
The Long-Term Impact of Rotavirus Vaccines in Korea, 2008–2020; Emergence of G8P[8] Strain


Abstract
:1. Introduction
2. Materials and Methods
3. Results
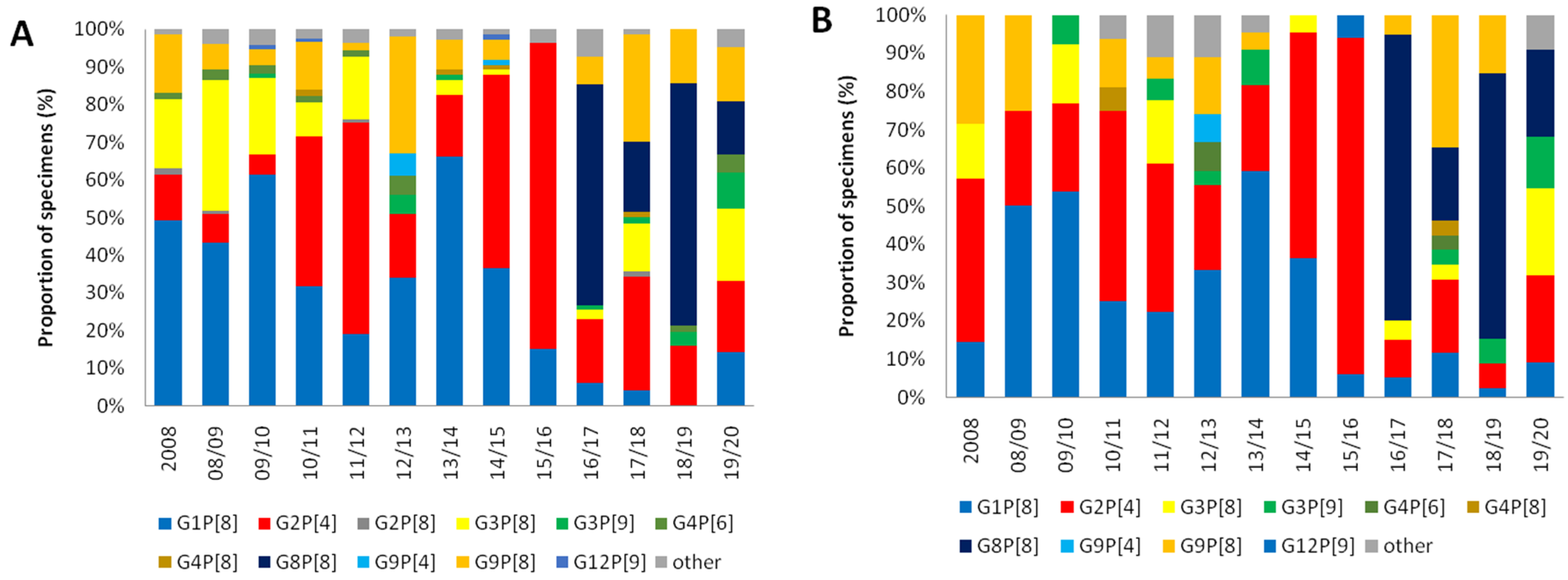
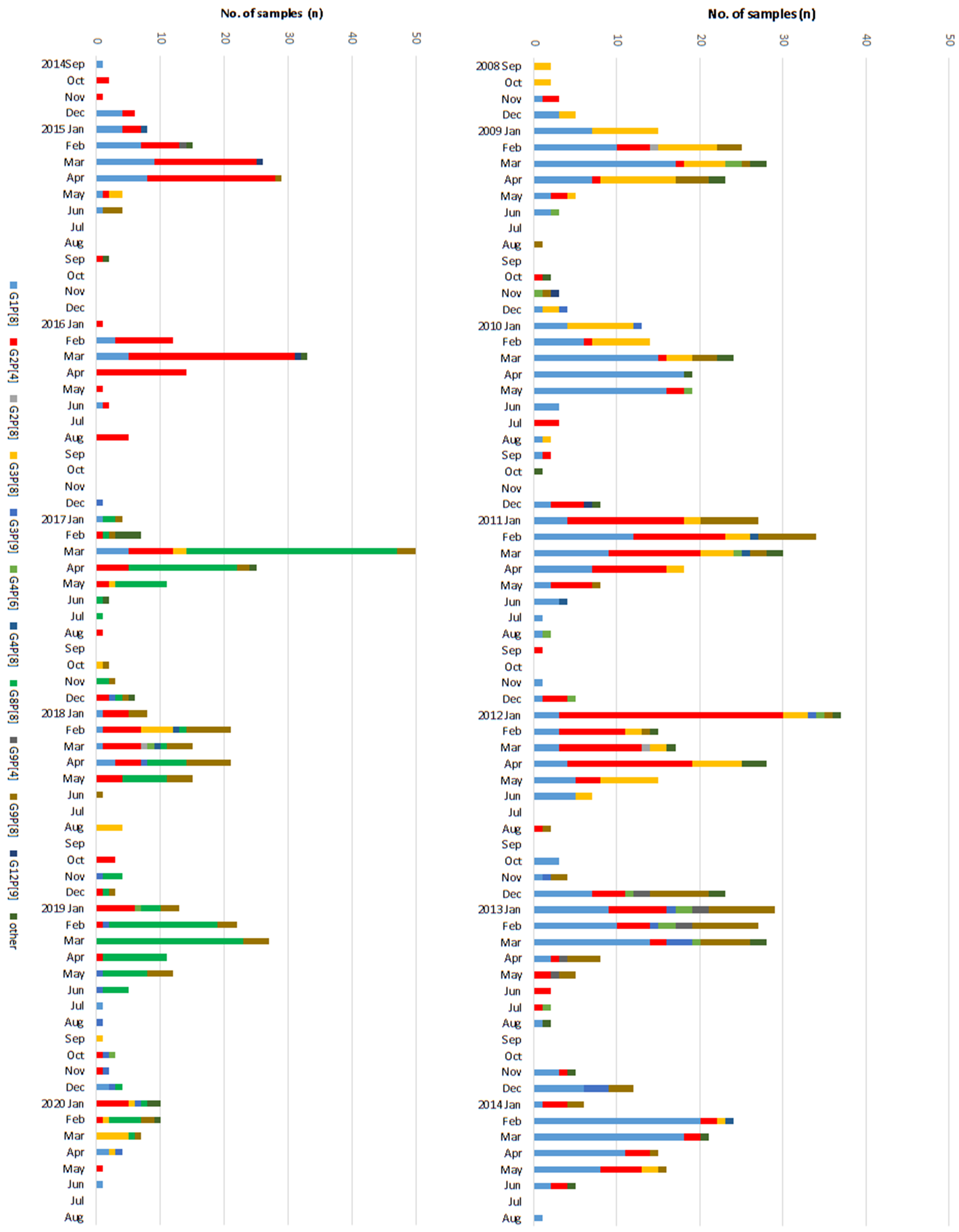
3.1. Changes in Distribution of the RV Genotypes Following Vaccine Introduction
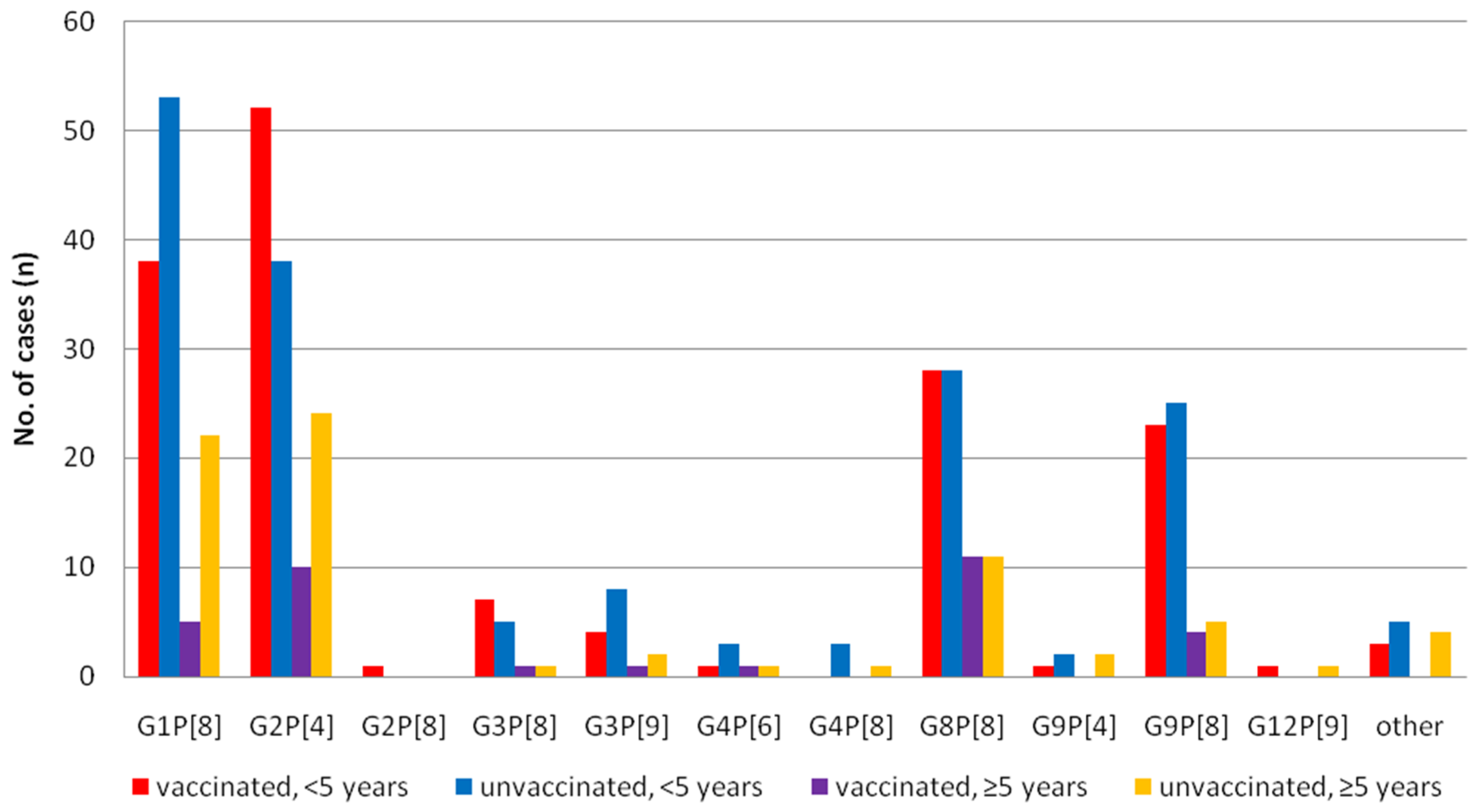
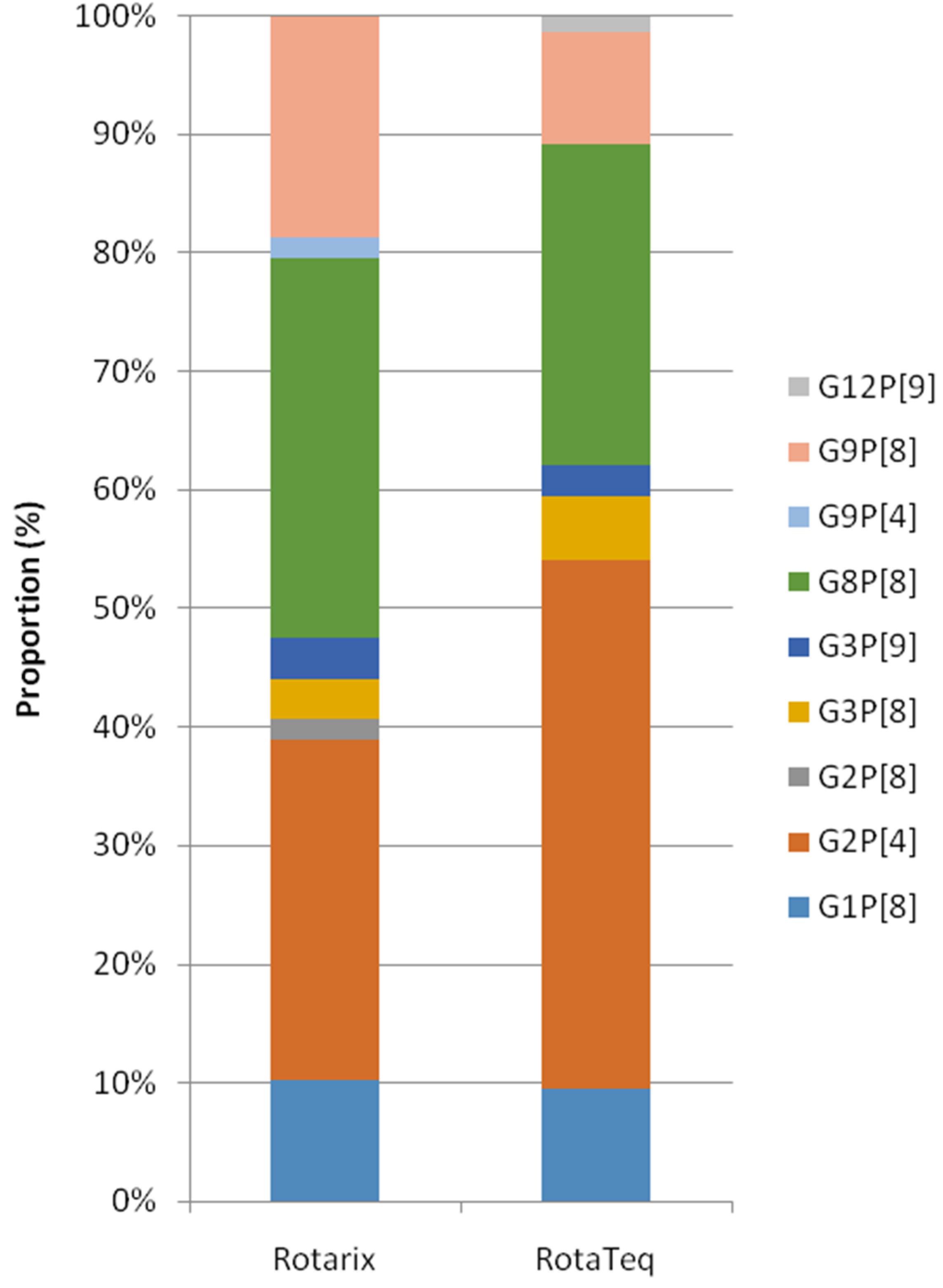
3.2. Differences of the RV Genotypes by Vaccine Status
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dhama, K.; Chauhan, R.S.; Mahendran, M.; Malik, S.V.S. Rotavirus diarrhea in bovines and other domestic animals. Vet. Res. Commun. 2009, 33, 1–23. [Google Scholar] [CrossRef]
- Gozalbo-Rovira, R.; Rubio-del-Campo, A.; Santiso-Bellón, C.; Vila-Vicent, S.; Buesa, J.; Delgado, S.; Molinero, N.; Margolles, A.; Yebra, M.J.; Collado, M.C.; et al. Interaction of intestinal bacteria with human rotavirus during infection in children. Int. J. Mol. Sci. 2021, 22, 1010. [Google Scholar] [CrossRef]
- Bibera, G.L.; Chen, J.; Pereira, P.; Benninghoff, B. Dynamics of G2P [4] strain evolution and rotavirus vaccination: A review of evidence for Rotarix. Vaccine 2020, 38, 5591–5600. [Google Scholar] [CrossRef]
- Crawford, S.E.; Ramani, S.; Tate, J.E.; Parashar, U.D.; Svensson, L.; Hagbom, M.; Franco, M.A.; Greenberg, H.B.; O’Ryan, M.; Kang, G.; et al. Rotavirus infection. Nat. Rev. Dis. Prim. 2017, 3, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.C.; Tan, L.B.; Huang, L.M.; Chen, K.T. Rotavirus infection and the current status of rotavirus vaccines. J. Formos. Med. Assoc. 2012, 111, 183–193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cho, H.; Lee, H.; Kim, D.S.; Kim, H.M.; Kim, J.H.; Kim, A.-Y.; Kang, H.-Y. Socioeconomic Impact of the Rotavirus Vaccine in Korea: Comparing the Epidemiologic and Economic Characteristics of Rotavirus Gastroenteritis before and after the Introduction of Vaccines. Pediatr. Infect. Dis. J. 2020, 39, 460–465. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.G.; Jeon, S.Y.; Park, K.S. Rotavirus vaccine coverage and related factors. J. Korean Soc. Matern. Child. Health 2019, 23, 175–184. [Google Scholar] [CrossRef]
- Burnett, E.; Parashar, U.D.; Tate, J.E. Real-world effectiveness of rotavirus vaccines, 2006–19: A literature review and meta-analysis. Lancet Glob. Health 2020, 8, e1195–e1202. [Google Scholar] [CrossRef]
- Burnett, E.; Jonesteller, C.L.; Tate, J.E.; Yen, C.; Parashar, U.D. Global impact of rotavirus vaccination on childhood hospitalizations and mortality from diarrhea. J. Infect. Dis. 2017, 215, 1666–1672. [Google Scholar] [CrossRef] [PubMed]
- Troeger, C.; Khalil, I.A.; Rao, P.C.; Cao, S.; Blacker, B.F.; Ahmed, T.; Armah, H.; Bines, J.E.; Brewer, T.G.; Colombara, D.V.; et al. Rotavirus vaccination and the global burden of rotavirus diarrhea among children younger than 5 years. JAMA Pediatr. 2018, 172, 958–965. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karafillakis, E.; Hassounah, S.; Atchison, C. Effectiveness and impact of rotavirus vaccines in Europe, 2006–2014. Vaccine 2015, 33, 2097–2107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Markkula, J.; Hemming-Harlo, M.; Savolainen-Kopra, C.; Al-Hello, H.; Vesikari, T. Continuing rotavirus circulation in children and adults despite high coverage rotavirus vaccination in Finland. J. Infect. 2020, 80, 76–83. [Google Scholar] [CrossRef] [Green Version]
- Zlamy, M.; Kofler, S.; Orth, D.; Wüzner, R.; Heinz-Erian, P.; Streng, A.; Prelog, M. The impact of Rotavirus mass vaccination on hospitalization rates, nosocomial Rotavirus gastroenteritis and secondary blood stream infections. BMC Infect. Dis. 2013, 13, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gil-Prieto, R.; Gonzalez-Escalada, A.; Alvaro-Meca, A.; Garcia-Garcia, L.; San-Martin, M.; González-López, A.; Gil-de-Miguel, A. Impact of non-routine rotavirus vaccination on hospitalizations for diarrhoea and rotavirus infections in Spain. Vaccine 2013, 31, 5000–5004. [Google Scholar] [CrossRef]
- Leshem, E.; Tate, J.E.; Steiner, C.A.; Curns, A.T.; Lopman, B.A.; Parashar, U.D. National estimates of reductions in acute gastroenteritis–related hospitalizations and associated costs in US children after implementation of rotavirus vaccines. J. Pediatr. Infect. Dis.Soc. 2018, 7, 257–260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suzuki, H.; Sakai, T.; Tanabe, N.; Okabe, N. Peak rotavirus activity shifted from winter to early spring in Japan. Pediatr. Infect. Dis. J. 2005, 24, 257–260. [Google Scholar] [CrossRef]
- Tate, J.E.; Panozzo, C.A.; Payne, D.C. Decline and change in seasonality of US rotavirus activity after the introduction of rotavirus vaccine. Pediatrics 2009, 124, 465–471. [Google Scholar] [CrossRef]
- Aliabadi, N.; Tate, J.E.; Haynes, A.K.; Parashar, U.D. Sustained decrease in laboratory detection of rotavirus after implementation of routine vaccination—United States, 2000–2014. MMWR 2015, 64, 337. [Google Scholar]
- Sadiq, A.; Bostan, N.; Bokhari, H. Molecular characterization of human group A rotavirus genotypes circulating in Rawalpindi, Islamabad, Pakistan during 2015–2016. PLoS ONE 2019, 14, e0220387. [Google Scholar] [CrossRef] [Green Version]
- Pitzer, V.E.; Bilcke, J.; Heylen, E. Did large-scale vaccination drive changes in the circulating rotavirus population in Belgium? Sci. Rep. 2015, 5, 1–14. [Google Scholar]
- Bowen, M.D.; Mijatovic-Rustempasic, S.; Esona, M.D. Rotavirus strain trends during the postlicensure vaccine era: United States, 2008–2013. J. Infect. Dis. 2016, 214, 732–738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dóró, R.; László, B.; Martella, V.; Leshem, E.; Gentsch, J.; Parashar, U.; Bányai, K. Review of global rotavirus strain prevalence data from six years post vaccine licensure surveillance: Is there evidence of strain selection from vaccine pressure? Infect. Genet. Evol. 2014, 28, 446–461. [Google Scholar] [CrossRef]
- Lee, S.Y.; Hong, S.K.; Lee, S.G.; Suh, C.I.; Park, S.W.; Lee, J.H.; Kim, J.H.; Jang, Y.T.; Ma, S.H.; Kim, S.Y.; et al. Human rotavirus genotypes in hospitalized children, South Korea, April 2005 to March 2007. Vaccine 2009, 27, F97–F101. [Google Scholar] [CrossRef] [PubMed]
- Le, V.P.; Kim, J.Y.; Cho, S.L.; Nam, S.-W.; Lim, I.; Lee, H.-J.; Kim, K.; Chung, S.-I.; Song, W.; Lee, K.M.; et al. Detection of unusual rotavirus genotypes G8P[8] and G12P[6] in South Korea. J. Med. Virol. 2008, 80, 175–182. [Google Scholar] [CrossRef] [PubMed]
- Han, T.H.; Kim, C.H.; Chung, J.Y.; Park, S.H.; Hwang, E.S. Genetic characterization of rotavirus in children in South Korea from 2007 to 2009. Arch. Virol. 2010, 155, 1663–1673. [Google Scholar] [CrossRef] [PubMed]
- Thanh, H.D.; Tran, V.T.; Lim, I.; Kim, W. Emergence of human G2P [4] rotaviruses in the post-vaccination era in South Korea: Footprints of multiple interspecies re-assortment events. Sci. Rep. 2018, 8, 1–10. [Google Scholar]
- Zeller, M.; Rahman, M.; Heylen, E.; De Coster, S.; De Vos, S.; Arijs, I.; Novo, L.; Verstappen, N.; Van Ranst, M.; Matthijnssens, J. Rotavirus incidence and genotype distribution before and after national rotavirus vaccine introduction in Belgium. Vaccine 2010, 28, 7507–7513. [Google Scholar] [CrossRef]
- Hoque, S.A.; Khandoker, N.; Thongprachum, A.; Khamrin, P.; Takanshi, S.; Okitsu, S.; Nishimura, S.; Kikuta, H.; Yamamoto, A.; Sugita, K.; et al. Distribution of rotavirus genotypes in Japan from 2015 to 2018: Diversity in genotypes before and after introduction of rotavirus vaccines. Vaccine 2020, 38, 3980–3986. [Google Scholar] [CrossRef]
- Tanaka, T.; Kamiya, H.; Asada, K.; Suga, S.; Ido, M.; Umemoto, M.; Ouchi, K.; Ito, H.; Kuroki, H.; Nakano, T.; et al. Changes in rotavirus genotypes before and after vaccine introduction: A multicenter, prospective observational study in three areas of Japan. Jpn. J. Infect. Dis. 2017, 70, 448–452. [Google Scholar] [CrossRef] [Green Version]
- Pasittungkul, S.; Lestari, F.B.; Puenpa, J.; Chuchaona, W.; Posuwan, N.; Chansaenroj, J.; Mauleekoonphairoj, J.; Sudhinaraset, N.; Wanlapakorn, N.; Poovorawan, Y. High prevalence of circulating DS-1-like human rotavirus A and genotype diversity in children with acute gastroenteritis in Thailand from 2016 to 2019. PeerJ 2021, 9, e10954. [Google Scholar] [CrossRef]
- Hoque, S.A.; Kobayashi, M.; Takanashi, S.; Anwar, K.S.; Watanabe, T.; Khamrin, P.; Okitsu, S.; Hayakawa, S.; Ushijima, H. Role of rotavirus vaccination on an emerging G8P [8] rotavirus strain causing an outbreak in central Japan. Vaccine 2018, 36, 43–49. [Google Scholar] [CrossRef] [PubMed]
- Kondo, K.; Tsugawa, T.; Ono, M.; Ohara, T.; Fujibayashi, S.; Tahara, Y.; Kubo, N.; Nakata, S.; Higashidate, Y.; Fujii, Y.; et al. Clinical and molecular characteristics of human rotavirus G8P [8] outbreak strain, Japan, 2014. Emerg. Infect. Dis. 2017, 23, 968. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Degiuseppe, J.I.; Stupka, J.A. Emergence of Unusual Rotavirus G9p [4] and G8p [8] Strains During Post Vaccination Surveillance in Argentina, 2017–2018. Res. Sq. 2020. [Google Scholar] [CrossRef]
- Huyen, D.T.T.; Hong, D.T.; Trung, N.T.; Hoa, T.T.N.; Oanh, N.K.; Thang, H.V.; Thao, N.T.T.; Hung, D.M.; Iijima, M.; Fox, K.; et al. Epidemiology of acute diarrhea caused by rotavirus in sentinel surveillance sites of Vietnam, 2012–2015. Vaccine 2018, 36, 7894–7900. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Yi, D.Y.; Lim, I.; Ward, A.C.; Kim, W. Detection of an unusual G8P [8] rotavirus in a Rotarix-vaccinated child with acute gastroenteritis using NanoporeMinION sequencing: A case report. Medicine 2020, 99, 40. [Google Scholar]
- Hoa-Tran, T.N.; Nakagomi, T.; Vu, H.M.; Do, L.P.; Gauchan, P.; Agbemabiese, C.A.; Nguyen, T.T.T.; Nakagomi, O.; Thanh, T.T.H. Abrupt emergence and predominance in Vietnam of rotavirus A strains possessing a bovine-like G8 on a DS-1-like background. Arch. Virol. 2016, 161, 479–482. [Google Scholar] [CrossRef] [PubMed]
- Moutelíková, R.; Sauer, P.; DvořákováHeroldová, M.; Holá, V.; Prodělalová, J. Emergence of Rare Bovine–Human Reassortant DS-1-Like Rotavirus A Strains with G8P [8] Genotype in Human Patients in the Czech Republic. Viruses 2019, 11, 1015. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.K.; Choi, S.; Shin, S.H.; Lee, E.J.; Hyun, J.; Kim, J.-S.; Kim, H.S. Emergence of G8P [6] rotavirus strains in Korean neonates. Gut. Pathog. 2018, 10, 1–8. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pre-Vaccine | Transitional | Post-Vaccine | p-Value | |
|---|---|---|---|---|
| Age group | ||||
| <1 years | 35/1145 (3.1%) | 20/955 (2.1%) | 28/1291 (2.2%) | 0.17 |
| 1 years | 507/3567 (14.2%) | 130/2301 (5.6%) | 75/1539 (4.9%) | <0.01 |
| 2 years | 619/2138 (29%) | 186/1174 (15.8%) | 78/927 (8.4%) | <0.01 |
| 3 years | 300/1024 (29.3%) | 111/686 (16.2%) | 65/697 (9.3%) | <0.01 |
| 4 years | 140/667 (21%) | 90/513 (17.5%) | 58/575 (10.1%) | <0.01 |
| 5–9 years | 157/1518 (10.3%) | 95/1103 (8.6%) | 115/1855 (6.2%) | <0.01 |
| 10–16 years | 19/657 (2.9%) | 18/408 (4.4%) | 11/734 (1.5%) | 0.07 |
| adults (>17 years) | 4/139 (2.8%) | 6/139 (3.2%) | 17/500 (3%) | 0.76 |
| Total | 1792/11009 (16.3%) | 676/7567 (8.9%) | 451/8326 (5.4%) | <0.01 |
| Roravirus genotypes | ||||
| G1P[8] | 188/467 (40.3%) | 186/564 (33.0%) | 18/355 (5.1%) | <0.01 |
| G2P[4] | 84/467 (18.0%) | 218/564 (38.7%) | 63/355 (17.7%) | 0.93 |
| G3P[8] | 81/467 (17.3%) | 27/564 (4.8%) | 22/355 (6.2%) | <0.01 |
| G8P[8] | - | - | 157/355 (44.2%) | <0.01 |
| G9P[8] | 42/467 (9.0%) | 51/564 (9.0%) | 54/355 (15.2%) | <0.01 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, K.g.; Kee, H.-y.; Park, H.j.; Chung, J.K.; Kim, T.s.; Kim, M.J. The Long-Term Impact of Rotavirus Vaccines in Korea, 2008–2020; Emergence of G8P[8] Strain. Vaccines 2021, 9, 406. https://doi.org/10.3390/vaccines9040406
Kim Kg, Kee H-y, Park Hj, Chung JK, Kim Ts, Kim MJ. The Long-Term Impact of Rotavirus Vaccines in Korea, 2008–2020; Emergence of G8P[8] Strain. Vaccines. 2021; 9(4):406. https://doi.org/10.3390/vaccines9040406
Chicago/Turabian StyleKim, Kwang gon, Hye-young Kee, Hye jung Park, Jae Keun Chung, Tae sun Kim, and Min Ji Kim. 2021. "The Long-Term Impact of Rotavirus Vaccines in Korea, 2008–2020; Emergence of G8P[8] Strain" Vaccines 9, no. 4: 406. https://doi.org/10.3390/vaccines9040406
APA StyleKim, K. g., Kee, H. -y., Park, H. j., Chung, J. K., Kim, T. s., & Kim, M. J. (2021). The Long-Term Impact of Rotavirus Vaccines in Korea, 2008–2020; Emergence of G8P[8] Strain. Vaccines, 9(4), 406. https://doi.org/10.3390/vaccines9040406