease Virus. Vaccines 2021, 9, 4. https://doi.org/10.3390/vaccines9010004
Wolff J, Moritz T, Schlottau K, Hoffmann D, Beer M, Hoffmann B. Development of a Safe and Highly Efficient Inactivated Vaccine Candidate against Lumpy Skin Disease Virus. Vaccines. 2021; 9(1):4. https://doi.org/10.3390/vaccines9010004
Chicago/Turabian StyleWolff, Janika, Tom Moritz, Kore Schlottau, Donata Hoffmann, Martin Beer, and Bernd Hoffmann. 2021. "Development of a Safe and Highly Efficient Inactivated Vaccine Candidate against Lumpy Skin Disease Virus" Vaccines 9, no. 1: 4. https://doi.org/10.3390/vaccines9010004
APA StyleWolff, J., Moritz, T., Schlottau, K., Hoffmann, D., Beer, M., & Hoffmann, B. (2021). Development of a Safe and Highly Efficient Inactivated Vaccine Candidate against Lumpy Skin Disease Virus. Vaccines, 9(1), 4. https://doi.org/10.3390/vaccines9010004