
Recombinant BCG-Prime and DNA-Boost Immunization Confers Mice with Enhanced Protection against Mycobacterium kansasii

 , , and
, , and 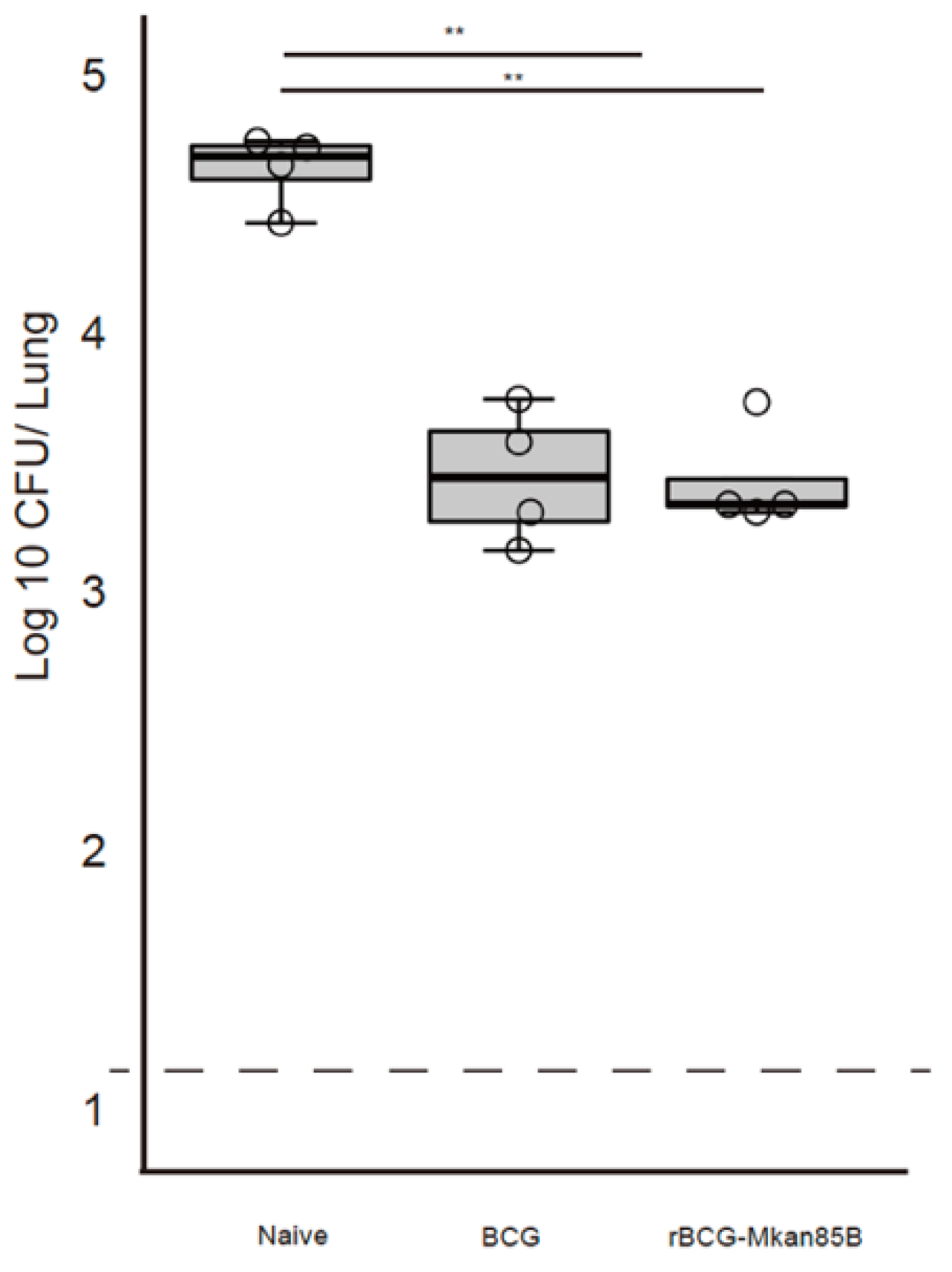{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. BCG and rBCG-Mkan85B Culture
2.3. Immunization
2.4. Bacterial Infection
2.5. Infection Assay
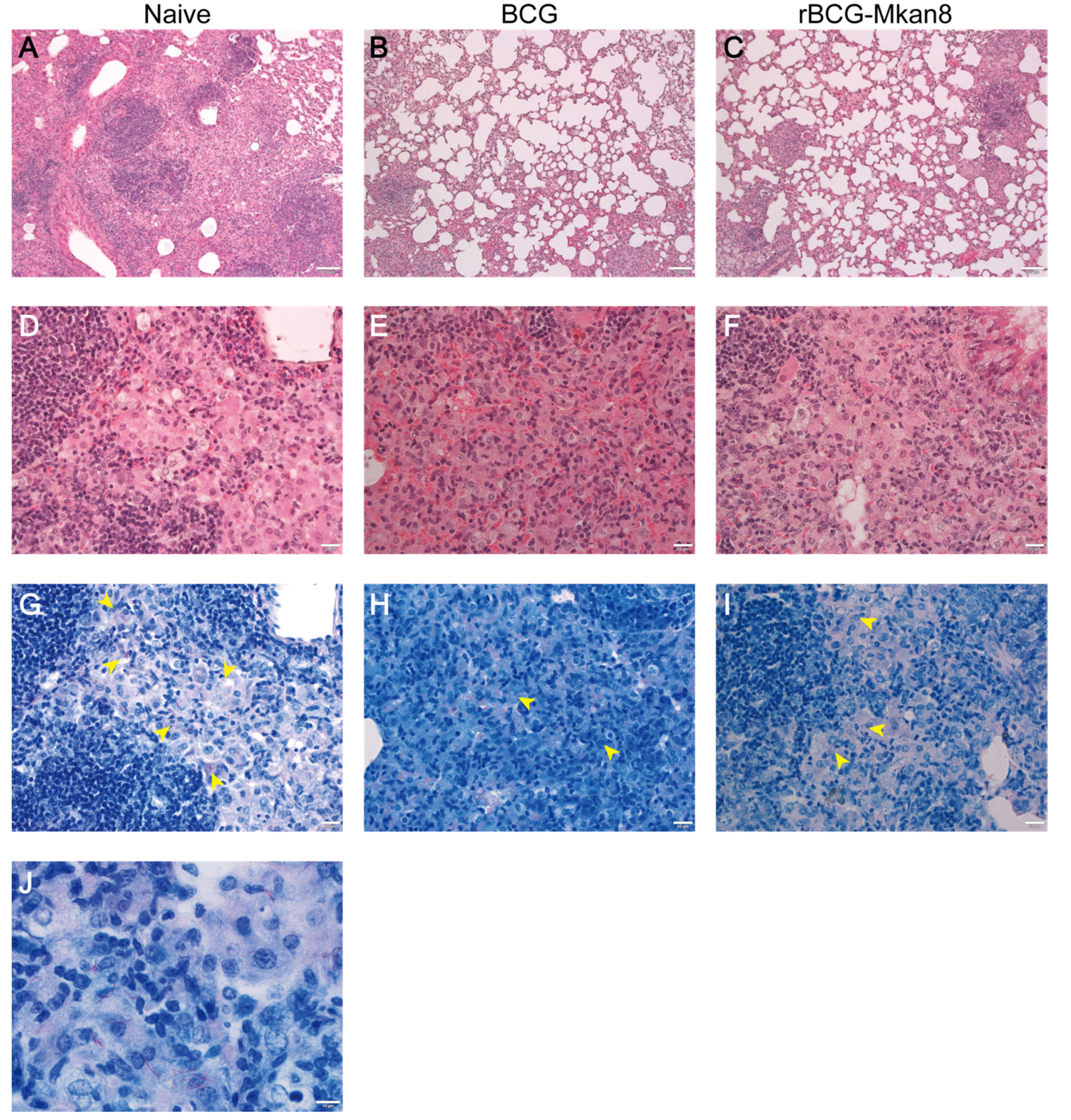
2.6. Histopathology
2.7. Polychromatic Flow Cytometry for Intracellular Cytokine Production
2.8. Data Analysis and Statistics
3. Results
3.1. BCG or rBCG-Mkan85B Vaccination Protects against M. kansasii Infection
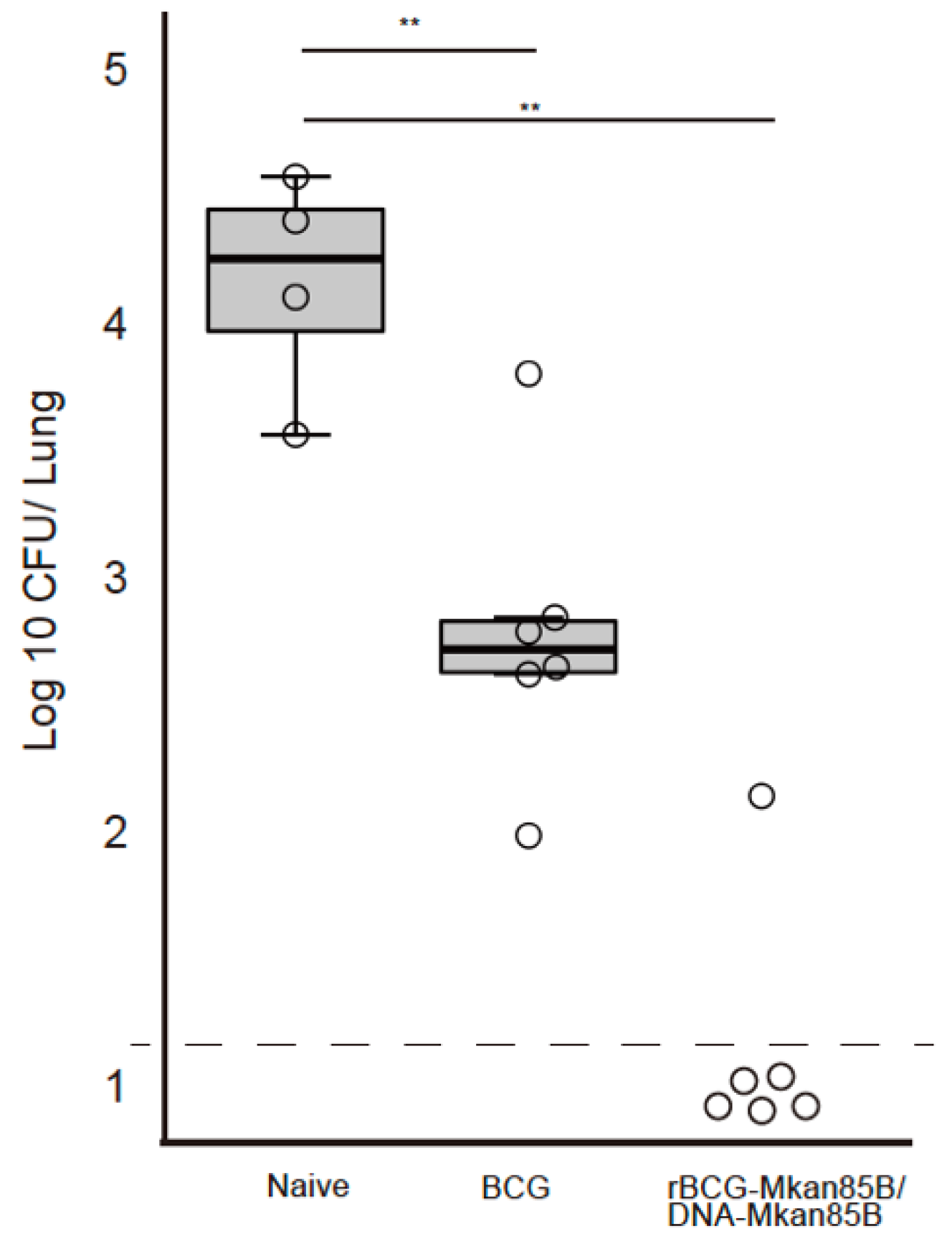
3.2. rBCG-Mkan85B/DNA-Mkan85B Prime–Boost Immunization Elicits Protection against Intratracheal Infection with M. kansasii
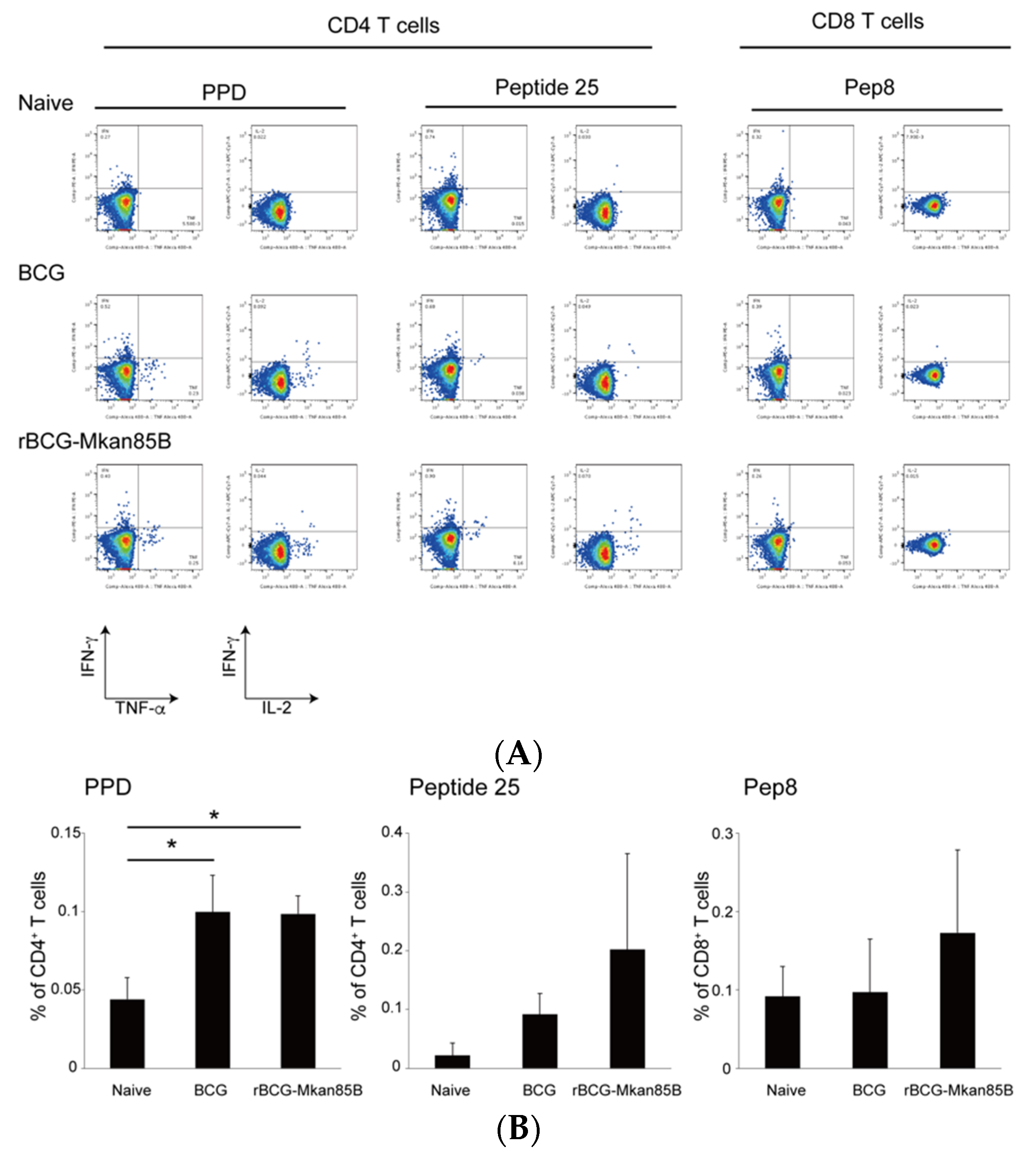
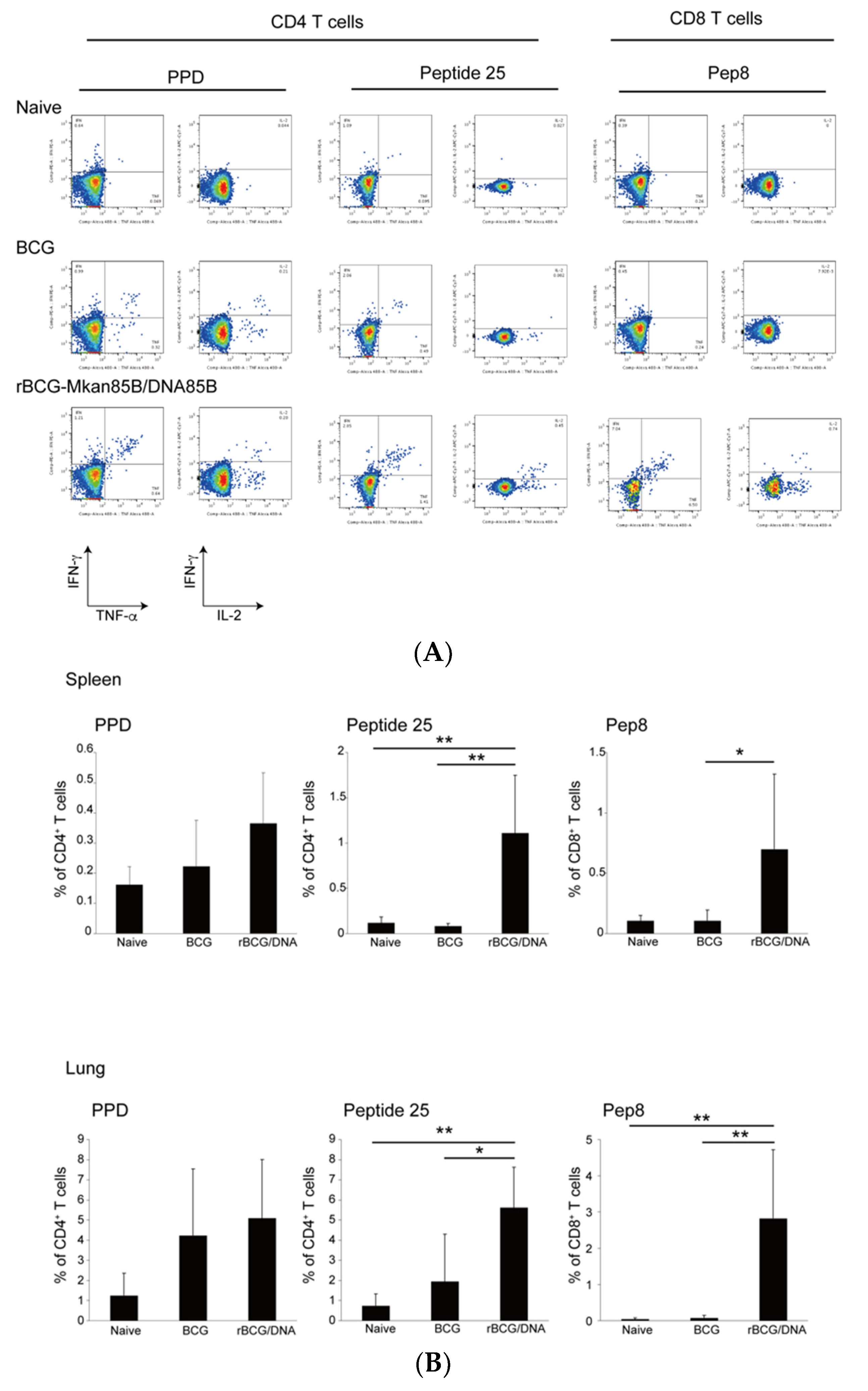
3.3. rBCG-Mkan85B/DNA-Mkan85B Prime–Boost Vaccination Induced Polyfunctional CD8+ T Cells Specific for the Major Secretory Protein Ag85B in the Lung and Spleen
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Namkoong, H.; Kurashima, A.; Morimoto, K.; Hoshino, Y.; Hasegawa, N.; Ato, M.; Mitarai, S. Epidemiology of Pulmonary Nontuberculous Mycobacterial Disease, Japan. Emerg. Infect. Dis. 2016, 22, 1116–1117. [Google Scholar] [CrossRef] [Green Version]
- Prevots, D.R.; Marras, T.K. Epidemiology of human pulmonary infection with nontuberculous mycobacteria: A review. Clin. Chest Med. 2015, 36, 13–34. [Google Scholar] [CrossRef] [Green Version]
- Lee, Y.M.; Kim, M.J.; Kim, Y.J. Increasing Trend of Nontuberculous Mycobacteria Isolation in a Referral Clinical Laboratory in South Korea. Medicina 2021, 57, 720. [Google Scholar] [CrossRef] [PubMed]
- Maekura, R.; Okuda, Y.; Hirotani, A.; Kitada, S.; Hiraga, T.; Yoshimura, K.; Yano, I.; Kobayashi, K.; Ito, M. Clinical and prognostic importance of serotyping Mycobacterium avium-Mycobacterium intracellulare complex isolates in human immunodeficiency virus-negative patients. J. Clin. Microbiol. 2005, 43, 3150–3158. [Google Scholar] [CrossRef] [Green Version]
- de Mello, K.G.; Mello, F.C.; Borga, L.; Rolla, V.; Duarte, R.S.; Sampaio, E.P.; Holland, S.M.; Prevots, D.R.; Dalcolmo, M.P. Clinical and therapeutic features of pulmonary nontuberculous mycobacterial disease, Brazil, 1993–2011. Emerg. Infect. Dis. 2013, 19, 393–399. [Google Scholar] [CrossRef]
- Griffith, D.E.; Aksamit, T.; Brown-Elliott, B.A.; Catanzaro, A.; Daley, C.; Gordin, F.; Holland, S.M.; Horsburgh, R.; Huitt, G.; Iademarco, M.F.; et al. An official ATS/IDSA statement: Diagnosis, treatment, and prevention of nontuberculous mycobacterial diseases. Am. J. Respir Crit. Care Med. 2007, 175, 367–416. [Google Scholar] [CrossRef] [PubMed]
- Silwal, P.; Kim, I.S.; Jo, E.K. Autophagy and Host Defense in Nontuberculous Mycobacterial Infection. Front. Immunol. 2021, 12, 728742. [Google Scholar] [CrossRef] [PubMed]
- Kasperbauer, S.H.; De Groote, M.A. The treatment of rapidly growing mycobacterial infections. Clin. Chest Med. 2015, 36, 67–78. [Google Scholar] [CrossRef]
- Horsburgh, C.R., Jr.; Hanson, D.L.; Jones, J.L.; Thompson, S.E., 3rd. Protection from Mycobacterium avium complex disease in human immunodeficiency virus-infected persons with a history of tuberculosis. J. Infect. Dis. 1996, 174, 1212–1217. [Google Scholar] [CrossRef] [Green Version]
- Brode, S.K.; Daley, C.L.; Marras, T.K. The epidemiologic relationship between tuberculosis and non-tuberculous mycobacterial disease: A systematic review. Int. J. Tuberc. Lung Dis. 2014, 18, 1370–1377. [Google Scholar] [CrossRef]
- Abate, G.; Hamzabegovic, F.; Eickhoff, C.S.; Hoft, D.F. BCG Vaccination Induces M. avium and M. abscessus Cross-Protective Immunity. Front. Immunol. 2019, 10, 234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zimmermann, P.; Finn, A.; Curtis, N. Does BCG Vaccination Protect Against Nontuberculous Mycobacterial Infection? A Systematic Review and Meta-Analysis. J. Infect. Dis. 2018, 218, 679–687. [Google Scholar] [CrossRef] [PubMed]
- Komine-Aizawa, S.; Jiang, J.; Mizuno, S.; Hayakawa, S.; Matsuo, K.; Boyd, L.F.; Margulies, D.H.; Honda, M. MHC-restricted Ag85B-specific CD8(+) T cells are enhanced by recombinant BCG prime and DNA boost immunization in mice. Eur. J. Immunol. 2019, 49, 1399–1414. [Google Scholar] [CrossRef]
- Wiker, H.G.; Harboe, M. The antigen 85 complex: A major secretion product of Mycobacterium tuberculosis. Microbiol. Rev. 1992, 56, 648–661. [Google Scholar] [CrossRef]
- Launois, P.; Drowart, A.; Bourreau, E.; Couppie, P.; Farber, C.M.; Van Vooren, J.P.; Huygen, K. T cell reactivity against mycolyl transferase antigen 85 of M. tuberculosis in HIV-TB coinfected subjects and in AIDS patients suffering from tuberculosis and nontuberculous mycobacterial infections. Clin. Dev. Immunol. 2011, 2011, 640309. [Google Scholar] [CrossRef] [PubMed]
- Launois, P.; DeLeys, R.; Niang, M.N.; Drowart, A.; Andrien, M.; Dierckx, P.; Cartel, J.L.; Sarthou, J.L.; Van Vooren, J.P.; Huygen, K. T-cell-epitope mapping of the major secreted mycobacterial antigen Ag85A in tuberculosis and leprosy. Infect. Immun. 1994, 62, 3679–3687. [Google Scholar] [CrossRef] [Green Version]
- Honda, M.; Wang, R.; Kong, W.P.; Kanekiyo, M.; Akahata, W.; Xu, L.; Matsuo, K.; Natarajan, K.; Robinson, H.; Asher, T.E.; et al. Different vaccine vectors delivering the same antigen elicit CD8+ T cell responses with distinct clonotype and epitope specificity. J. Immunol. 2009, 183, 2425–2434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kariyone, A.; Tamura, T.; Kano, H.; Iwakura, Y.; Takeda, K.; Akira, S.; Takatsu, K. Immunogenicity of Peptide-25 of Ag85B in Th1 development: Role of IFN-gamma. Int. Immunol. 2003, 15, 1183–1194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katila, M.L.; Brander, E.; Backman, A. Neonatal BCG vaccination and mycobacterial cervical adenitis in childhood. Tubercle 1987, 68, 291–296. [Google Scholar] [CrossRef]
- Romanus, V.; Hallander, H.O.; Wahlen, P.; Olinder-Nielsen, A.M.; Magnusson, P.H.; Juhlin, I. Atypical mycobacteria in extrapulmonary disease among children. Incidence in Sweden from 1969 to 1990, related to changing BCG-vaccination coverage. Tuber. Lung Dis. 1995, 76, 300–310. [Google Scholar] [CrossRef]
- Trnka, L.; Dankova, D.; Svandova, E. Six years’ experience with the discontinuation of BCG vaccination. 4. Protective effect of BCG vaccination against the Mycobacterium avium intracellulare complex. Tuber. Lung Dis. 1994, 75, 348–352. [Google Scholar] [CrossRef]
- Ottenhoff, T.H.; Kaufmann, S.H. Vaccines against tuberculosis: Where are we and where do we need to go? PLoS Pathog. 2012, 8, e1002607. [Google Scholar] [CrossRef]
- Ryan, A.A.; Nambiar, J.K.; Wozniak, T.M.; Roediger, B.; Shklovskaya, E.; Britton, W.J.; Fazekas de St Groth, B.; Triccas, J.A. Antigen load governs the differential priming of CD8 T cells in response to the bacille Calmette Guerin vaccine or Mycobacterium tuberculosis infection. J. Immunol. 2009, 182, 7172–7177. [Google Scholar] [CrossRef] [Green Version]
- Honda, M.; Matsuo, K.; Nakasone, T.; Okamoto, Y.; Yoshizaki, H.; Kitamura, K.; Sugiura, W.; Watanabe, K.; Fukushima, Y.; Haga, S.; et al. Protective immune responses induced by secretion of a chimeric soluble protein from a recombinant Mycobacterium bovis bacillus Calmette-Guerin vector candidate vaccine for human immunodeficiency virus type 1 in small animals. Proc. Natl. Acad. Sci. USA 1995, 92, 10693–10697. [Google Scholar] [CrossRef] [Green Version]
- Mazzaccaro, R.J.; Gedde, M.; Jensen, E.R.; van Santen, H.M.; Ploegh, H.L.; Rock, K.L.; Bloom, B.R. Major histocompatibility class I presentation of soluble antigen facilitated by Mycobacterium tuberculosis infection. Proc. Natl. Acad. Sci. USA 1996, 93, 11786–11791. [Google Scholar] [CrossRef] [Green Version]
- Kwon, B.E.; Ahn, J.H.; Park, E.K.; Jeong, H.; Lee, H.J.; Jung, Y.J.; Shin, S.J.; Jeong, H.S.; Yoo, J.S.; Shin, E.; et al. B Cell-Based Vaccine Transduced With ESAT6-Expressing Vaccinia Virus and Presenting alpha-Galactosylceramide Is a Novel Vaccine Candidate against ESAT6-Expressing Mycobacterial Diseases. Front. Immunol. 2019, 10, 2542. [Google Scholar] [CrossRef] [Green Version]
- Winthrop, K.; Rivera, A.; Engelmann, F.; Rose, S.; Lewis, A.; Ku, J.; Bermudez, L.; Messaoudi, I. A Rhesus Macaque Model of Pulmonary Nontuberculous Mycobacterial Disease. Am. J. Respir. Cell Mol. Biol. 2016, 54, 170–176. [Google Scholar] [CrossRef]
- Gilbertson, B.; Germano, S.; Steele, P.; Turner, S.; Fazekas de St Groth, B.; Cheers, C. Bystander activation of CD8+ T lymphocytes during experimental mycobacterial infection. Infect. Immun. 2004, 72, 6884–6891. [Google Scholar] [CrossRef] [Green Version]
- Stabel, J.R.; Waters, W.R.; Bannantine, J.P.; Palmer, M.V. Disparate host immunity to Mycobacterium avium subsp. paratuberculosis antigens in calves inoculated with M. avium subsp. paratuberculosis, M. avium subsp. avium, M. kansasii, and M. bovis. Clin. Vaccine Immunol. 2013, 20, 848–857. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.Y.; Huang, D.; Wang, R.C.; Shen, L.; Zeng, G.; Yao, S.; Shen, Y.; Halliday, L.; Fortman, J.; McAllister, M.; et al. A critical role for CD8 T cells in a nonhuman primate model of tuberculosis. PLoS Pathog. 2009, 5, e1000392. [Google Scholar] [CrossRef]
- Woodworth, J.S.; Behar, S.M. Mycobacterium tuberculosis-specific CD8+ T cells and their role in immunity. Crit. Rev. Immunol. 2006, 26, 317–352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bermudez, L.E.; Petrofsky, M. Host defense against Mycobacterium avium does not have an absolute requirement for major histocompatibility complex class I-restricted T cells. Infect. Immun. 1999, 67, 3108–3111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chapman, R.; Bourn, W.R.; Shephard, E.; Stutz, H.; Douglass, N.; Mgwebi, T.; Meyers, A.; Chin’ombe, N.; Williamson, A.L. The use of directed evolution to create a stable and immunogenic recombinant BCG expressing a modified HIV-1 Gag antigen. PLoS ONE 2014, 9, e103314. [Google Scholar] [CrossRef] [Green Version]
- Kanekiyo, M.; Matsuo, K.; Hamatake, M.; Hamano, T.; Ohsu, T.; Matsumoto, S.; Yamada, T.; Yamazaki, S.; Hasegawa, A.; Yamamoto, N.; et al. Mycobacterial codon optimization enhances antigen expression and virus-specific immune responses in recombinant Mycobacterium bovis bacille Calmette-Guerin expressing human immunodeficiency virus type 1 Gag. J. Virol. 2005, 79, 8716–8723. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ami, Y.; Izumi, Y.; Matsuo, K.; Someya, K.; Kanekiyo, M.; Horibata, S.; Yoshino, N.; Sakai, K.; Shinohara, K.; Matsumoto, S.; et al. Priming-boosting vaccination with recombinant Mycobacterium bovis bacillus Calmette-Guerin and a nonreplicating vaccinia virus recombinant leads to long-lasting and effective immunity. J. Virol. 2005, 79, 12871–12879. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Someya, K.; Ami, Y.; Nakasone, T.; Izumi, Y.; Matsuo, K.; Horibata, S.; Xin, K.Q.; Yamamoto, H.; Okuda, K.; Yamamoto, N.; et al. Induction of positive cellular and humoral immune responses by a prime-boost vaccine encoded with simian immunodeficiency virus gag/pol. J. Immunol. 2006, 176, 1784–1795. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kilpelainen, A.; Saubi, N.; Guitart, N.; Olvera, A.; Hanke, T.; Brander, C.; Joseph, J. Recombinant BCG Expressing HTI Prime and Recombinant ChAdOx1 Boost Is Safe and Elicits HIV-1-Specific T-Cell Responses in BALB/c Mice. Vaccines 2019, 7, 78. [Google Scholar] [CrossRef] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Komine-Aizawa, S.; Mizuno, S.; Matsuo, K.; Namiki, T.; Hayakawa, S.; Honda, M. Recombinant BCG-Prime and DNA-Boost Immunization Confers Mice with Enhanced Protection against Mycobacterium kansasii. Vaccines 2021, 9, 1260. https://doi.org/10.3390/vaccines9111260
Komine-Aizawa S, Mizuno S, Matsuo K, Namiki T, Hayakawa S, Honda M. Recombinant BCG-Prime and DNA-Boost Immunization Confers Mice with Enhanced Protection against Mycobacterium kansasii. Vaccines. 2021; 9(11):1260. https://doi.org/10.3390/vaccines9111260
Chicago/Turabian StyleKomine-Aizawa, Shihoko, Satoru Mizuno, Kazuhiro Matsuo, Takahiro Namiki, Satoshi Hayakawa, and Mitsuo Honda. 2021. "Recombinant BCG-Prime and DNA-Boost Immunization Confers Mice with Enhanced Protection against Mycobacterium kansasii" Vaccines 9, no. 11: 1260. https://doi.org/10.3390/vaccines9111260
APA StyleKomine-Aizawa, S., Mizuno, S., Matsuo, K., Namiki, T., Hayakawa, S., & Honda, M. (2021). Recombinant BCG-Prime and DNA-Boost Immunization Confers Mice with Enhanced Protection against Mycobacterium kansasii. Vaccines, 9(11), 1260. https://doi.org/10.3390/vaccines9111260