
Evaluation of Vertebrate-Specific Replication-Defective Zika Virus, a Novel Single-Cycle Arbovirus Vaccine, in a Mouse Model

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Cultures, Virus Stocks, Escherichia coli, Yeast Strains, and Antibodies
2.2. Plaque Assay
2.3. Fluorescent Focus Assay and Immunostaining
2.4. VSRD-ZIKV Vaccination and Challenge on AG129 Mice
2.5. Intracellular Cytokine Staining (ICS)
2.6. IFN-γ Immunoassay
2.7. RNA Extraction and Quantitative Real-Time (RT)-PCR
2.8. Haematoxylin and Eosin (H&E) Staining
2.9. Statistical Analysis
3. Results
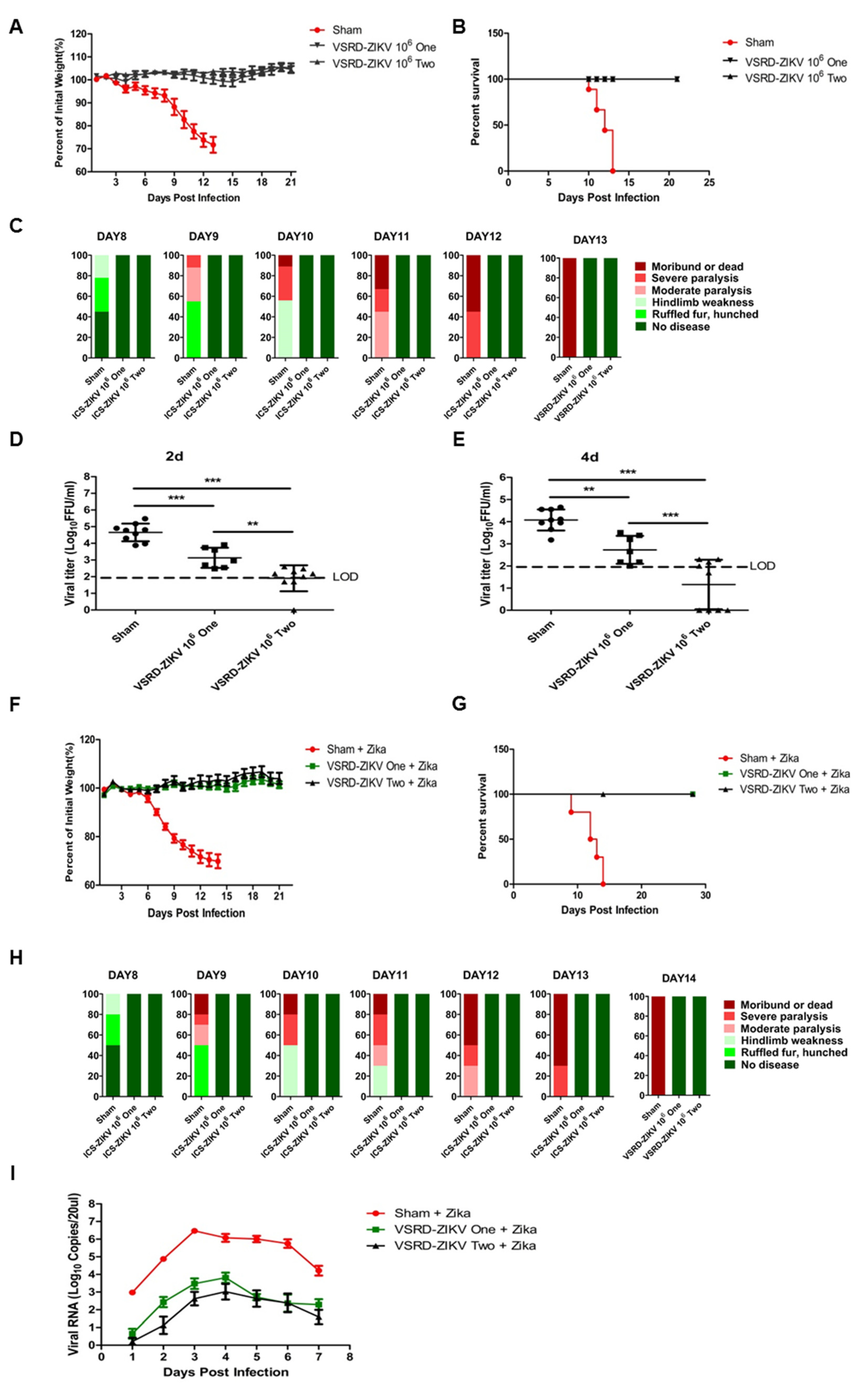
3.1. Prime-Boost VSRD-ZIKV Vaccination Provides Robust Protection Against Lethal ZIKV Challenge in Immunocompromised Mice
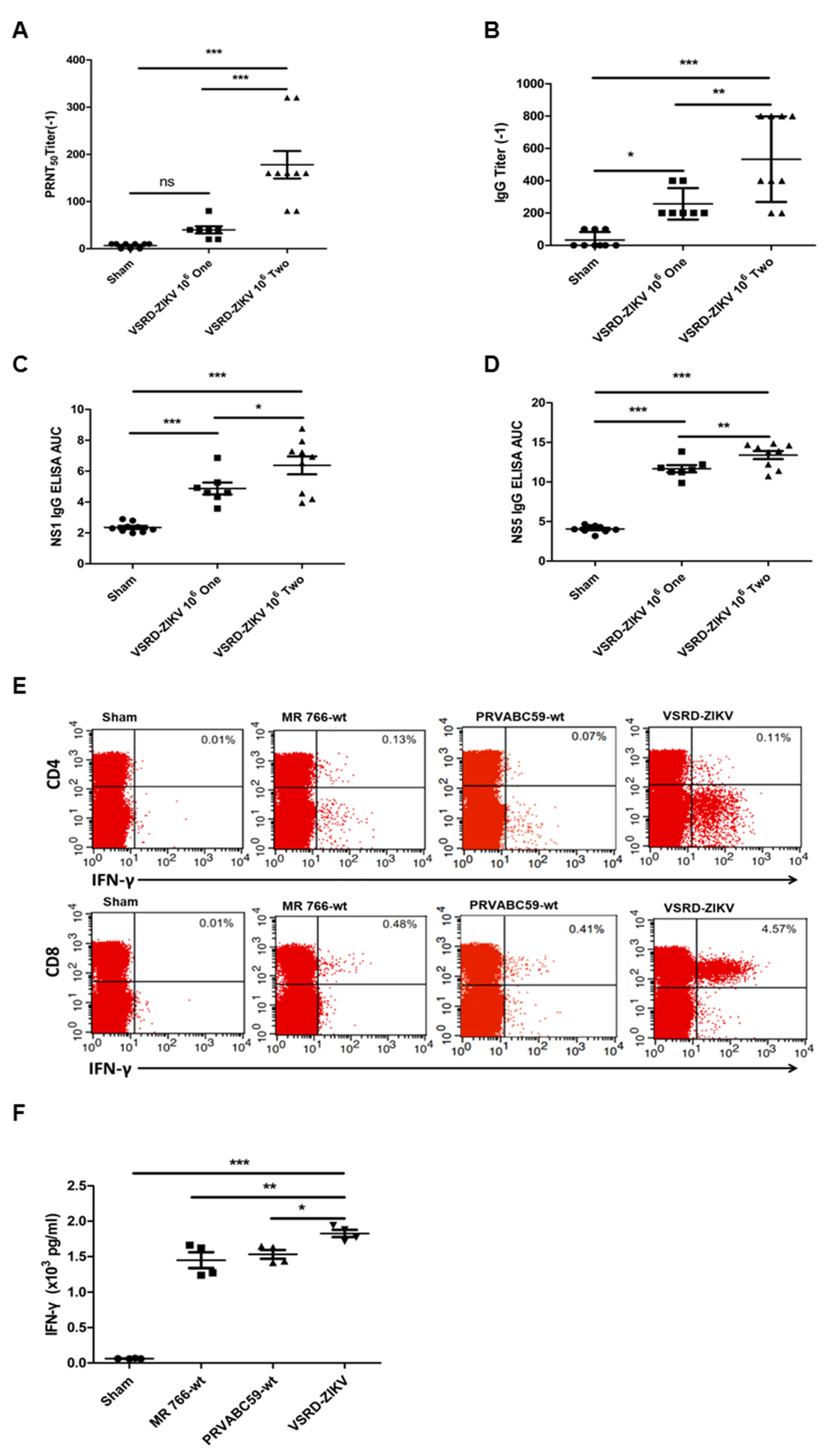
3.2. Vaccination of VSRD-ZIKV is Associated with ZIKV-Specific Humoral and Cellular Immune Responses
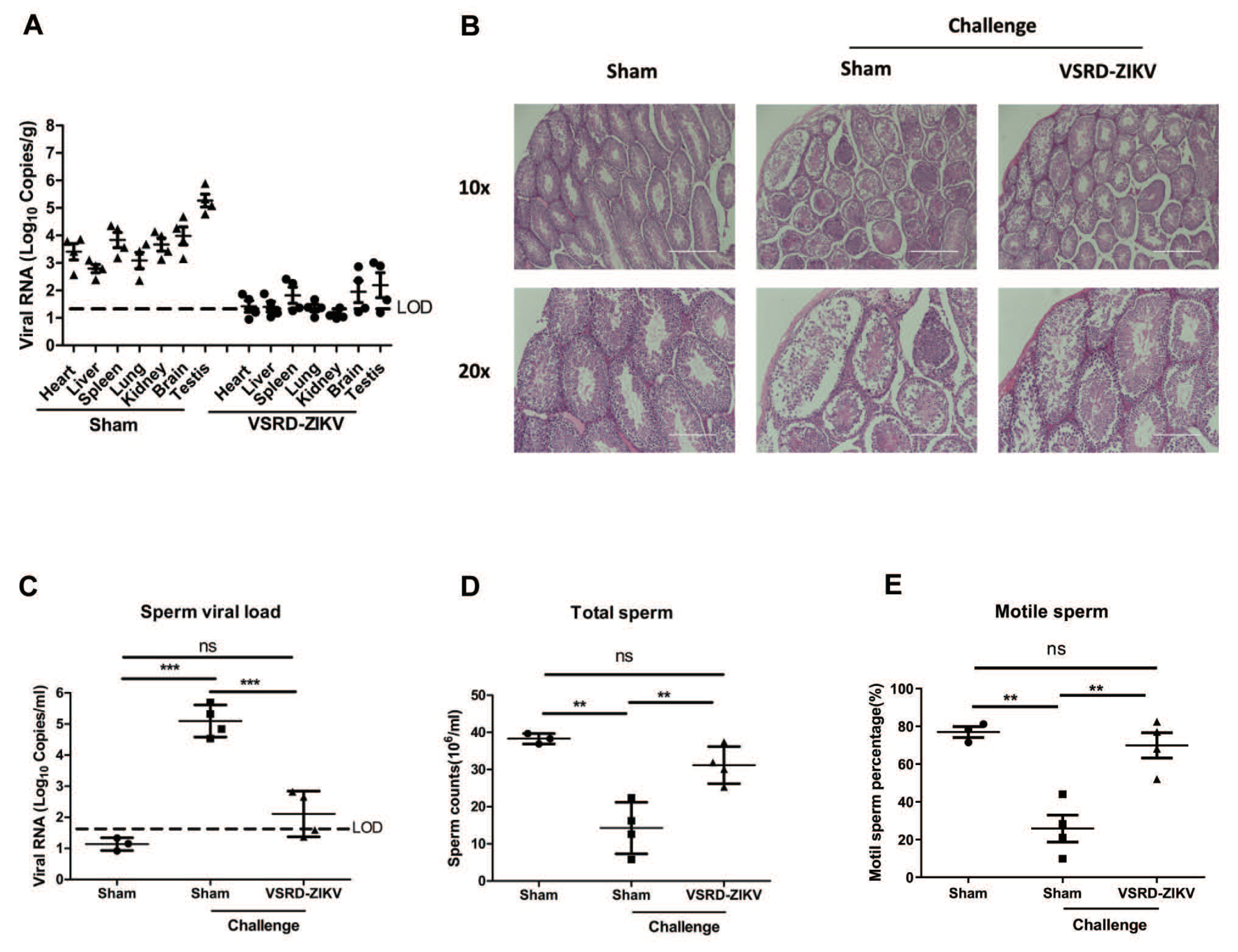
3.3. VSRD-ZIKV Provides Protection and Prevented Virus Accumulation in the Testes of Male Mice Challenged with Lethal ZIKV
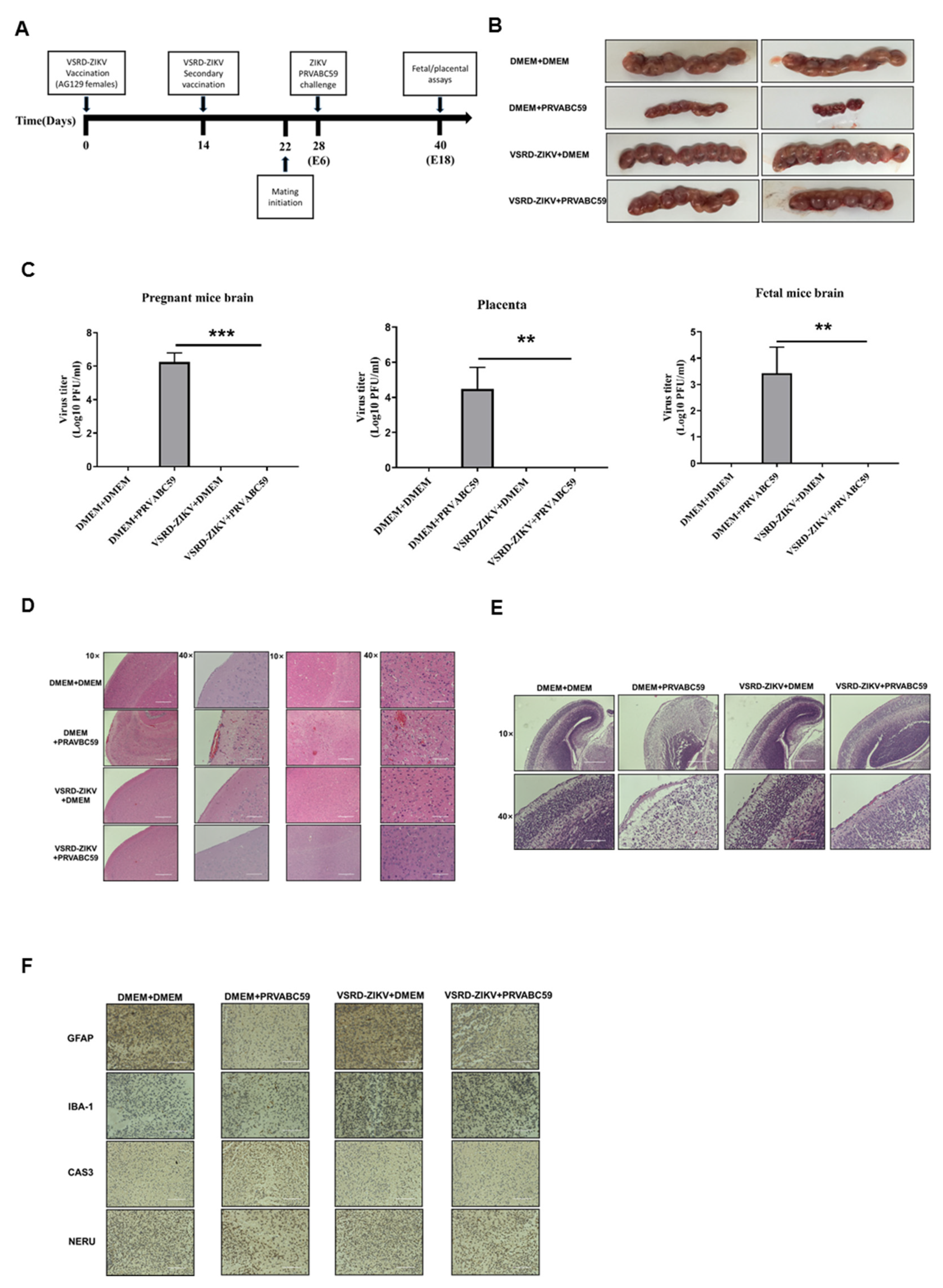
3.4. Immunization with VSRD-ZIKV Protects against Vertical Transmission in Pregnant Mice Challenged with Lethal ZIKV
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Dick, G.W.A.; Kitchen, S.F.; Haddow, A.J. Zika Virus (I). Isolations and serological specificity. Trans. R. Soc. Trop. Med. Hyg. 1952, 46, 509–520. [Google Scholar] [CrossRef]
- Pielnaa, P.; Al-Saadawe, M.; Saro, A.; Dama, M.F.; Zhou, M.; Huang, Y.; Huang, J.; Xia, Z. Zika virus-spread, epidemiology, genome, transmission cycle, clinical manifestation, associated challenges, vaccine and antiviral drug development. Virology 2020, 543, 34–42. [Google Scholar] [CrossRef]
- Gatherer, D.; Kohl, A. Zika virus: A previously slow pandemic spreads rapidly through the Americas. J. Gen. Virol. 2016, 97, 269–273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katz, I.; Gilburd, B.; Shovman, O. Zika autoimmunity and Guillain-Barré syndrome. Curr. Opin. Rheumatol. 2019, 31, 484–487. [Google Scholar] [CrossRef] [PubMed]
- Freitas, D.A.; Souza-Santos, R.; Carvalho, L.M.A.; Barros, W.B.; Neves, L.M.; Brasil, P.; Wakimoto, M.D. Congenital Zika syndrome: A systematic review. PLoS ONE 2020, 15, e0242367. [Google Scholar] [CrossRef]
- Gulland, A. Zika virus is a global public health emergency, declares WHO. BMJ 2016, 352, i657. [Google Scholar] [CrossRef] [Green Version]
- McNeil, C.J.; Shetty, A.K. Zika Virus: A Serious Global Health Threat. J. Trop. Pediatr. 2017, 63, 242–248. [Google Scholar] [CrossRef] [Green Version]
- Pattnaik, A.; Sahoo, B.R.; Pattnaik, A.K. Current Status of Zika Virus Vaccines: Successes and Challenges. Vaccines 2020, 8, 266. [Google Scholar] [CrossRef]
- Xie, X.; Kum, D.B.; Xia, H.; Luo, H.; Shan, C.; Zou, J.; Muruato, A.E.; Medeiros, D.B.; Nunes, B.T.; Dallmeier, K.; et al. A Single-Dose Live-Attenuated Zika Virus Vaccine with Controlled Infection Rounds that Protects against Vertical Transmission. Cell Host Microbe 2018, 24, 487–499.e5. [Google Scholar] [CrossRef] [Green Version]
- Modjarrad, K.; Lin, L.; George, S.L.; E Stephenson, K.; Eckels, K.H.; A De La Barrera, R.; Jarman, R.G.; Sondergaard, E.; Tennant, J.; Ansel, J.L.; et al. Preliminary aggregate safety and immunogenicity results from three trials of a purified inactivated Zika virus vaccine candidate: Phase 1, randomised, double-blind, placebo-controlled clinical trials. Lancet 2018, 391, 563–571. [Google Scholar] [CrossRef] [Green Version]
- Li, X.-F.; Dong, H.-L.; Wang, H.-J.; Huang, X.-Y.; Qiu, Y.-F.; Ji, X.; Ye, Q.; Li, C.; Liu, Y.; Deng, Y.-Q.; et al. Development of a chimeric Zika vaccine using a licensed live-attenuated flavivirus vaccine as backbone. Nat. Commun. 2018, 9, 673. [Google Scholar] [CrossRef] [Green Version]
- Boigard, H.; Alimova, A.; Martin, G.R.; Katz, A.; Gottlieb, P.; Galarza, J.M. Zika virus-like particle (VLP) based vaccine. PLOS Neglected Trop. Dis. 2017, 11, e0005608. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, R.; Liu, Q.; Pang, W.; Gao, F.; Liang, H.; Zhang, W.; Lin, Y.; Li, M.; Liu, Z.; Gao, G.F.; et al. Two immunogenic recombinant protein vaccine candidates showed disparate protective efficacy against Zika virus infection in rhesus macaques. Vaccine 2021, 39, 915–925. [Google Scholar] [CrossRef] [PubMed]
- Richner, J.M.; Jagger, B.W.; Shan, C.; Fontes, C.R.; Dowd, K.A.; Cao, B.; Himansu, S.; Caine, E.A.; Nunes, B.T.; Medeiros, D.B.; et al. Vaccine Mediated Protection against Zika Virus-Induced Congenital Disease. Cell 2017, 170, 273–283.e12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dowd, K.A.; Ko, S.-Y.; Morabito, K.M.; Yang, E.S.; Pelc, R.S.; DeMaso, C.R.; Castilho, L.R.; Abbink, P.; Boyd, M.; Nityanandam, R.; et al. Rapid development of a DNA vaccine for Zika virus. Science 2016, 354, 237–240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, M.; Sun, H.; Lai, H.; Hurtado, J.; Chen, Q. Plant-produced Zika virus envelope protein elicits neutralizing immune responses that correlate with protective immunity against Zika virus in mice. Plant Biotechnol. J. 2018, 16, 572–580. [Google Scholar] [CrossRef] [PubMed]
- Abbink, P.; LaRocca, R.A.; De La Barrera, R.A.; Bricault, C.A.; Moseley, E.T.; Boyd, M.; Kirilova, M.; Li, Z.; Ng’Ang’A, D.; Nanayakkara, O.; et al. Protective efficacy of multiple vaccine platforms against Zika virus challenge in rhesus monkeys. Science 2016, 353, 1129–1132. [Google Scholar] [CrossRef] [Green Version]
- Lambert, P.-H.; Liu, M.; Siegrist, C.-A. Can successful vaccines teach us how to induce efficient protective immune responses? Nat. Med. 2005, 11, S54–S62. [Google Scholar] [CrossRef]
- Amanna, I.J.; Slifka, M.K. Wanted, dead or alive: New viral vaccines. Antivir. Res. 2009, 84, 119–130. [Google Scholar] [CrossRef] [Green Version]
- Widman, D.G.; Frolov, I.; Mason, P.W. Third-generation flavivirus vaccines based on single-cycle, encapsidation-defective viruses. Adv. Virus. Res. 2008, 72, 77–126. [Google Scholar]
- Pang, X.; Guo, Y.; Zhou, Y.; Fu, W.; Gu, X. Highly efficient production of a dengue pseudoinfectious virus. Vaccine 2014, 32, 3854–3860. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mason, P.W.; Shustov, A.V.; Frolov, I. Production and characterization of vaccines based on flaviviruses defective in replication. Virology 2006, 351, 432–443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wan, S.; Cao, S.; Wang, X.; Zhou, Y.; Yan, W.; Gu, X.; Wu, T.-C.; Pang, X. Generation and preliminary characterization of vertebrate-specific replication-defective Zika virus. Virology 2021, 552, 73–82. [Google Scholar] [CrossRef] [PubMed]
- Caine, E.A.; Jagger, B.W.; Diamond, M.S. Animal Models of Zika Virus Infection during Pregnancy. Viruses 2018, 10, 598. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lazear, H.M.; Govero, J.; Smith, A.M.; Platt, D.J.; Fernandez, E.; Miner, J.J.; Diamond, M.S. A Mouse Model of Zika Virus Pathogenesis. Cell Host Microbe 2016, 19, 720–730. [Google Scholar] [CrossRef] [Green Version]
- Hoft, D.F.; Babusis, E.; Worku, S.; Spencer, C.T.; Lottenbach, K.; Truscott, S.M.; Abate, G.; Sakala, I.G.; Edwards, K.M.; Creech, C.B.; et al. Live and Inactivated Influenza Vaccines Induce Similar Humoral Responses, but Only Live Vaccines Induce Diverse T-Cell Responses in Young Children. J. Infect. Dis. 2011, 204, 845–853. [Google Scholar] [CrossRef] [PubMed]
- Wen, J.; Shresta, S. T Cell Immunity to Zika and Dengue Viral Infections. J. Interf. Cytokine Res. 2017, 37, 475–479. [Google Scholar] [CrossRef]
- Morrison, T.E.; Diamond, M.S. Animal Models of Zika Virus Infection, Pathogenesis, and Immunity. J. Virol. 2017, 91, e00009-17. [Google Scholar] [CrossRef] [Green Version]
- Uraki, R.; Hwang, J.; Jurado, K.A.; Householder, S.; Yockey, L.J.; Hastings, A.K.; Homer, R.J.; Iwasaki, A.; Fikrig, E. Zika virus causes testicular atrophy. Sci. Adv. 2017, 3, e1602899. [Google Scholar] [CrossRef] [Green Version]
- Heinz, F.X.; Stiasny, K. Flaviviruses and flavivirus vaccines. Vaccine 2012, 30, 4301–4306. [Google Scholar] [CrossRef]
- LaRocca, R.A.; Abbink, P.; Peron, J.P.S.; Paolo, M.D.A.; Iampietro, M.J.; Badamchi-Zadeh, A.; Boyd, M.; Ng’Ang’A, D.; Kirilova, M.; Nityanandam, R.; et al. Vaccine protection against Zika virus from Brazil. Nature 2016, 536, 474–478. [Google Scholar] [CrossRef] [PubMed]
- Shan, C.; Muruato, A.E.; Jagger, B.W.; Richner, J.; Nunes, B.T.D.; Medeiros, D.B.A.; Xie, X.; Nunes, J.G.C.; Morabito, K.M.; Kong, W.-P.; et al. A single-dose live-attenuated vaccine prevents Zika virus pregnancy transmission and testis damage. Nat. Commun. 2017, 8, 676. [Google Scholar] [CrossRef] [PubMed]
- Morel, P.A.; Falkner, D.; Plowey, J.; Larregina, A.T.; Falo, L.D. DNA immunisation: Altering the cellular localisation of expressed protein and the immunisation route allows manipulation of the immune response. Vaccine 2004, 22, 447–456. [Google Scholar] [CrossRef] [PubMed]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wan, S.; Cao, S.; Wang, X.; Zhou, Y.; Yan, W.; Gu, X.; Wu, T.-C.; Pang, X. Evaluation of Vertebrate-Specific Replication-Defective Zika Virus, a Novel Single-Cycle Arbovirus Vaccine, in a Mouse Model. Vaccines 2021, 9, 338. https://doi.org/10.3390/vaccines9040338
Wan S, Cao S, Wang X, Zhou Y, Yan W, Gu X, Wu T-C, Pang X. Evaluation of Vertebrate-Specific Replication-Defective Zika Virus, a Novel Single-Cycle Arbovirus Vaccine, in a Mouse Model. Vaccines. 2021; 9(4):338. https://doi.org/10.3390/vaccines9040338
Chicago/Turabian StyleWan, Shengfeng, Shengbo Cao, Xugang Wang, Yanfei Zhou, Weidong Yan, Xinbin Gu, Tzyy-Choou Wu, and Xiaowu Pang. 2021. "Evaluation of Vertebrate-Specific Replication-Defective Zika Virus, a Novel Single-Cycle Arbovirus Vaccine, in a Mouse Model" Vaccines 9, no. 4: 338. https://doi.org/10.3390/vaccines9040338
APA StyleWan, S., Cao, S., Wang, X., Zhou, Y., Yan, W., Gu, X., Wu, T. -C., & Pang, X. (2021). Evaluation of Vertebrate-Specific Replication-Defective Zika Virus, a Novel Single-Cycle Arbovirus Vaccine, in a Mouse Model. Vaccines, 9(4), 338. https://doi.org/10.3390/vaccines9040338