
Vaccine Quality Is a Key Factor to Determine Thermal Stability of Commercial Newcastle Disease (ND)Vaccines



Abstract
:1. Introduction
2. Material and Methods
2.1. Determination of Infectivity
2.2. Hemagglutination Assay
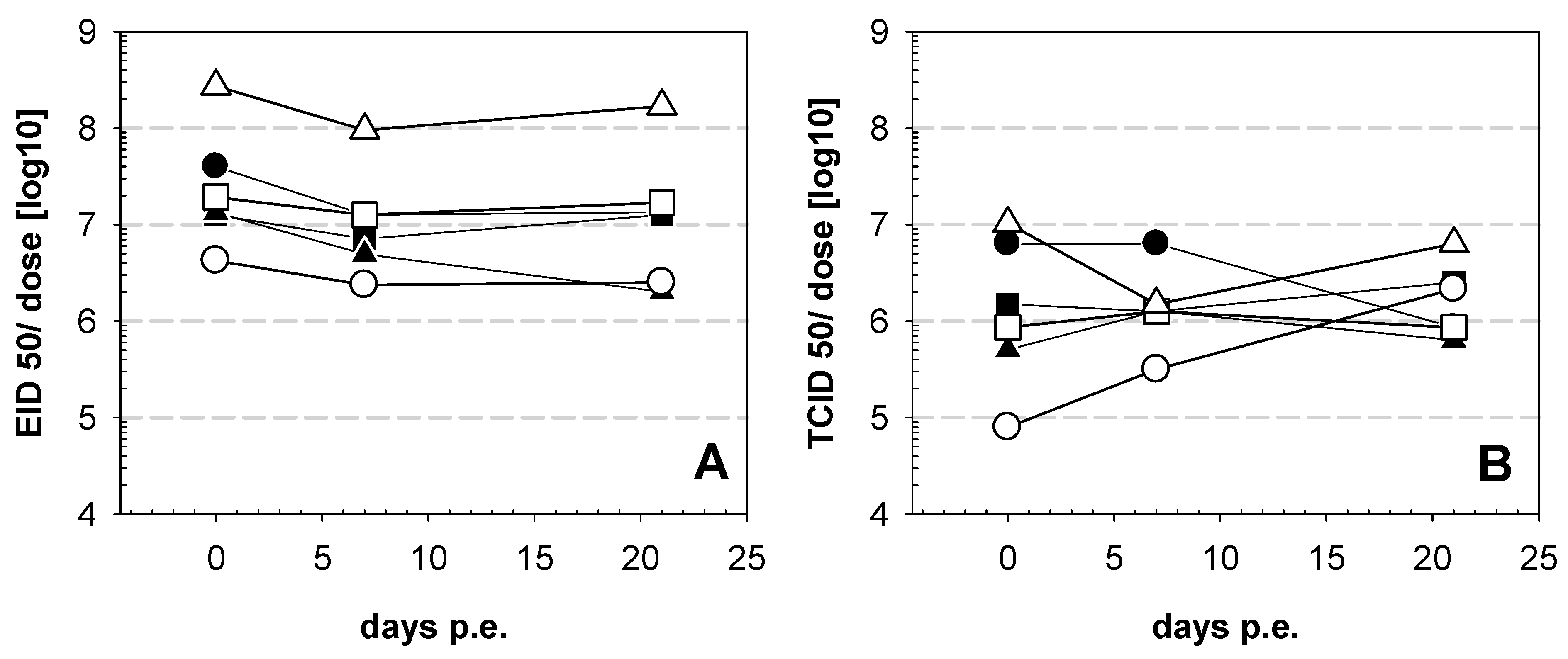
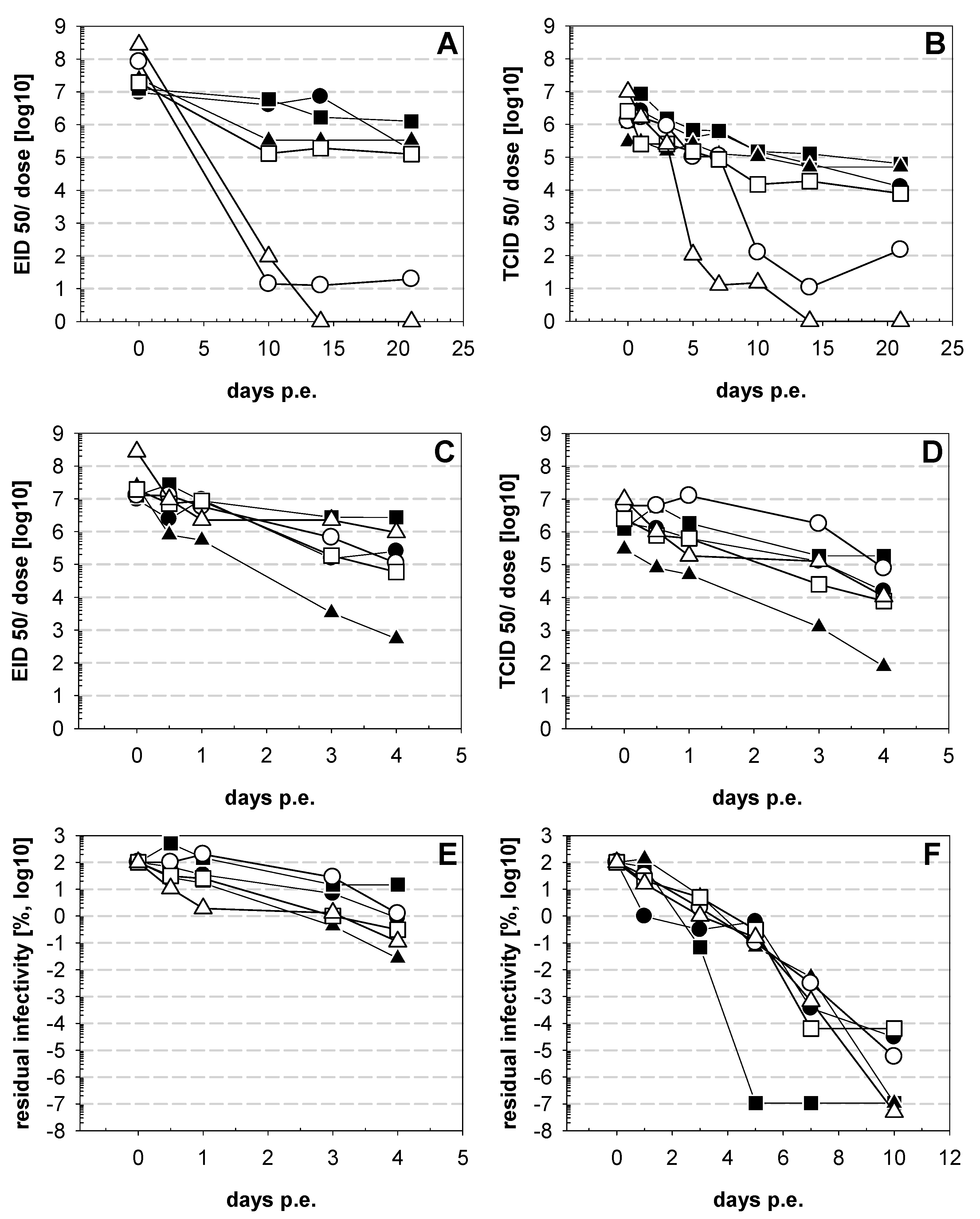
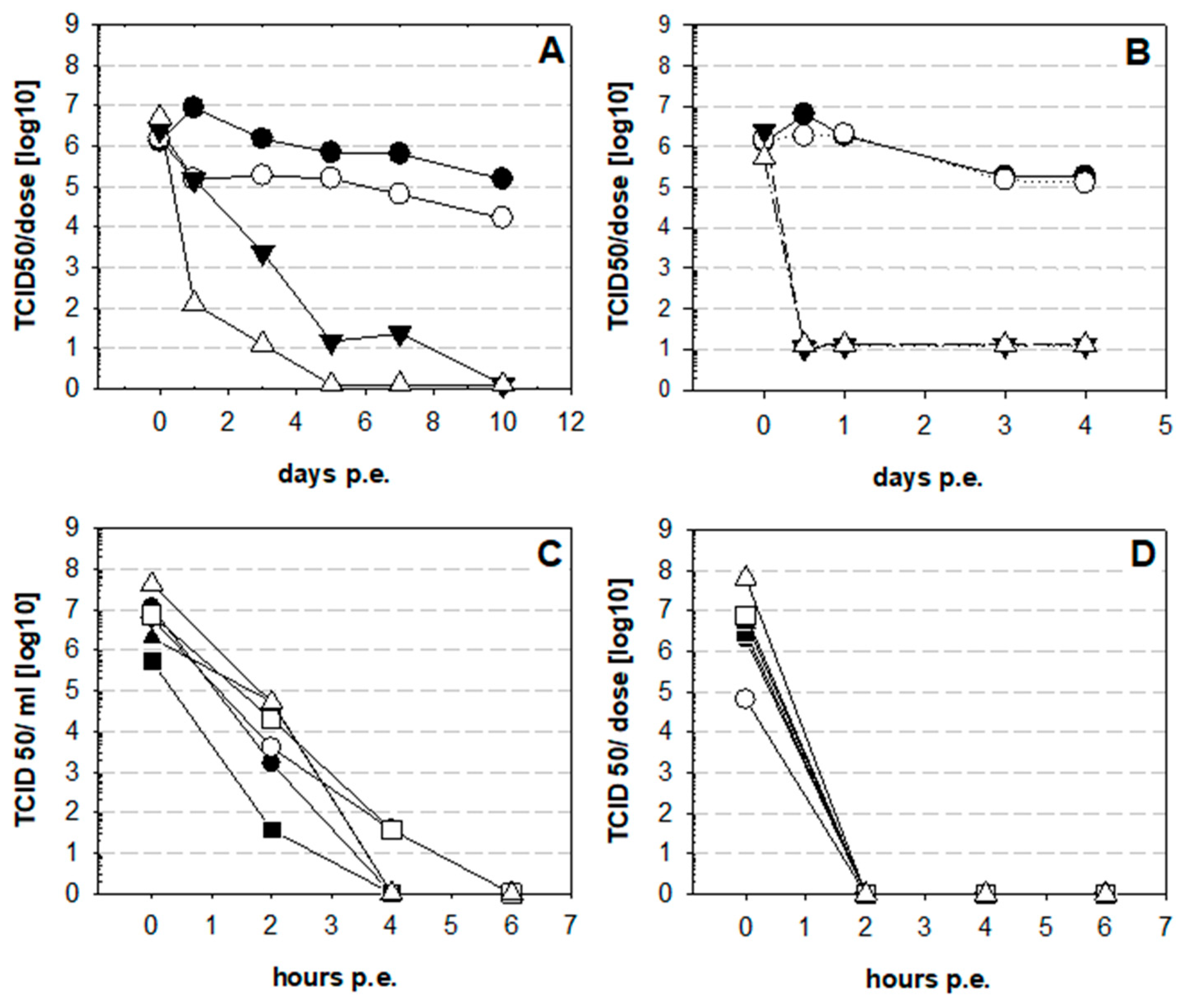
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bagust, T.J. Poultry health and disease control in developing countries. In Poultry Development Review; FAO: Rome, Italy, 2014; ISBN 978-92-5-108067-2. [Google Scholar]
- Ganar, K.; Das, M.; Sinha, S.; Kumar, S. Newcastle disease virus: Current status and our understanding. Virus Res. 2014, 184, 71–81. [Google Scholar] [CrossRef] [PubMed]
- Khatun, M.; Islam, I.; Ershaduzzaman, M.; Islam, H.; Yasmin, S.; Hossen, A.; Hasan, M. Economic Impact of Newcastle Disease on Village Chickens-A Case of Bangladesh. Asian Inst. Res. J. Econ. Bus. 2018, 1, 358–367. [Google Scholar] [CrossRef]
- Shittu, I.; Joannis, T.M.; Odaibo, G.N.; Olaleye, O.D. Newcastle disease in Nigeria: Epizootiology and current knowledge of circulating genotypes. VirusDisease 2016, 27, 329–339. [Google Scholar] [CrossRef] [Green Version]
- Cattoli, G.; Susta, L.; Terregino, C.; Brown, C. Newcastle disease: A review of field recognition and current methods of laboratory detection. J. Vet. Diagn. Investig. 2011, 23, 637–656. [Google Scholar] [CrossRef] [Green Version]
- ICTV International Committee on Taxonomy of Viruses. Virus Taxonomy. 2019. Available online: https://talk.ictvonline.org/taxonomy/ (accessed on 19 November 2020).
- Samal, S.K. Newcastle disease and related avian paramyxoviruses. In The Biology of Paramyxoviruses; Samal, S.K., Ed.; Caister Academic Press: Norfolk, UK, 2011; pp. 69–114. [Google Scholar]
- Goldhaft, T.M. Historical note on the origin of the LaSota strain of Newcastle disease virus. Avian Dis. 1980, 24, 297–301. [Google Scholar] [CrossRef] [Green Version]
- Hitchner, S.B. Serendipity in science. Discovery of the B 1 strain of Newcastle disease virus. Avian Dis. 1975, 19, 215–223. [Google Scholar] [CrossRef]
- Simmons, G.C. The isolation of Newcastle disease virus in Queensland. Aust. Vet. J. 1967, 43, 29–30. [Google Scholar] [CrossRef]
- Alders, R.G.; Bagnol, B.; Young, M.P. Technically sound and sustainable Newcastle disease control in village chickens: Lessons learnt over fifteen years. Worlds Poult. Sci. J. 2010, 66, 433–440. [Google Scholar] [CrossRef]
- Bessell, P.R.; Woolley, R.; Stevenson, S.; Al-Riyami, L.; Opondo, P.; Lai, L.; Gammon, N. An analysis of the impact of Newcastle disease vaccination and husbandry practice on smallholder chicken productivity in Uganda. Prev. Vet. Med. 2020, 177, 104975. [Google Scholar] [CrossRef] [PubMed]
- Aini, I.; Ibrahim, A.L.; Spradbrow, P.B. Field trials of a food-based vaccine to protect village chickens against Newcastle disease. Res. Vet. Sci. 1990, 49, 216–219. [Google Scholar] [CrossRef]
- Allan, W.H.; Lancaster, J.E.; Toth, B. Newcastle Disease Vaccines Their Production and Use; FAO Animal Production Series No. 10; FAO: Rome, Italy, 1978. [Google Scholar]
- Young, M.; Alders, R.; Grimes, S.; Spradbrow, P.; Dias, P.; da Silva, A.; Lobo, Q. Controlling Newcastle Disease in Village Chickens: A Laboratory Manual, 2nd ed.; ACIAR Monograph; ACIAR: Bruce, Australia, 2012; Volume 87, pp. 31–66. [Google Scholar]
- Matthias, D.M.; Robertson, J.; Garrison, M.M.; Newland, S.; Nelson, C. Freezing temperatures in the vaccine cold chain: A systematic literature review. Vaccine 2007, 25. [Google Scholar] [CrossRef] [Green Version]
- Nelson, C.; Froes, P.; Van Dyck, A.M.; Chavarría, J.; Boda, E.; Coca, A.; Crespo, G.; Lima, H. Monitoring temperatures in the vaccine cold chain in Bolivia. Vaccine 2007, 25, 3980–3986. [Google Scholar] [CrossRef]
- Milhomme, P. Cold chain study: Danger of freezing vaccines. Can. Commun. Dis. Rep. 1993, 19, 33–38. [Google Scholar]
- Setia, S.; Mainzer, H.; Washington, M.L.; Coil, G.; Snyder, R.; Weniger, B.G. Frequency and causes of vaccine wastage. Vaccine 2002, 20, 1148–1156. [Google Scholar] [CrossRef]
- Schlehuber, L.D.; McFadyen, I.J.; Shu, Y.; Carignan, J.; Duprex, W.P.; Forsyth, W.R.; Ho, J.H.; Kitsos, C.M.; Lee, G.Y.; Levinson, D.A.; et al. Towards ambient temperature-stable vaccines: The identification of thermally stabilizing liquid formulations for measles virus using an innovative high-throughput infectivity assay. Vaccine 2011, 29, 5031–5039. [Google Scholar] [CrossRef] [Green Version]
- Wen, G.; Shang, Y.; Guo, J.; Chen, C.; Shao, H.; Luo, Q.; Yang, J.; Wang, H.; Cheng, G. Complete genome sequence and molecular characterization of thermostable newcastle disease virus strain Ts09-C. Virus Genes 2013, 46, 542–545. [Google Scholar] [CrossRef]
- Bensink, Z.; Spradbrow, P. Newcastle disease virus strain I2—A prospective thermostable vaccine for use in developing countries. Vet. Microbiol. 1999, 68, 131–139. [Google Scholar] [CrossRef]
- Wen, G.; Shang, Y.; Guo, J.; Yang, J.; Wang, H.; Luo, Q.; Zahng, R.; Shao, H.B. Serial passages culture of Newcastle disease virus thermostable TS09-C strain in BHK cells. Hubei Agric. Cult. Sci. 2012, 51, 5424–5427. [Google Scholar]
- Mazija, H.; Cajavec, S.; Ergotić, N.; Ciglar-Grozdanić, I.; Gottstein, Z.; Ragland, W.L. Immunogenicity and safety of Queensland V4 and Ulster 2C strains of Newcastle disease virus given to maternally immune, newly hatched chickens by nebulization. Avian Dis. 2010, 54, 99–103. [Google Scholar] [CrossRef] [PubMed]
- Wambura, P.N.; Kataga, S. Putative protective antibody response following oral vaccination of multi-age free ranging helmeted guinea fowls (Numida meleagris) with Newcastle disease virus strain I-2 coated on oiled rice. Trop. Anim. Health Prod. 2011, 43, 99–102. [Google Scholar] [CrossRef] [PubMed]
- Wen, G.; Chen, C.; Guo, J.; Zhang, Z.; Shang, Y.; Shao, H.; Luo, Q.; Yang, J.; Wang, H.; Wang, H.; et al. Development of a novel thermostable Newcastle disease virus vaccine vector for expression of a heterologous gene. J. Gen. Virol. 2015, 96, 1219–1228. [Google Scholar] [CrossRef]
- Wen, G.; Hu, X.; Zhao, K.; Wang, H.; Zhang, Z.; Zhang, T.; Yang, J.; Luo, Q.; Zhang, R.; Pan, Z.; et al. Molecular basis for the thermostability of Newcastle disease virus. Sci. Rep. 2016, 3, 22492. [Google Scholar] [CrossRef] [Green Version]
- Kawaguchi, T.; Nomura, K.; Hirayama, Y.; Kitagawa, T. Establishment and Characterization of a Chicken Hepatocellular Carcinoma Cell Line, LMH. Cancer Res. 1987, 47, 4460–4463. [Google Scholar]
- Office International des Epizooties (OIE). Newcastle disease. In Manual of Diagnostic Tests and Vaccines for Terrestrial Animals, 6th ed.; OIE: Paris, France, 2008; Volume 1, pp. 576–589. [Google Scholar]
- Reed, L.J.; Muench, H. A simple method of estimating fifty per cent endpoints. Am. J. Epidemiol. 1938, 27, 493–497. [Google Scholar] [CrossRef]
- Boumart, Z.; Hamdi, J.; Daouam, S.; Elarkam, A.; Tadlaoui, K.O.; El Harrak, M. Thermal Stability Study of Five Newcastle Disease Attenuated Vaccine Strains. Avian Dis. 2016, 60, 779–783. [Google Scholar] [CrossRef] [PubMed]
- Wood, G.W.; Hebert, C.N.; Thornton, D.H. The stability of live Newcastle disease vaccines. J. Biol. Stand. 1980, 8, 157–163. [Google Scholar] [CrossRef]
- Pisal, S.; Wawde, G.; Salvankar, S.; Lade, S.; Kadam, S. Vacuum foam drying for preservation of LaSota virus: Effect of additives. AAPS PharmSciTech 2006, 7, E30–E37. [Google Scholar] [CrossRef]
- Kumru, O.S.; Joshi, S.B.; Smith, D.E.; Middaugh, C.R.; Prusik, T.; Volkin, D.B. Vaccine instability in the cold chain: Mechanisms, analysis and formulation strategies. Biologicals 2014, 42, 237–259. [Google Scholar] [CrossRef] [Green Version]
- Rani, S.; Gogoi, P.; Kumar, S. Spectrum of Newcastle disease virus stability in gradients of temperature and pH. Biologicals 2014, 42, 293–297. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Code | Vaccine | Strain | Dose/Vial | Vol. 1 | EID 2 | TCID 2 | HAU 3 | EID/TCID | EID50/HAU | |
|---|---|---|---|---|---|---|---|---|---|---|
| A | La Sota thermostable ND | Hester | La Sota | 100 | 2 mL | 4.7 × 106 | 9.3 × 105 | 128 | 5.1 | 3.7 × 106 |
| B | ND clone 30 | MSD AH | clone 30 | 1000 | 2 mL | 6.3 × 106 | 6.3 × 106 | 128 | 10.0 | 4.9 × 107 |
| C | Hipraviar CLON | Hipra | CI/79 | 5000 | 2 mL | 1.3 × 107 | 1.3 × 105 | 512 | 84.8 | 1.2 × 108 |
| D | ND-Kukustar | Brentec | I-2 | 500 | 1 mL | 8.3 × 107 | 6.3 × 108 | 256 | 65.8 | 1.6 × 108 |
| E | Avivax ND I-2 lyophilisé | MCI Santé Animale | I-2 | 100 | 2 mL | 9.6 × 106 | 1.3 × 106 | 512 | 7.6 | 1.9 × 106 |
| F | Avivax I-2 | Kevevapi | I-2 | 100 | 2 mL | 1.4 × 108 | 4.9 × 106 | 64 | 28 | 2.1 × 106 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Osman, N.; Goovaerts, D.; Sultan, S.; Salt, J.; Grund, C. Vaccine Quality Is a Key Factor to Determine Thermal Stability of Commercial Newcastle Disease (ND)Vaccines. Vaccines 2021, 9, 363. https://doi.org/10.3390/vaccines9040363
Osman N, Goovaerts D, Sultan S, Salt J, Grund C. Vaccine Quality Is a Key Factor to Determine Thermal Stability of Commercial Newcastle Disease (ND)Vaccines. Vaccines. 2021; 9(4):363. https://doi.org/10.3390/vaccines9040363
Chicago/Turabian StyleOsman, Nabila, Danny Goovaerts, Serageldeen Sultan, Jeremy Salt, and Christian Grund. 2021. "Vaccine Quality Is a Key Factor to Determine Thermal Stability of Commercial Newcastle Disease (ND)Vaccines" Vaccines 9, no. 4: 363. https://doi.org/10.3390/vaccines9040363
APA StyleOsman, N., Goovaerts, D., Sultan, S., Salt, J., & Grund, C. (2021). Vaccine Quality Is a Key Factor to Determine Thermal Stability of Commercial Newcastle Disease (ND)Vaccines. Vaccines, 9(4), 363. https://doi.org/10.3390/vaccines9040363