
ISCOM-like Nanoparticles Formulated with Quillaja brasiliensis Saponins Are Promising Adjuvants for Seasonal Influenza Vaccines

 ,
,  , , and
, , and 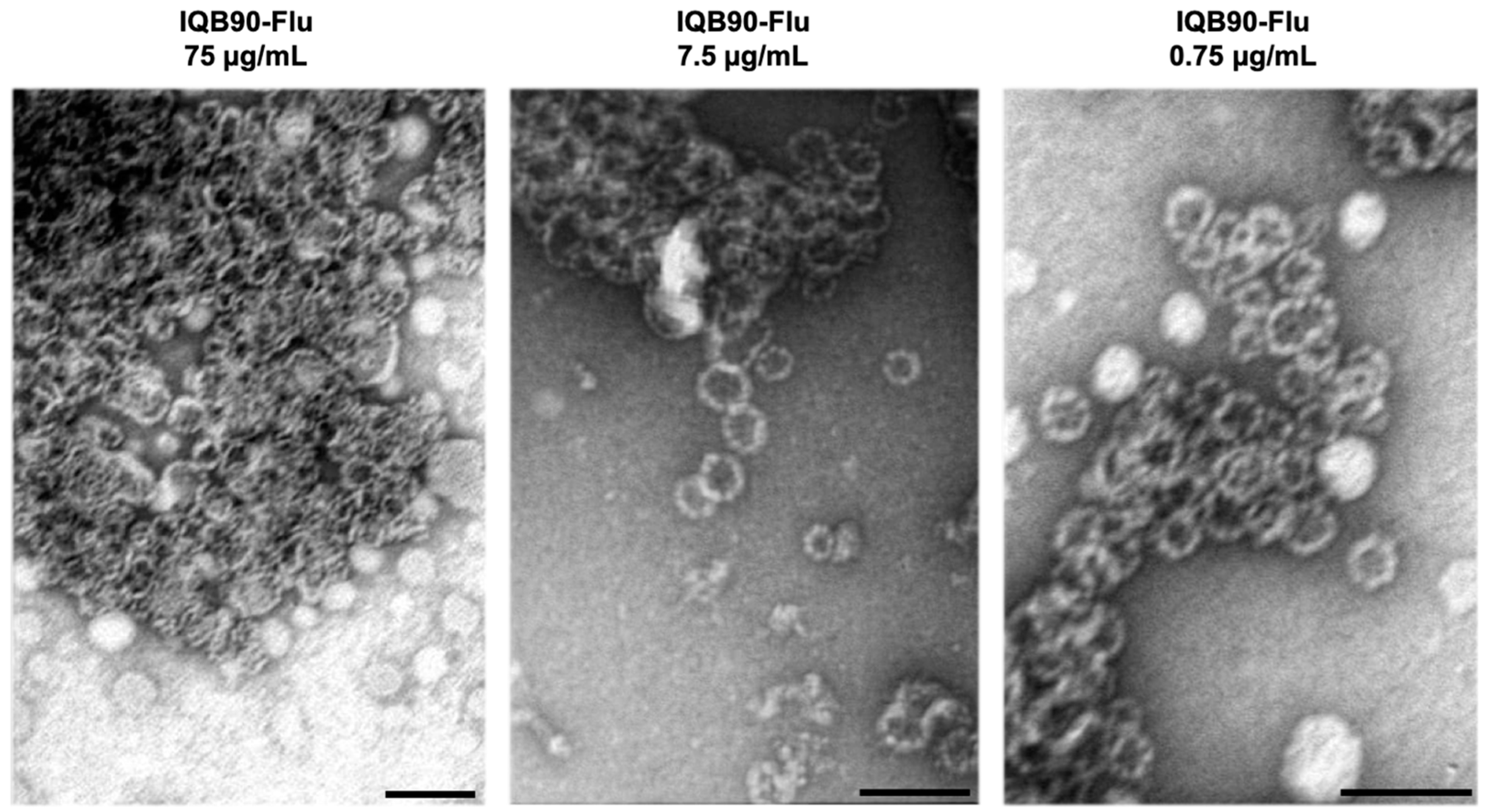{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Vaccine Adjuvants
2.2. Antigen Preparation
2.3. Experimental Vaccine Preparations
2.3.1. Unadjuvanted Influenza Vaccine
2.3.2. IQB90 Adjuvanted Influenza Vaccine
2.4. Transmission Electron Microscopy (TEM)
2.5. Mouse Immunizations and Sample Collection
2.6. Delayed-Type Hypersensitivity (DTH) Assay
2.7. Antibody-Level Determination Assays
2.8. ELISpot Assay for Influenza-Specific IgG Antibody Secreting Cells
2.9. Mouse Challenge
2.10. Statistical Analyses
3. Results and Discussion
3.1. QB-90 ISCOMs Formulated with Influenza Antigen (IQB90-Flu) Are Non-Toxic Cage-Like Structures
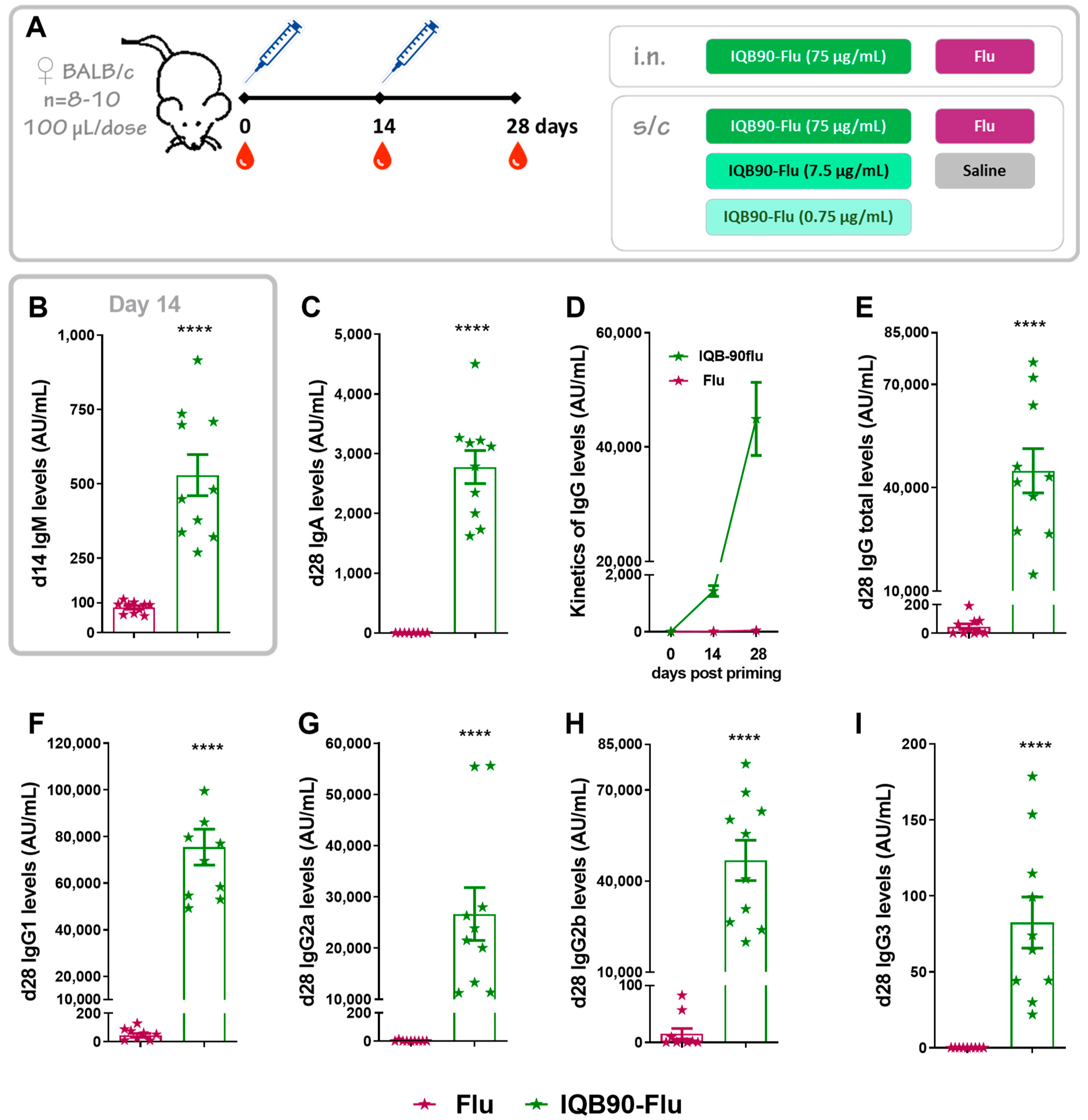
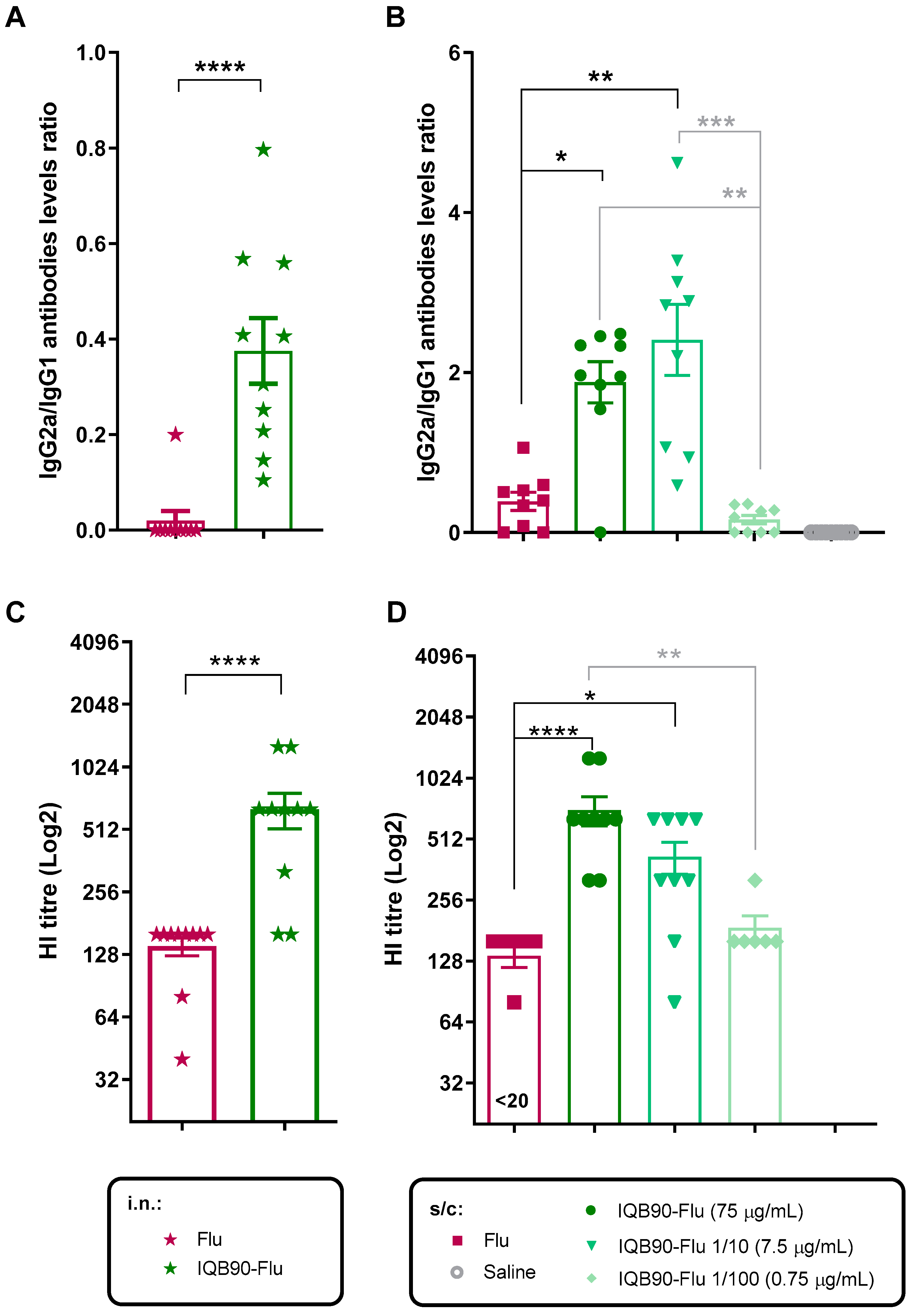
3.2. Intranasal or Subcutaneous Administration of IQB90-Flu Vaccine Induced Higher Antibody Levels Than the Commercial Influenza Vaccine in Mice
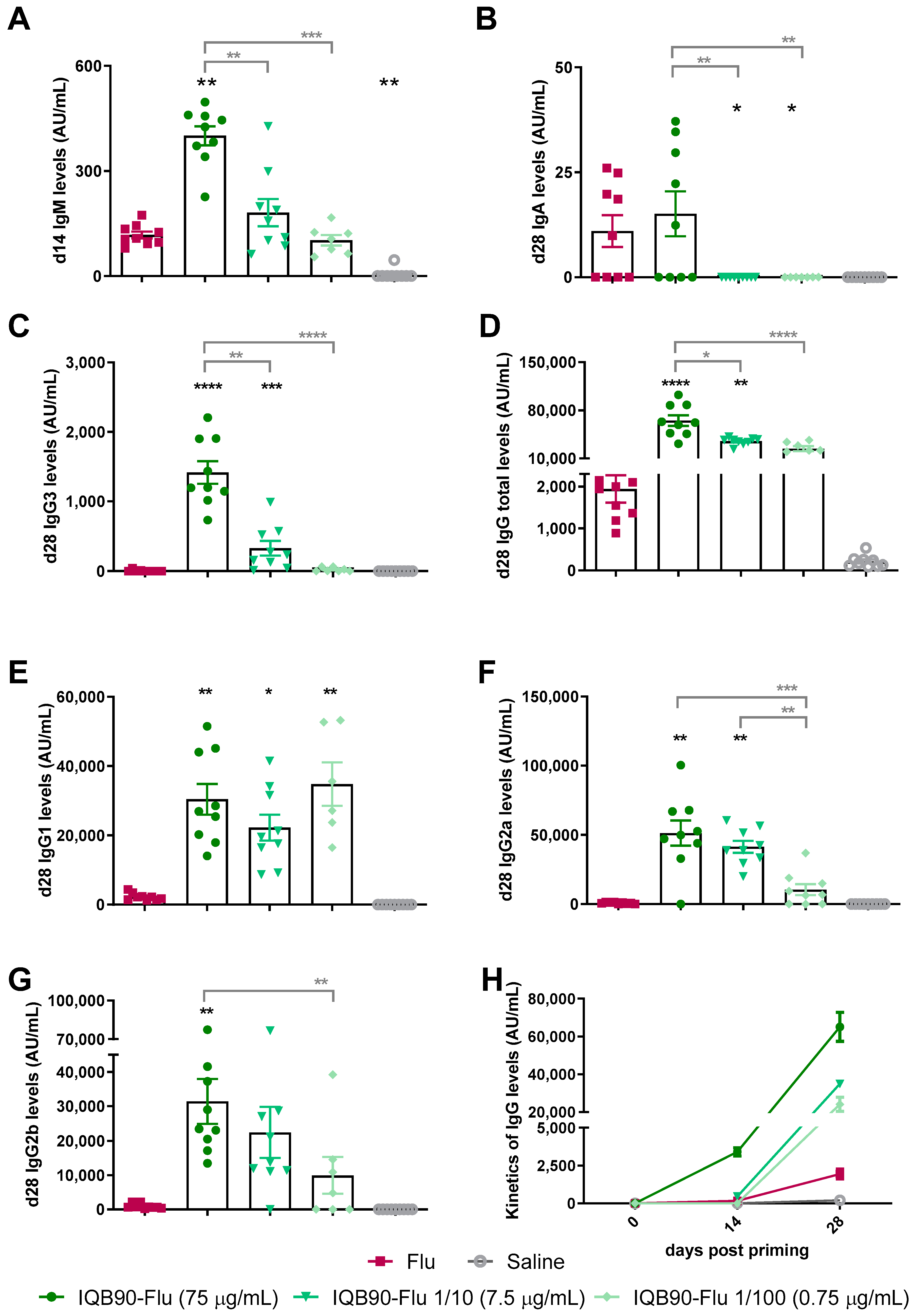
3.3. IQB90-Flu Conferred a Ten-Fold Dose-Sparing Effect in a Split-Inactivated Influenza Vaccine
3.4. The Immune Response Elicited by Subcutaneous Injection of IQB90-Flu Vaccines Was Adequate for Conferring Protection against Influenza Infection in Mice
3.5. IQB90-Flu Vaccines Trended to Protect Mice from Influenza
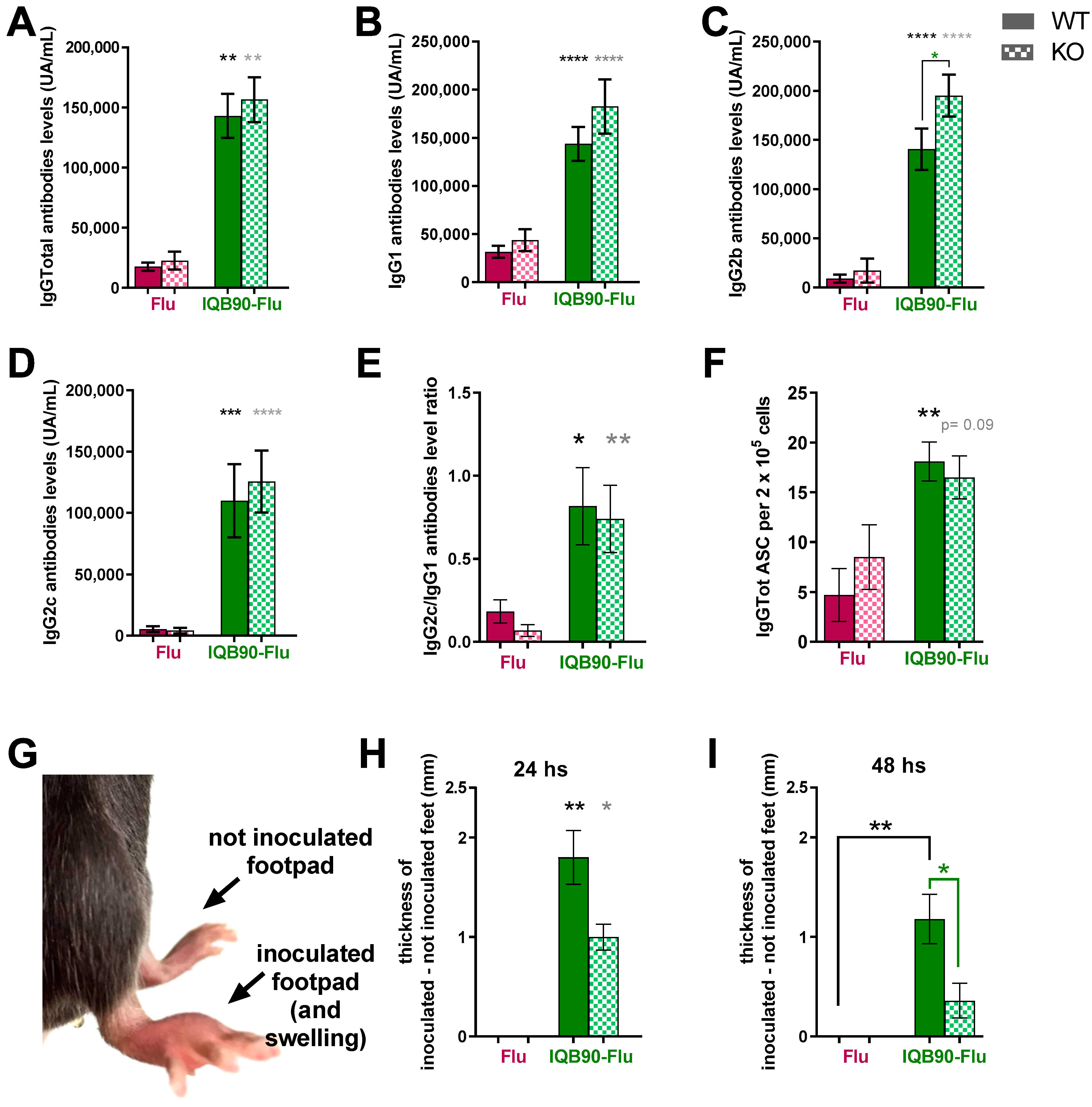
3.6. Subcutaneous Administration of IQB90 Elicited High Antibody Levels and Specific ASC in Wild-Type and Casp-1/11 Knockout Mice
3.7. Casp-1/11 ko Mice Vaccinated with IQB90-Flu Developed a Lower Delayed-Type Hypersensitivity (DTH) Response Than wt Mice
4. Concluding Remarks and Perspectives
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- World Health Organisation. Global Influenza Strategy Summary 2019–2030 Influenza. Glob. Influ. Strateg. 2019–2030 Influ. 2019, 1, 2. [Google Scholar]
- Paules, C.I.; Fauci, A.S. Influenza Vaccines: Good, But We Can Do Better. J. Infect. Dis. 2019, 219, S1–S4. [Google Scholar] [CrossRef]
- Hobson, D.; Curry, R.L.; Beare, A.S.; Ward-Gardner, A. The role of serum haemagglutination-inhibiting antibody in protection against challenge infection with influenza A2 and B viruses. J. Hyg. Lond. 1972, 70, 767–777. [Google Scholar] [CrossRef] [Green Version]
- Seibert, C.W.; Rahmat, S.; Krause, J.C.; Eggink, D.; Albrecht, R.A.; Goff, P.H.; Krammer, F.; Duty, J.A.; Bouvier, N.M.; Garcia-Sastre, A.; et al. Recombinant IgA Is Sufficient To Prevent Influenza Virus Transmission in Guinea Pigs. J. Virol. 2013, 87, 7793–7804. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tregoning, J.S.; Russell, R.F.; Kinnear, E. Adjuvanted influenza vaccines. Hum. Vaccines Immunother. 2018, 14, 550–564. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Hagan, D.T.; Lodaya, R.N.; Lofano, G. The continued advance of vaccine adjuvants–‘we can work it out’. Semin. Immunol. 2020, 101426. [Google Scholar] [CrossRef] [PubMed]
- Dalsgaard, K. Saponin Adjuvants-Isolation of Substance/tom Quillaja saponaria Molina with Adjuvant Activity in Foot-and-Mouth Disease Vaccine. Arch. Gesamte Virusforsch. 1974, 254, 243–254. [Google Scholar] [CrossRef]
- Magedans, Y.V.S.; Yendo, A.C.A.; de Costa, F.; Gosmann, G.; Arthur, G. Foamy matters: An update on Quillaja saponins and their use as immunoadjuvants. Future Med. Chem. 2019, 11, 1485–1499. [Google Scholar] [CrossRef]
- Lacaille-Dubois, M.A. Updated insights into the mechanism of action and clinical profile of the immunoadjuvant QS-21: A review. Phytomedicine 2019, 60, 152905. [Google Scholar] [CrossRef]
- Keech, C.; Albert, G.; Cho, I.; Robertson, A.; Reed, P.; Neal, S.; Plested, J.S.; Zhu, M.; Clark, S.C.; Zhou, H.; et al. Phase 1–2 Trial of a SARS-CoV-2 Recombinant Spike Protein Nanoparticle Vaccine. N. Engl. J. Med. 2020, 383, 2320–2332. [Google Scholar] [CrossRef]
- Heath, P.T.; Galiza, E.P.; Baxter, D.N.; Boffito, M.; Browne, D.; Burns, F.; Chadwick, D.R.; Clark, R.; Cosgrove, C.; Galloway, J.; et al. Safety and Efficacy of NVX-CoV2373 Covid-19 Vaccine. N. Engl. J. Med. 2021, 385, 1172–1183. [Google Scholar] [CrossRef]
- Morein, B.; Sundquist, B.; Höglund, S.; Dalsgaard, K.; Osterhaus, A. Iscom, a novel structure for antigenic presentation of membrane proteins from enveloped viruses. Nature 1984, 308, 457–460. [Google Scholar] [CrossRef] [PubMed]
- Cibulski, S.P.; Silveira, F.; Mourglia-Ettlin, G.; Teixeira, T.F.; dos Santos, H.F.; Yendo, A.C.; de Costa, F.; Fett-Neto, A.G.; Gosmann, G.; Roehe, P.M. Quillaja brasiliensis saponins induce robust humoral and cellular responses in a bovine viral diarrhea virus vaccine in mice. Comp. Immunol. Microbiol. Infect. Dis. 2016, 45, 1–8. [Google Scholar] [CrossRef]
- De Costa, F.; Yendo, A.C.A.; Cibulski, S.P.; Fleck, J.D.; Roehe, P.M.; Spilki, F.R.; Gosmann, G.; Fett-Neto, A.G. Alternative inactivated poliovirus vaccines adjuvanted with Quillaja brasiliensis or Quil-A saponins are equally effective in inducing specific immune responses. PLoS ONE 2014, 9, e105374. [Google Scholar] [CrossRef] [Green Version]
- Yendo, A.C.A.; de Costa, F.; Cibulski, S.P.; Teixeira, T.F.; Colling, L.C.; Mastrogiovanni, M.; Soule, S.; Roehe, P.M.; Gosmann, G.; Ferreira, F.A.; et al. A rabies vaccine adjuvanted with saponins from leaves of the soap tree (Quillaja brasiliensis) induces specific immune responses and protects against lethal challenge. Vaccine 2016, 34, 2305–2311. [Google Scholar] [CrossRef]
- Silveira, F.; Cibulski, S.P.; Varela, A.P.; Marqu??s, J.M.; Chabalgoity, A.; de Costa, F.; Yendo, A.C.A.; Gosmann, G.; Roehe, P.M.; Fern??ndez, C.; et al. Quillaja brasiliensis saponins are less toxic than Quil A and have similar properties when used as an adjuvant for a viral antigen preparation. Vaccine 2011, 29, 9177–9182. [Google Scholar] [CrossRef] [Green Version]
- Cibulski, S.; Teixeira, T.F.; Varela, A.P.M.; de Lima, M.F.; Casanova, G.; Nascimento, Y.M.; Fechine Tavares, J.; da Silva, M.S.; Sesterheim, P.; Souza, D.O.; et al. IMXQB-80: A Quillaja brasiliensis saponin-based nanoadjuvant enhances Zika virus specific immune responses in mice. Vaccine 2021, 39, 571–579. [Google Scholar] [CrossRef] [PubMed]
- Cibulski, S.; Rivera-Patron, M.; Suárez, N.; Pirez, M.; Rossi, S.; Yendo, A.C.; de Costa, F.; Gosmann, G.; Fett-Neto, A.; Roehe, P.M.; et al. Leaf saponins of Quillaja brasiliensis enhance long-term specific immune responses and promote dose-sparing effect in BVDV experimental vaccines. Vaccine 2018, 36, 55–65. [Google Scholar] [CrossRef]
- Fleck, J.D.; De Costa, F.; Yendo, A.C.A.; Segalin, J.; Dalla Costa, T.C.T.; Fett-Neto, A.G.; Gosmann, G. Determination of new immunoadjuvant saponin named QB-90, and analysis of its organ-specific distribution in Quillaja brasiliensis by HPLC. Nat. Prod. Res. 2013, 27, 907–910. [Google Scholar] [CrossRef]
- Luebert, F. Taxonomy and distribution of the genus Quillaja Molina (Quillajaceae). Feddes Repert. 2014, 124, 157–162. [Google Scholar] [CrossRef]
- Fleck, J.D.; Kauffmann, C.; Spilki, F.; Lencina, C.L.; Roehe, P.M.; Gosmann, G. Adjuvant activity of Quillaja brasiliensis saponins on the immune responses to bovine herpesvirus type 1 in mice. Vaccine 2006, 24, 7129–7134. [Google Scholar] [CrossRef] [PubMed]
- Cibulski, S.P.; Mourglia-Ettlin, G.; Teixeira, T.F.; Quirici, L.; Roehe, P.M.; Ferreira, F.; Silveira, F. Novel ISCOMs from Quillaja brasiliensis saponins induce mucosal and systemic antibody production, T-cell responses and improved antigen uptake. Vaccine 2016, 34, 1162–1171. [Google Scholar] [CrossRef] [PubMed]
- Cibulski, S.P.; Rivera-Patron, M.; Mourglia-Ettlin, G.; Casaravilla, C.; Yendo, A.C.; Fett-Neto, A.G.; Chabalgoity, J.A.; Moreno, M.; Roehe, P.M.; Silveira, F. Quillaja brasiliensis saponin-based nanoparticulate adjuvants are capable of triggering early immune responses. Sci. Rep. 2018, 8, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Yendo, A.; de Costa, F.; Kauffmann, C.; Fleck, J.; Gosmann, G.; Fett-Neto, A. Purification of an Immunoadjuvant Saponin Fraction from Quillaja brasiliensis Leaves by Reversed-Phase Silica Gel Chromatography. In Vaccine Adjuvants: Methods and Protocols; Fox, C.B., Ed.; Springer: New York, NY, USA, 2017; Volume 1494, pp. 87–93. [Google Scholar] [CrossRef]
- Lendemans, D.G.; Myschik, J.; Hook, S.; Rades, T. Immuno-stimulating complexes prepared by ethanol injection. J. Pharm. Pharmacol. 2005, 57, 729–733. [Google Scholar] [CrossRef]
- Lv, J.; Wang, D.; Hua, Y.H.; Pei, S.J.; Wang, J.; Hu, W.W.; Wang, X.L.; Jia, N.; Jiang, Q.S. Pulmonary immune responses to 2009 pandemic influenza A (H1N1) virus in mice. BMC Infect. Dis. 2014, 14, 197. [Google Scholar] [CrossRef] [Green Version]
- Sun, H.X.; Xie, Y.; Ye, Y.P. ISCOMs and ISCOMATRIXTM. Vaccine 2009, 27, 4388–4401. [Google Scholar] [CrossRef]
- McKenzie, A.; Watt, M.; Gittleson, C. ISCOMATRIX vaccines: Safety in human clinical studies. Hum. Vaccin. 2010, 6, 237–246. [Google Scholar] [CrossRef]
- Cebon, J.S.; Gore, M.; Thompson, J.F.; Davis, I.D.; McArthur, G.A.; Walpole, E.; Smithers, M.; Cerundolo, V.; Dunbar, P.R.; MacGregor, D.; et al. Results of a randomized, double-blind phase II clinical trial of NY-ESO-1 vaccine with ISCOMATRIX adjuvant versus ISCOMATRIX alone in participants with high-risk resected melanoma. J. Immunother. Cancer 2020, 8, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Bigaeva, E.; van Doorn, E.; Liu, H.; Hak, E. Meta-Analysis on Randomized Controlled Trials of Vaccines with QS-21 or ISCOMATRIX Adjuvant: Safety and Tolerability. PLoS ONE 2016, 11, e0154757. [Google Scholar] [CrossRef] [Green Version]
- Holmgren, J.; Czerkinsky, C. Mucosal immunity and vaccines. Nat. Med. 2005, 11, S45. [Google Scholar] [CrossRef] [PubMed]
- De Magistris, M.T. Mucosal delivery of vaccine antigens and its advantages in pediatrics. Adv. Drug Deliv. Rev. 2006, 58, 52–67. [Google Scholar] [CrossRef] [PubMed]
- Mitragotri, S. Immunization without needles. Nat. Rev. Immunol. 2005, 5, 905–916. [Google Scholar] [CrossRef] [PubMed]
- De Temmerman, M.L.; Rejman, J.; Demeester, J.; Irvine, D.J.; Gander, B.; De Smedt, S.C. Particulate vaccines: On the quest for optimal delivery and immune response. Drug Discov. Today 2011, 16, 569–582. [Google Scholar] [CrossRef]
- Kunda, N.K.; Somavarapu, S.; Gordon, S.B.; Hutcheon, G.A.; Saleem, I.Y. Nanocarriers targeting dendritic cells for pulmonary vaccine delivery. Pharm. Res. 2013, 30, 325–341. [Google Scholar] [CrossRef] [PubMed]
- Sjölander, S.; Drane, D.; Davis, R.; Beezum, L.; Pearse, M.; Cox, J. Intranasal immunisation with influenza-ISCOM induces strong mucosal as well as systemic antibody and cytotoxic T-lymphocyte responses. Vaccine 2001, 19, 4072–4080. [Google Scholar] [CrossRef]
- Tomar, J.; Patil, H.P.; Bracho, G.; Tonnis, W.F.; Frijlink, H.W.; Petrovsky, N.; Vanbever, R.; Huckriede, A.; Hinrichs, W.L.J. Advax augments B and T cell responses upon influenza vaccination via the respiratory tract and enables complete protection of mice against lethal influenza virus challenge. J. Control Release 2018, 288, 199–211. [Google Scholar] [CrossRef] [PubMed]
- Podda, A. The adjuvanted influenza vaccines with novel adjuvants: Experience with the MF59-adjuvanted vaccine. Vaccine 2001, 19, 2673–2680. [Google Scholar] [CrossRef]
- Krammer, F.; Smith, G.J.D.; Fouchier, R.A.M.; Peiris, M.; Kedzierska, K.; Doherty, P.C.; Palese, P.; Shaw, M.L.; Treanor, J.; Webster, R.G.; et al. Influenza. Nat. Rev. Dis. Prim. 2018, 4, 1–21. [Google Scholar] [CrossRef]
- Reber, A.; Katz, J. Immunological assessment of influenza vaccines and immune correlates of protection. Expert Rev. Vaccines 2013, 12, 519–536. [Google Scholar] [CrossRef]
- Wilson, N.S.; Duewell, P.; Yang, B.; Li, Y.; Marsters, S.; Koernig, S.; Latz, E.; Maraskovsky, E.; Morelli, A.B.; Schnurr, M.; et al. Inflammasome-Dependent and -Independent IL-18 Production Mediates Immunity to the ISCOMATRIX Adjuvant. J. Immunol. 2014, 192, 3259–3268. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Willingham, S.B.; Ting, J.P.-Y.; Re, F. Cutting Edge: Inflammasome Activation by Alum and Alum’s Adjuvant Effect Are Mediated by NLRP3. J. Immunol. 2008, 181, 17–21. [Google Scholar] [CrossRef] [Green Version]
- Detienne, S.; Welsby, I.; Collignon, C.; Wouters, S.; Coccia, M.; Delhaye, S.; Van Maele, L.; Thomas, S.; Swertvaegher, M.; Detavernier, A.; et al. Central Role of CD169(+) Lymph Node Resident Macrophages in the Adjuvanticity of the QS-21 Component of AS01. Sci. Rep. 2016, 6, 39475. [Google Scholar] [CrossRef] [PubMed]
- Knudsen, N.P.H.; Olsen, A.; Buonsanti, C.; Follmann, F.; Zhang, Y.; Coler, R.N.; Fox, C.B.; Meinke, A.; D’Oro, U.; Casini, D.; et al. Different human vaccine adjuvants promote distinct antigen-independent immunological signatures tailored to different pathogens. Sci. Rep. 2016, 6, 19570. [Google Scholar] [CrossRef]
- Fernández-Tejada, A.; Chea, E.K.; George, C.; Gardner, J.R.; Livingston, P.O.; Ragupathi, G.; Tan, D.S.; Gin, D.Y. Design, synthesis, and immunologic evaluation of vaccine adjuvant conjugates based on QS-21 and tucaresol. Bioorganic Med. Chem. 2014, 22, 5917–5923. [Google Scholar] [CrossRef] [Green Version]
- Wallace, F.; Bennadji, Z.; Ferreira, F.; Olivaro, C. Structural characterisation of new immunoadjuvant saponins from leaves and the first study of saponins from the bark of Quillaja brasiliensis by liquid chromatography electrospray ionisation ion trap mass spectrometry. Phytochem. Anal. 2019, 30, 644–652. [Google Scholar] [CrossRef] [PubMed]
- Wallace, F.; Bennadji, Z.; Ferreira, F.; Olivaro, C. Analysis of an immunoadjuvant saponin fraction from Quillaja brasiliensis leaves by electrospray ionization ion trap multiple-stage mass spectrometry. Phytochem. Lett. 2017, 20, 228–233. [Google Scholar] [CrossRef]
- Fleck, J.D.; Schwwambach, J.; Almeida, M.E.; Yendo, A.C.A.; de Costa, F.; Gossmann, G.; Fett-Neto, A.G. Immunoadjuvant Saponin Production in Seedlings and Micropropagated Plants of Quillaja brasiliensis. Soc. Vitr. Biol. 2009, 45, 715–720. [Google Scholar] [CrossRef]
- Fernández-Tejada, A.; Tan, D.S.; Gin, D.Y. Development of Improved Vaccine Adjuvants Based on the Saponin Natural Product QS-21 through Chemical Synthesis. Acc. Chem. Res. 2016, 49, 1741–1756. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rivera-Patron, M.; Moreno, M.; Baz, M.; Roehe, P.M.; Cibulski, S.P.; Silveira, F. ISCOM-like Nanoparticles Formulated with Quillaja brasiliensis Saponins Are Promising Adjuvants for Seasonal Influenza Vaccines. Vaccines 2021, 9, 1350. https://doi.org/10.3390/vaccines9111350
Rivera-Patron M, Moreno M, Baz M, Roehe PM, Cibulski SP, Silveira F. ISCOM-like Nanoparticles Formulated with Quillaja brasiliensis Saponins Are Promising Adjuvants for Seasonal Influenza Vaccines. Vaccines. 2021; 9(11):1350. https://doi.org/10.3390/vaccines9111350
Chicago/Turabian StyleRivera-Patron, Mariana, María Moreno, Mariana Baz, Paulo M. Roehe, Samuel P. Cibulski, and Fernando Silveira. 2021. "ISCOM-like Nanoparticles Formulated with Quillaja brasiliensis Saponins Are Promising Adjuvants for Seasonal Influenza Vaccines" Vaccines 9, no. 11: 1350. https://doi.org/10.3390/vaccines9111350
APA StyleRivera-Patron, M., Moreno, M., Baz, M., Roehe, P. M., Cibulski, S. P., & Silveira, F. (2021). ISCOM-like Nanoparticles Formulated with Quillaja brasiliensis Saponins Are Promising Adjuvants for Seasonal Influenza Vaccines. Vaccines, 9(11), 1350. https://doi.org/10.3390/vaccines9111350