
Dry Formulation of Virus-Like Particles in Electrospun Nanofibers


 , ,
, ,  and
and
Abstract
:
{kind=link}
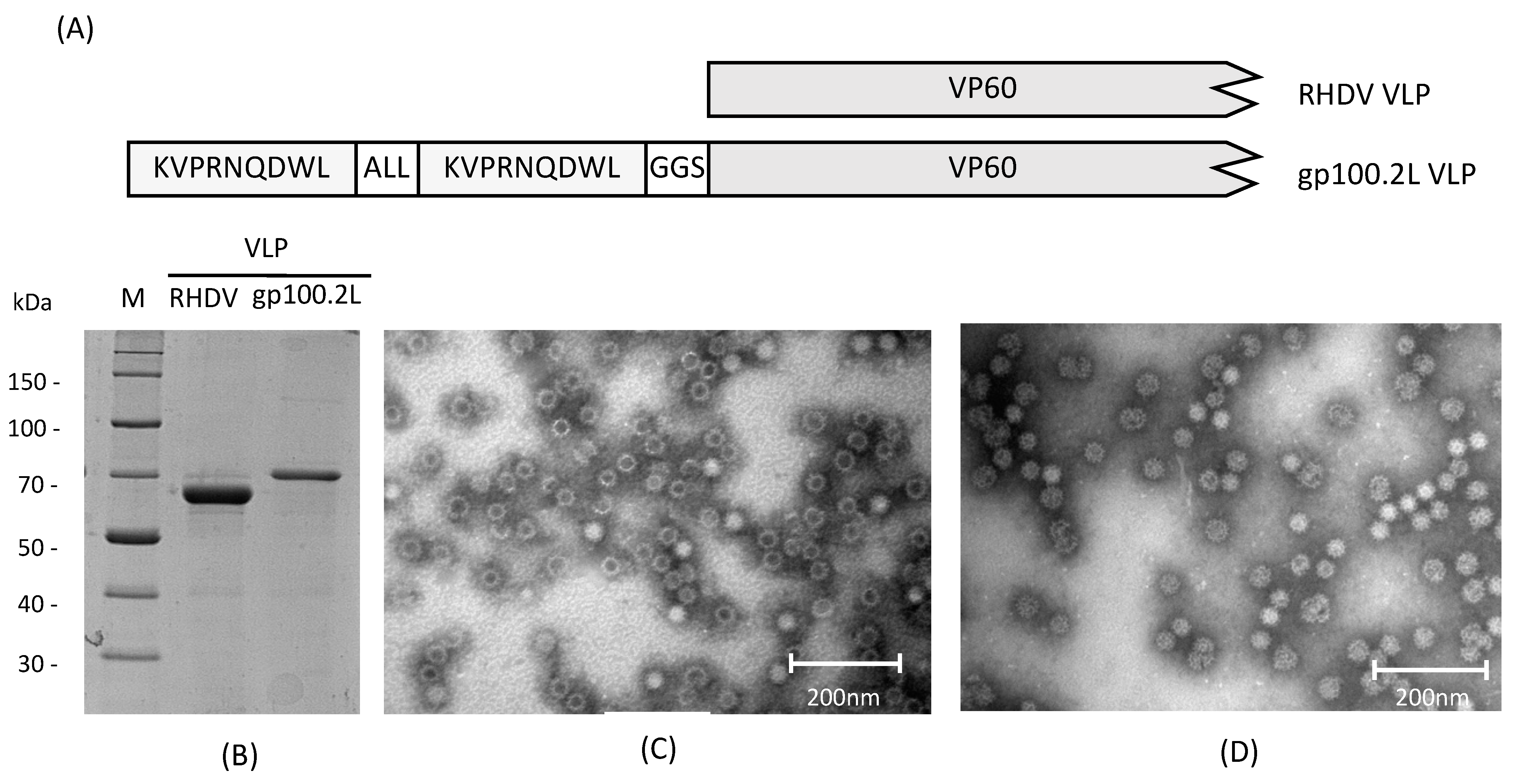{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Expression and Purification of VLP
2.2. Electrospinning
2.3. Electron Microscopy
2.4. Western Blot
2.5. Animals and Ethics
2.6. T-Cell Proliferation and Interferon-γ Production Assay
2.7. In Vivo Cytotoxicity
2.8. Serum Collection and ELISA
2.9. Statistical Analysis
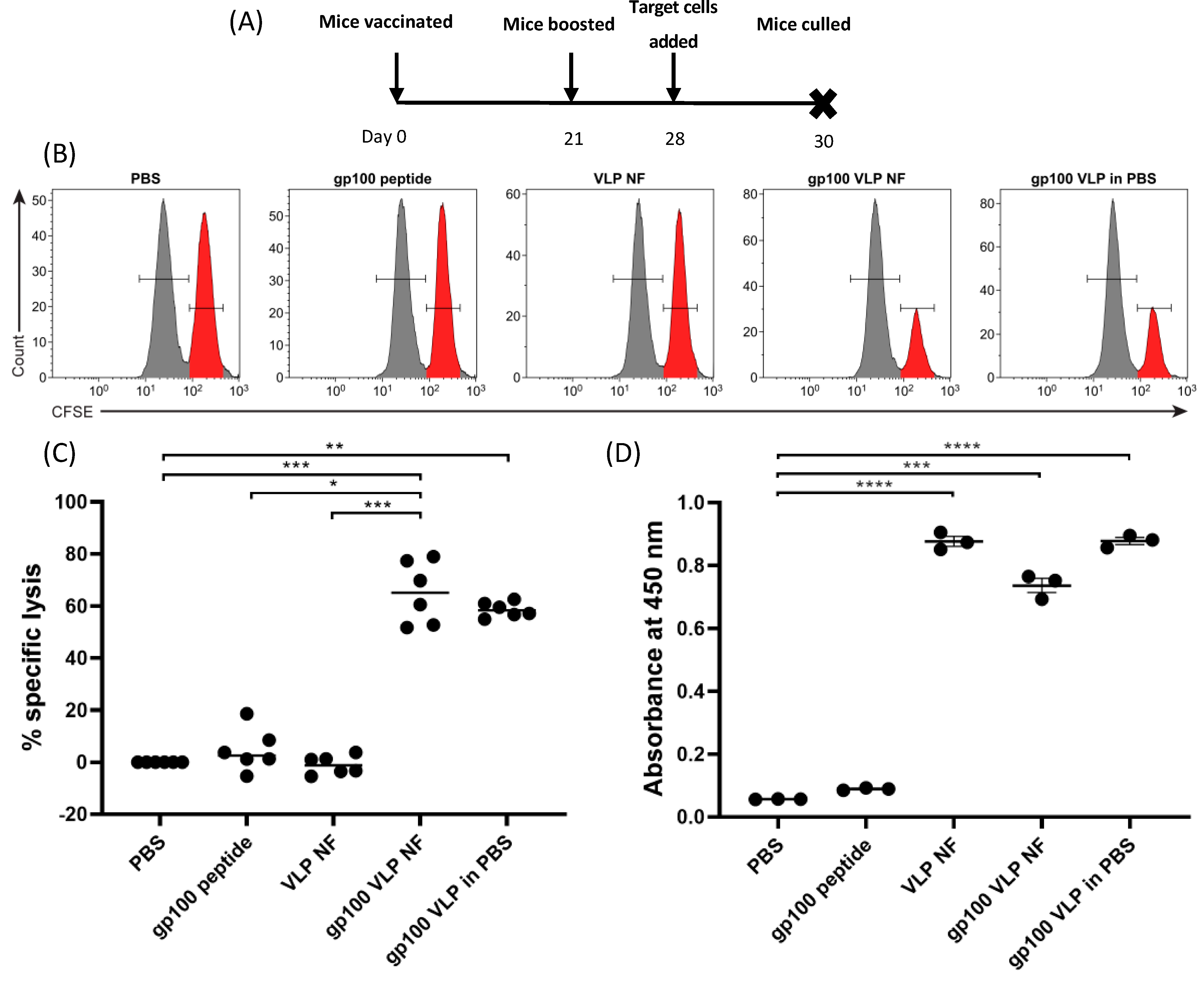
3. Results
3.1. VLP Expression and Purification
3.2. Incorporation of VLP into Nanofibers
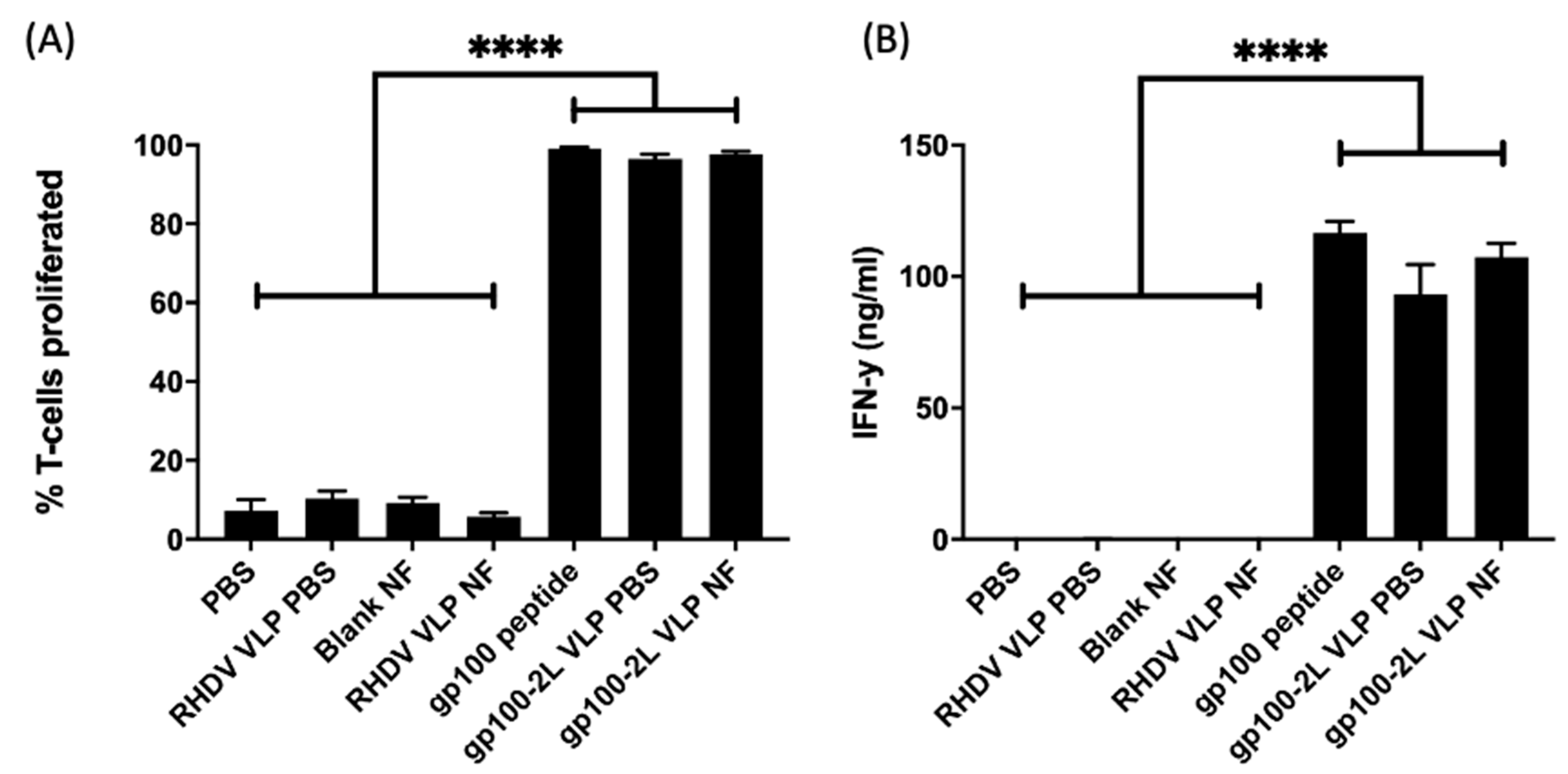
3.3. T-Cell Proliferation and IFN-γ Production Assays
3.4. In Vivo Cytotoxicity and Antibody Response
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Grgacic, E.V.; Anderson, D.A. Virus-like particles: Passport to immune recognition. Methods 2006, 40, 60–65. [Google Scholar] [CrossRef]
- Cimica, V.; Galarza, J.M. Adjuvant formulations for virus-like particle (VLP) based vaccines. J. Clin. Immunol. 2017, 183, 99–108. [Google Scholar] [CrossRef] [PubMed]
- Godi, A.; Bissett, S.L.; Miller, E.; Beddows, S. Relationship between Humoral Immune Responses against HPV16, HPV18, HPV31 and HPV45 in 12–15 Year Old Girls Receiving Cervarix® or Gardasil® Vaccine. PLoS ONE 2015, 10, e0140926. [Google Scholar]
- Pol, S.; Driss, F.; Michel, M.-L.; Nalpas, B.; Berthelot, P.; Brechot, C. Specific vaccine therapy in chronic hepatitis B infection. Lancet 1994, 344, 342. [Google Scholar] [CrossRef]
- Wu, X.; Chen, P.; Lin, H.; Hao, X.; Liang, Z. Hepatitis E virus: Current epidemiology and vaccine. Hum. Vaccines Immunother. 2016, 12, 2603–2610. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Wei, M.; Sun, G.; Wang, X.; Li, M.; Lin, Z.; Li, Z.; Li, Y.; Fang, M.; Zhang, J.; et al. Real-time stability of a hepatitis E vaccine (Hecolin(R)) demonstrated with potency assays and multifaceted physicochemical methods. Vaccine 2016, 34, 5871–5877. [Google Scholar] [CrossRef]
- Win, S.J.; McMillan, D.G.; Errington-Mais, F.; Ward, V.K.; Young, S.L.; Baird, M.A.; Melcher, A.A. Enhancing the immunogenicity of tumour lysate-loaded dendritic cell vaccines by conjugation to virus-like particles. Br. J. Cancer 2012, 106, 92–98. [Google Scholar] [CrossRef]
- Al-Barwani, F.; Donaldson, B.; Pelham, S.J.; Young, S.L.; Ward, V.K. Antigen delivery by virus-like particles for immunotherapeutic vaccination. Ther. Deliv. 2014, 5, 1223–1240. [Google Scholar] [CrossRef]
- Cubas, R.; Zhang, S.; Kwon, S.; Sevick-Muraca, E.M.; Li, M.; Chen, C.; Yao, Q. Virus-like particle (VLP) Lymphatic Trafficking and Immune Response Generation after Immunization by Different Routes. J. Immunother. 2009, 32, 118–128. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Cubas, R.; Li, M.; Chen, C.; Yao, Q. Virus-Like Particle Vaccine Activates Conventional B2 Cells and Promotes B Cell Differentiation to IgG2a Producing Plasma Cells. Mol. Immunol. 2009, 46, 1988–2001. [Google Scholar] [CrossRef] [Green Version]
- Matthias, D.M.; Robertson, J.; Garrison, M.M.; Newland, S.; Nelson, C. Freezing temperatures in the vaccine cold chain: A systematic literature review. Vaccine 2007, 25, 3980–3986. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Purssell, E. Reviewing the importance of the cold chain in the distribution of vaccines. Br. J. Community Nurs. 2015, 20, 481–486. [Google Scholar] [CrossRef] [PubMed]
- Donaldson, B.; Lateef, Z.; Walker, G.F.; Young, S.L.; Ward, V.K. Virus-like particle vaccines: Immunology and formulation for clinical translation. Expert Rev. Vaccines 2018, 17, 833–849. [Google Scholar] [CrossRef] [PubMed]
- Wirkas, T.; Toikilik, S.; Miller, N.; Morgan, C.; Clements, C.J. A vaccine cold chain freezing study in PNG highlights technology needs for hot climate countries. Vaccine 2007, 25, 691–697. [Google Scholar] [CrossRef]
- Maltesen, M.J.; van de Weert, M. Drying methods for protein pharmaceuticals. Drug Discov. Today Technol. 2008, 5, e81–e88. [Google Scholar] [CrossRef]
- Kanojia, G.; Have Rt Soema, P.C.; Frijlink, H.; Amorij, J.-P.; Kersten, G. Developments in the formulation and delivery of spray dried vaccines. Hum. Vaccines Immunother. 2017, 13, 2364–2378. [Google Scholar] [CrossRef]
- Lang, R.; Winter, G.; Vogt, L.; Zurcher, A.; Dorigo, B.; Schimmele, B. Rational design of a stable, freeze-dried virus-like particle-based vaccine formulation. Drug Dev. Ind. Pharm. 2009, 35, 83–97. [Google Scholar] [CrossRef]
- Czyż, M.; Pniewski, T. Thermostability of Freeze-Dried Plant-Made VLP-Based Vaccines. In Sustainable Drying Technologies; Olvera, J.d.R., Ed.; IntechOpen: London, UK, 2016. [Google Scholar]
- LeClair, D.A.; Cranston, E.D.; Xing, Z.; Thompson, M.R. Optimization of Spray Drying Conditions for Yield, Particle Size and Biological Activity of Thermally Stable Viral Vectors. Pharm. Res. 2016, 33, 2763–2776. [Google Scholar] [CrossRef] [Green Version]
- Mumenthaler, M.; Hsu, C.C.; Pearlman, R. Feasibility study on spray-drying protein pharmaceuticals: Recombinant human growth hormone and tissue-type plasminogen activator. Pharm. Res. 1994, 11, 12–20. [Google Scholar] [CrossRef]
- Webb, S.D.; Golledge, S.L.; Cleland, J.L.; Carpenter, J.F.; Randolph, T.W. Surface adsorption of recombinant human interferon-gamma in lyophilized and spray-lyophilized formulations. J. Pharm. Sci. 2002, 91, 1474–1487. [Google Scholar] [CrossRef]
- Saboo, S.; Tumban, E.; Peabody, J.; Wafula, D.; Peabody, D.S.; Chackerian, B.; Muttil, P. Optimized Formulation of a Thermostable Spray-Dried Virus-Like Particle Vaccine against Human Papillomavirus. Mol. Pharm. 2016, 13, 1646–1655. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Fernando, G.J.; Crichton, M.L.; Flaim, C.; Yukiko, S.R.; Fairmaid, E.J.; Corbett, H.J.; Primiero, C.A.; Ansaldo, A.B.; Frazer, I.H.; et al. Improving the reach of vaccines to low-resource regions, with a needle-free vaccine delivery device and long-term thermostabilization. J. Control. Release 2011, 152, 349–355. [Google Scholar] [CrossRef] [Green Version]
- Leung, V.; Mapletoft, J.; Zhang, A.; Lee, A.; Vahedi, F.; Chew, M.; Szewczyk, A.; Jahanshahi-Anbuhi, S.; Ang, J.; Cowbrough, B.; et al. Thermal Stabilization of Viral Vaccines in Low-Cost Sugar Films. Sci. Rep. 2019, 9, 7631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lovalenti, P.M.; Anderl, J.; Yee, L.; Nguyen, V.; Ghavami, B.; Ohtake, S.; Saxena, A.; Voss, T.; Truong-Le, V. Stabilization of Live Attenuated Influenza Vaccines by Freeze Drying, Spray Drying, and Foam Drying. Pharm. Res. 2016, 33, 1144–1160. [Google Scholar] [CrossRef]
- Ramakrishna, S. An Introduction to Electrospinning and Nanofibers; World Scientific Publishing Co: Singapore, 2005. [Google Scholar]
- Ramakrishna, S.; Fujihara, K.; Teo, W.-E.; Yong, T.; Ma, Z.; Ramaseshan, R. Electrospun nanofibers: Solving global issues. Mater. Today 2006, 9, 40–50. [Google Scholar] [CrossRef]
- Rieger, K.; Birch, N.; Schiffman, J. Designing electrospun nanofiber mats to promote wound healing—A review. J. Mater. Chem. B 2013, 1, 4531–4541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agrahari, V.; Agrahari, V.; Meng, J.; Mitra, A.K. Chapter 9—Electrospun Nanofibers in Drug Delivery: Fabrication, Advances, and Biomedical Applications. In Emerging Nanotechnologies for Diagnostics, Drug Delivery and Medical Devices; Elsevier: Boston, MA, USA, 2017; pp. 189–215. [Google Scholar]
- Hu, X.; Liu, S.; Zhou, G.; Huang, Y.; Xie, Z.; Jing, X. Electrospinning of polymeric nanofibers for drug delivery applications. J. Control. Release 2014, 185, 12–21. [Google Scholar] [CrossRef]
- Xue, J.; Wu, T.; Dai, Y.; Xia, Y. Electrospinning and Electrospun Nanofibers: Methods, Materials, and Applications. Chem. Rev. 2019, 119, 5298–5415. [Google Scholar] [CrossRef]
- Tipduangta, P.; Belton, P.; Fábián, L.; Wang, L.Y.; Tang, H.; Eddleston, M.; Qi, S. Electrospun Polymer Blend Nanofibers for Tunable Drug Delivery: The Role of Transformative Phase Separation on Controlling the Release Rate. Mol. Pharm. 2016, 13, 25–39. [Google Scholar] [CrossRef]
- Babitha, S.; Rachita, L.; Karthikeyan, K.; Shoba, E.; Janani, I.; Poornima, B.; Sai, K.P. Electrospun protein nanofibers in healthcare: A review. Int. J. Pharm. 2017, 523, 52–90. [Google Scholar] [CrossRef]
- Kadajji, V.G.; Betageri, G.V. Water Soluble Polymers for Pharmaceutical Applications. Polymers 2011, 3, 1972–2009. [Google Scholar] [CrossRef] [Green Version]
- Dai, M.; Jin, S.; Nugen, S.R. Water-soluble electrospun nanofibers as a method for on-chip reagent storage. Biosensors 2012, 2, 388–395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gökmeşe, F.; Uslu, I.; Aytimur, A. Preparation and Characterization of PVA/PVP Nanofibers as Promising Materials for Wound Dressing. Polym. Plast. Technol. Eng. 2013, 52, 1259–1265. [Google Scholar] [CrossRef]
- Kamble, R.N.; Gaikwad, S.; Maske, A.; Patil, S.S. Fabrication of electrospun nanofibres of BCS II drug for enhanced dissolution and permeation across skin. J. Adv. Res. 2016, 7, 483–489. [Google Scholar] [CrossRef] [Green Version]
- Sriyanti, I.; Edikresnha, D.; Rahma, A.; Munir, M.M.; Rachmawati, H.; Khairurrijal, K. Mangosteen pericarp extract embedded in electrospun PVP nanofiber mats: Physicochemical properties and release mechanism of α-mangostin. Int. J. Nanomed. 2018, 13, 4927–4941. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Zhao, X.; Tian, Y.; Wang, Y.; Jan, A.K.; Chen, Y. Facile Electrospinning Synthesis of Carbonized Polyvinylpyrrolidone (PVP)/g-C3 N4 Hybrid Films for Photoelectrochemical Applications. Chem. Eur. J. 2017, 23, 419–426. [Google Scholar] [CrossRef] [PubMed]
- Dai, M.; Senecal, A.; Nugen, S.R. Electrospun water-soluble polymer nanofibers for the dehydration and storage of sensitive reagents. Nanotechnology 2014, 25, 225101. [Google Scholar] [CrossRef]
- Kramer, K.; Al-Barwani, F.; Baird, M.A.; Young, V.L.; Larsen, D.S.; Ward, V.K.; Young, S.L. Functionalisation of Virus-Like Particles Enhances Antitumour Immune Responses. J. Immunol. Res. 2019, 2019, 5364632. [Google Scholar] [CrossRef]
- Liu, G.; Ying, H.; Zeng, G.; Wheeler, C.J.; Black, K.L.; Yu, J.S. HER-2, gp100, and MAGE-1 are expressed in human glioblastoma and recognized by cytotoxic T cells. Cancer Res. 2004, 64, 4980–4986. [Google Scholar] [CrossRef] [Green Version]
- Patel, P.M.; Ottensmeier, C.H.; Mulatero, C.; Lorigan, P.; Plummer, R.; Pandha, H.; Elsheikh, S.; Hadjimichael, E.; Villasanti, N.; Adams, S.E.; et al. Targeting gp100 and TRP-2 with a DNA vaccine: Incorporating T cell epitopes with a human IgG1 antibody induces potent T cell responses that are associated with favourable clinical outcome in a phase I/II trial. Oncoimmunology 2018, 7, e1433516. [Google Scholar] [CrossRef] [Green Version]
- Donaldson, B.; Al-Barwani, F.; Pelham, S.J.; Young, K.; Ward, V.K.; Young, S.L. Multi-target chimaeric VLP as a therapeutic vaccine in a model of colorectal cancer. J. Immunotherap. Cancer 2017, 5, 69. [Google Scholar] [CrossRef] [Green Version]
- Peacey, M.; Wilson, S.; Perret, R.; Ronchese, F.; Ward, V.K.; Young, V.; Young, S.L.; Baird, M.A. Virus-like particles from rabbit hemorrhagic disease virus can induce an anti-tumor response. Vaccine 2008, 26, 5334–5337. [Google Scholar] [CrossRef] [PubMed]
- Peacey, M.; Wilson, S.; Baird, M.A.; Ward, V.K. Versatile RHDV virus-like particles: Incorporation of antigens by genetic modification and chemical conjugation. Biotechnol. Bioeng. 2007, 98, 968–977. [Google Scholar] [CrossRef] [PubMed]
- McKee, S.J.; Young, V.L.; Clow, F.; Hayman, C.M.; Baird, M.A.; Hermans, I.F.; Young, S.L.; Ward, V.K. Virus-like particles and alpha-galactosylceramide form a self-adjuvanting composite particle that elicits anti-tumor responses. J. Control. Release 2012, 159, 338–345. [Google Scholar] [CrossRef]
- Inaba, K.; Inaba, M.; Romani, N.; Aya, H.; Deguchi, M.; Ikehara, S.; Muramatsu, S.; Steinman, R.M. Generation of large numbers of dendritic cells from mouse bone marrow cultures supplemented with granulocyte/macrophage colony-stimulating factor. J. Exp. Med. 1992, 176, 1693–1702. [Google Scholar] [CrossRef] [PubMed]
- Faure, F.; Mantegazza, A.; Sadaka, C.; Sedlik, C.; Jotereau, F.; Amigorena, S. Long-lasting cross-presentation of tumor antigen in human DC. Eur. J. Immunol. 2009, 39, 380–390. [Google Scholar] [CrossRef] [PubMed]
- Win, S.J.; Ward, V.K.; Dunbar, P.R.; Young, S.L.; Baird, M.A. Cross-presentation of epitopes on virus-like particles via the MHC I receptor recycling pathway. Immunol. Cell Biol. 2011, 89, 681–688. [Google Scholar] [CrossRef]
- Abutaleb, A.; Lolla, D.; Aljuhani, A.; Shin, H.U.; Rajala, J.W.; Chase, G.G. Effects of Surfactants on the Morphology and Properties of Electrospun Polyetherimide Fibers. Fibers 2017, 5, 33. [Google Scholar] [CrossRef] [Green Version]
- Haider, A.; Haider, S.; Kang, I.-K. A comprehensive review summarizing the effect of electrospinning parameters and potential applications of nanofibers in biomedical and biotechnology. Arab. J. Chem. 2018, 11, 1165–1188. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dowlath, S.; Campbell, K.; Al-Barwani, F.; Young, V.L.; Young, S.L.; Walker, G.F.; Ward, V.K. Dry Formulation of Virus-Like Particles in Electrospun Nanofibers. Vaccines 2021, 9, 213. https://doi.org/10.3390/vaccines9030213
Dowlath S, Campbell K, Al-Barwani F, Young VL, Young SL, Walker GF, Ward VK. Dry Formulation of Virus-Like Particles in Electrospun Nanofibers. Vaccines. 2021; 9(3):213. https://doi.org/10.3390/vaccines9030213
Chicago/Turabian StyleDowlath, Sasheen, Katrin Campbell, Farah Al-Barwani, Vivienne L. Young, Sarah L. Young, Greg F. Walker, and Vernon K. Ward. 2021. "Dry Formulation of Virus-Like Particles in Electrospun Nanofibers" Vaccines 9, no. 3: 213. https://doi.org/10.3390/vaccines9030213
APA StyleDowlath, S., Campbell, K., Al-Barwani, F., Young, V. L., Young, S. L., Walker, G. F., & Ward, V. K. (2021). Dry Formulation of Virus-Like Particles in Electrospun Nanofibers. Vaccines, 9(3), 213. https://doi.org/10.3390/vaccines9030213