Biofouling Mitigation Approaches during Water Recovery from Fermented Broth via Forward Osmosis


 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Feedstocks and Treatment
2.1.1. Crude Glycerol
2.1.2. Organic Solvent Treatment of Crude Glycerol
2.1.3. Total Suspended Solids
2.2. Strain and Fermentation Conditions
2.3. Forward Osmosis Membranes and Operation
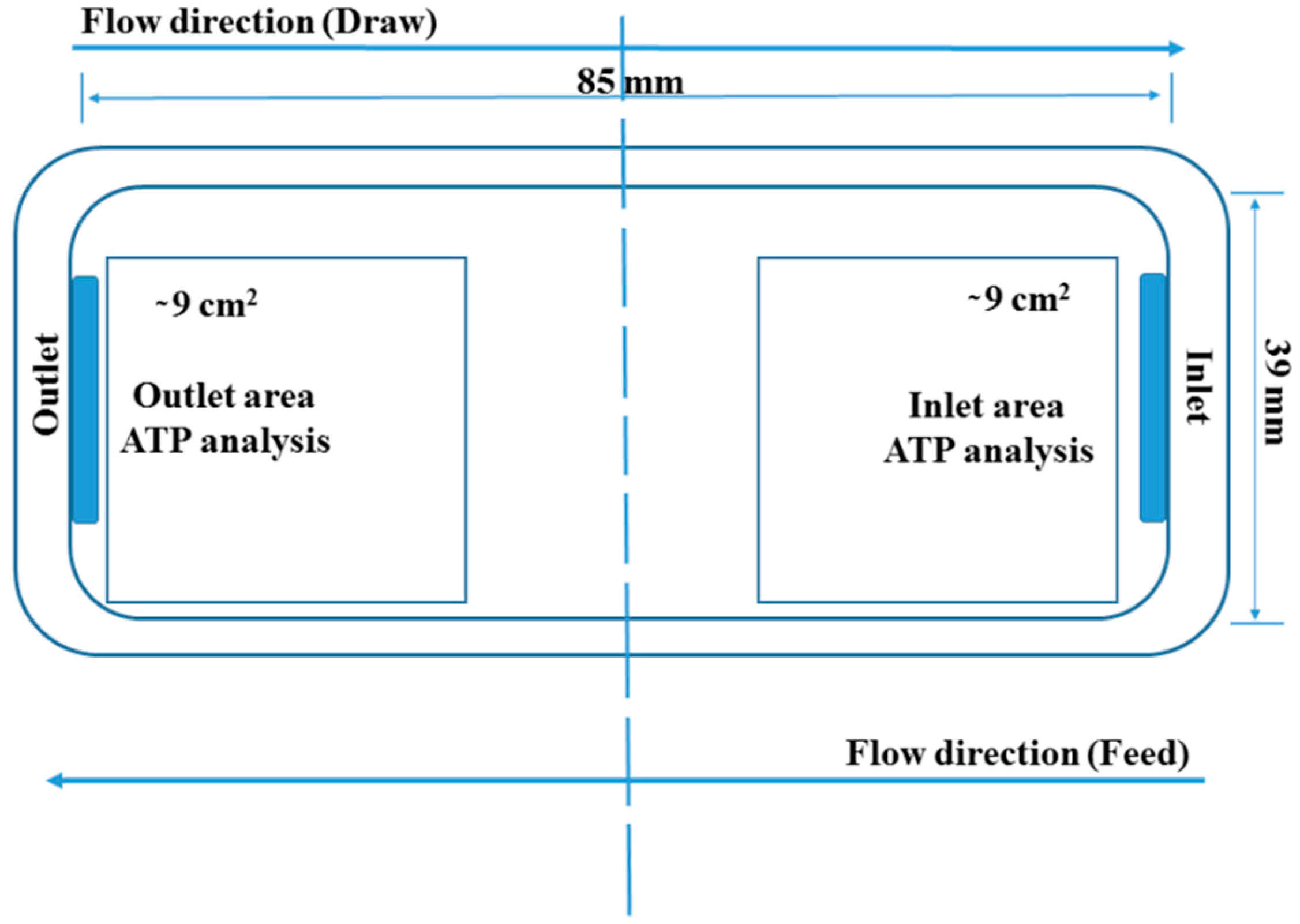
2.3.1. Membrane Coupons and Flow Cells
2.3.2. Operational Parameters
2.3.3. Forward Osmosis Baseline Experiments
2.4. Biofouling and Other Organic Fouling Experiments
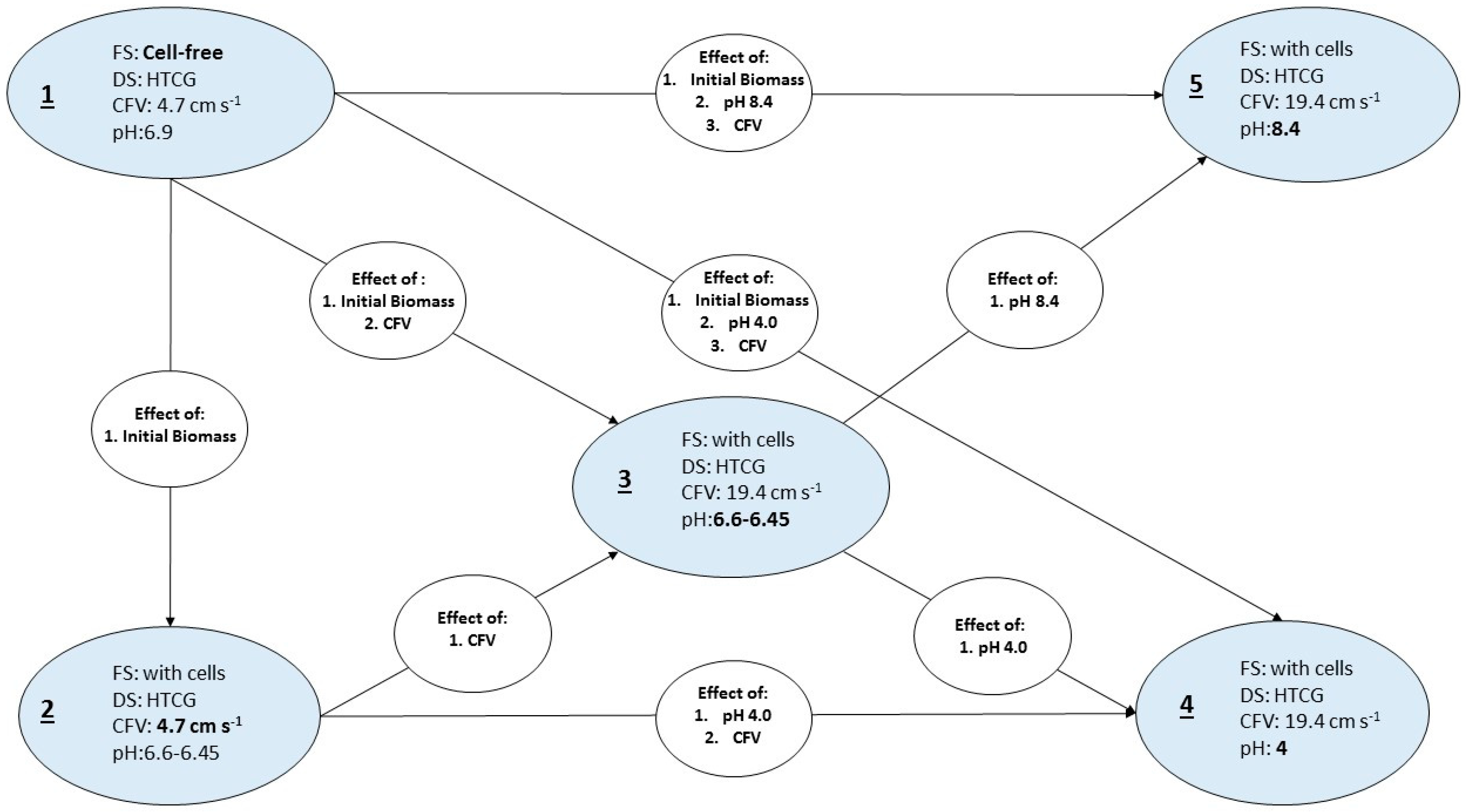
2.4.1. Experimental Parameters
2.4.2. ATP Analysis: Total and Free-ATP Quantification on the Membrane Surface
2.4.3. Calculations—Water Flux, Reverse Glycerol Flux, and Process Butanol Rejection
2.5. Analytical Methods
2.5.1. High Performance Liquid Chromatography (HPLC)
2.5.2. ATR-FTIR
2.5.3. Scanning Electron Microscopy (SEM)
3. Results and Discussion
3.1. Baseline Experiments
3.2. Biofouling
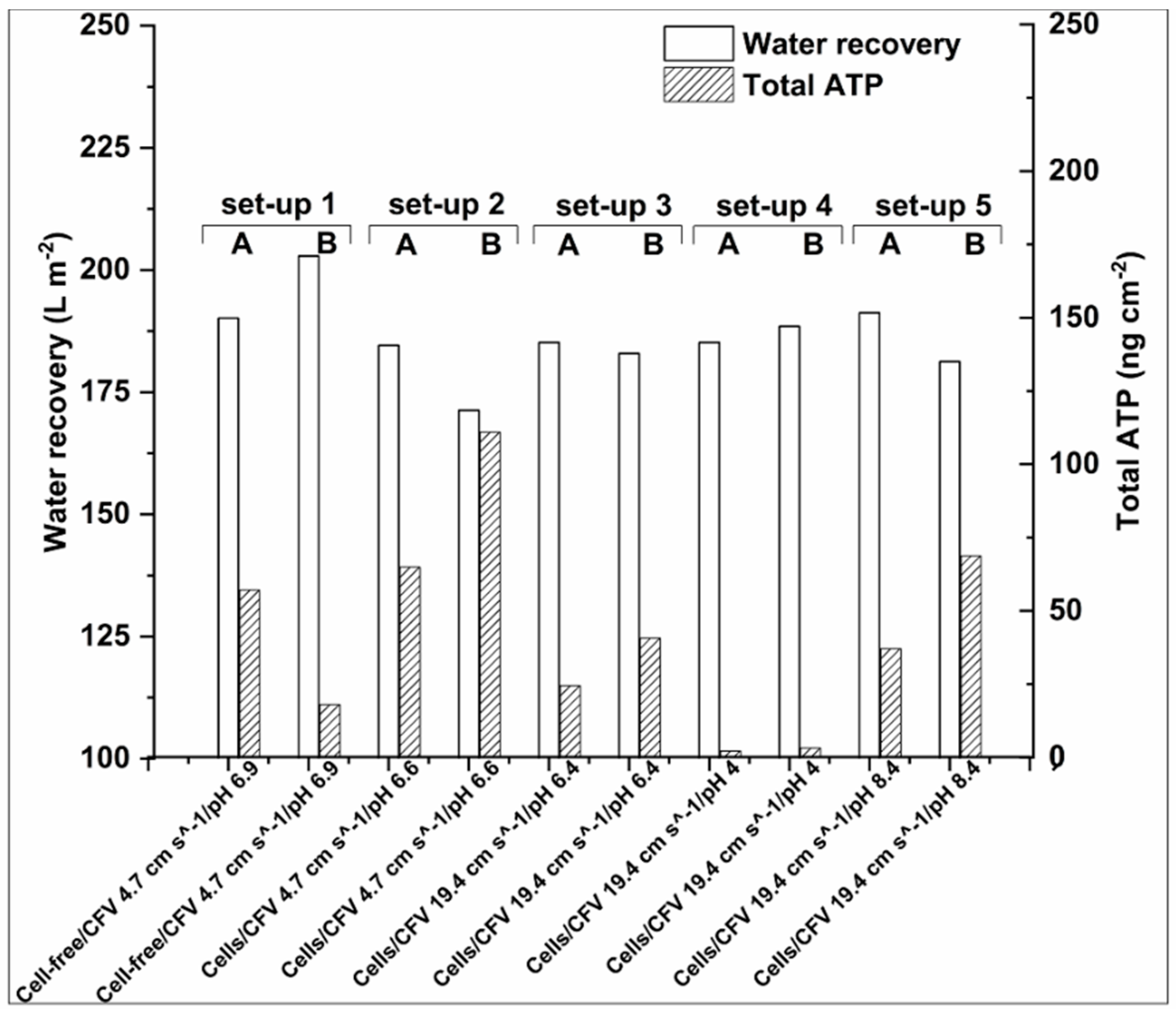
3.2.1. Water Recovery and ATP Accumulation
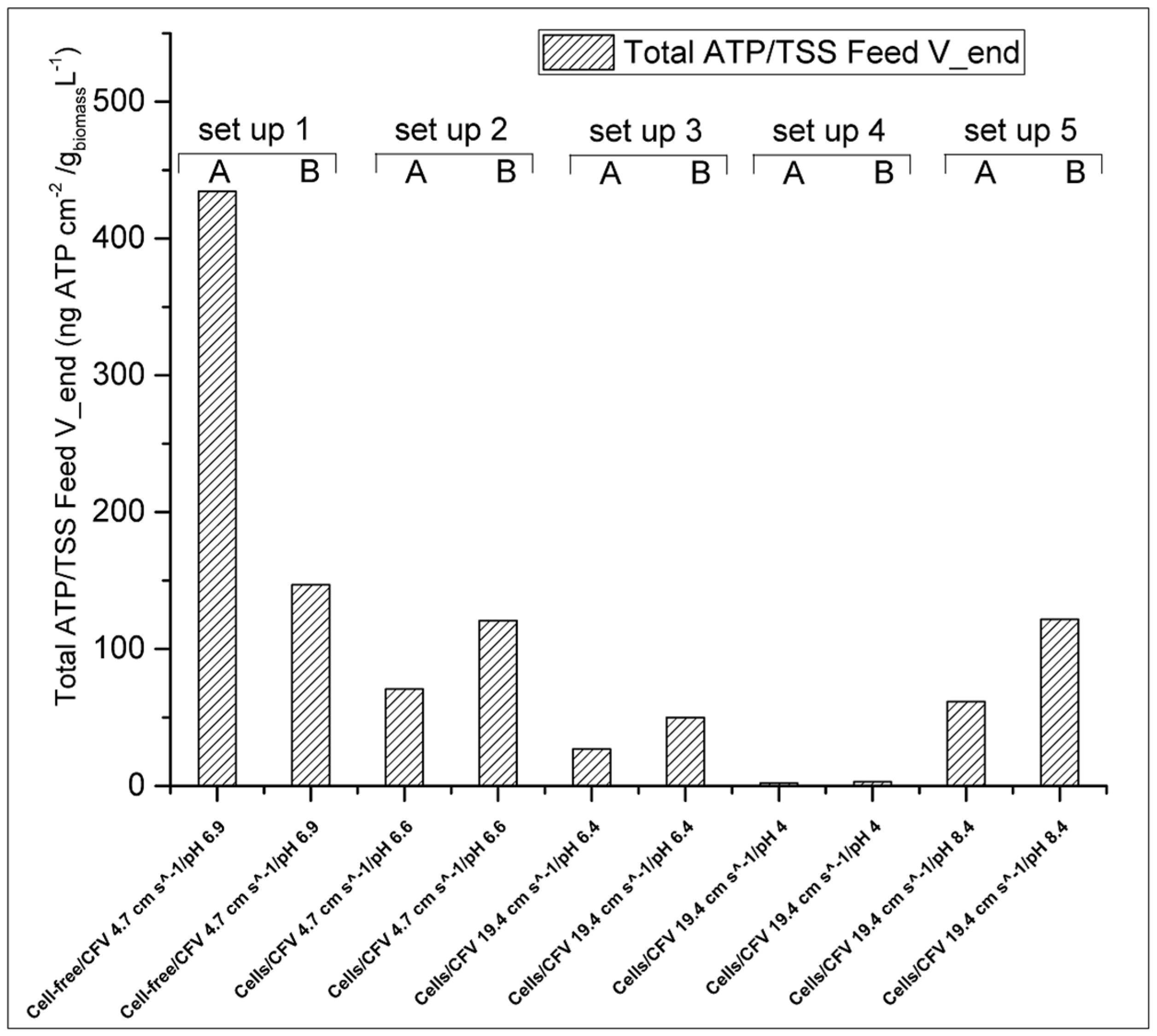
3.2.2. Effect of Initial Cell Concentration, Cross-Flow Velocity and pH in the Cellular Accumulation
3.2.3. Effect of Cellular Activity on Biofouling
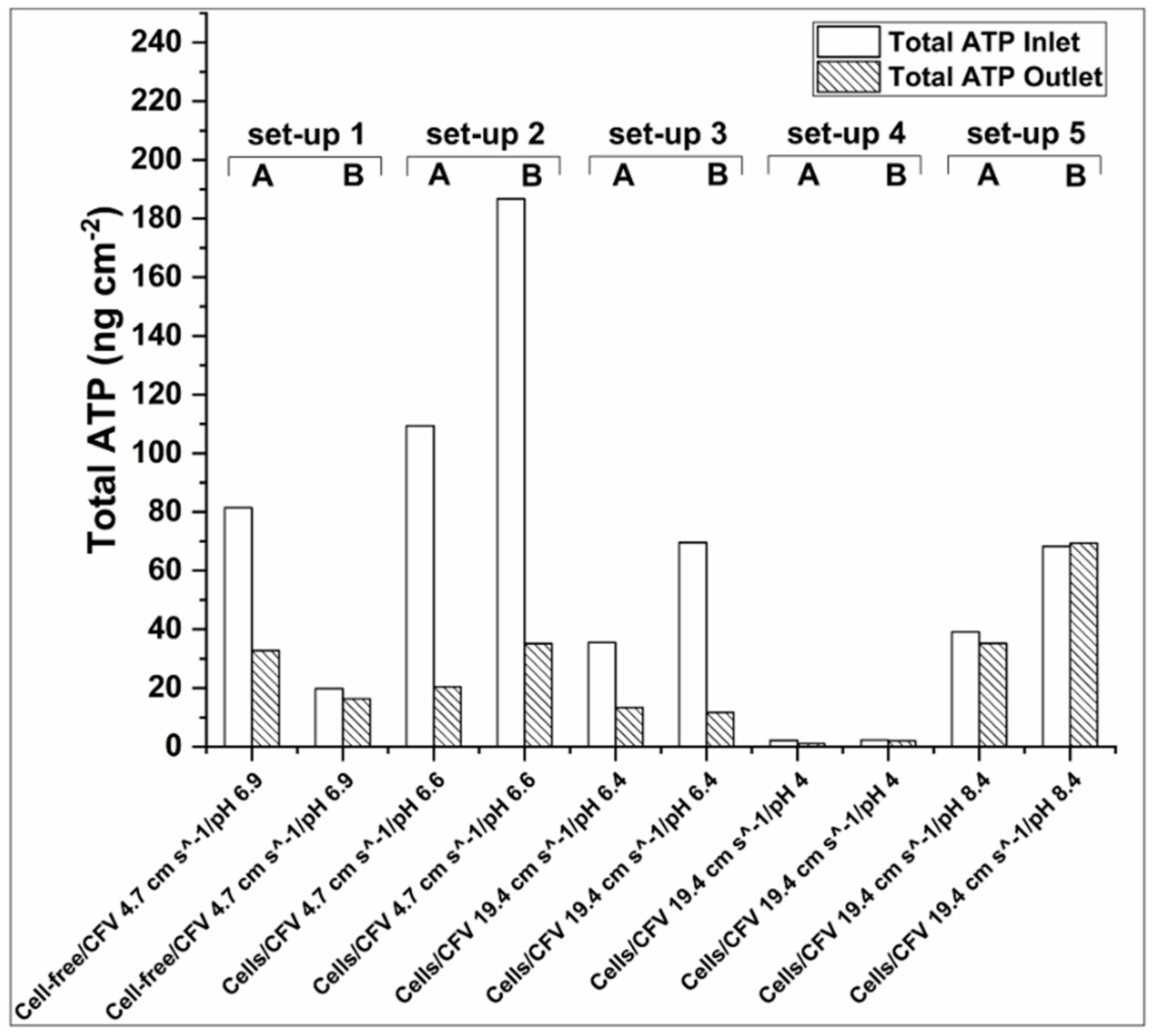
3.2.4. ATP Accumulation in the Membrane Inlet and Outlet
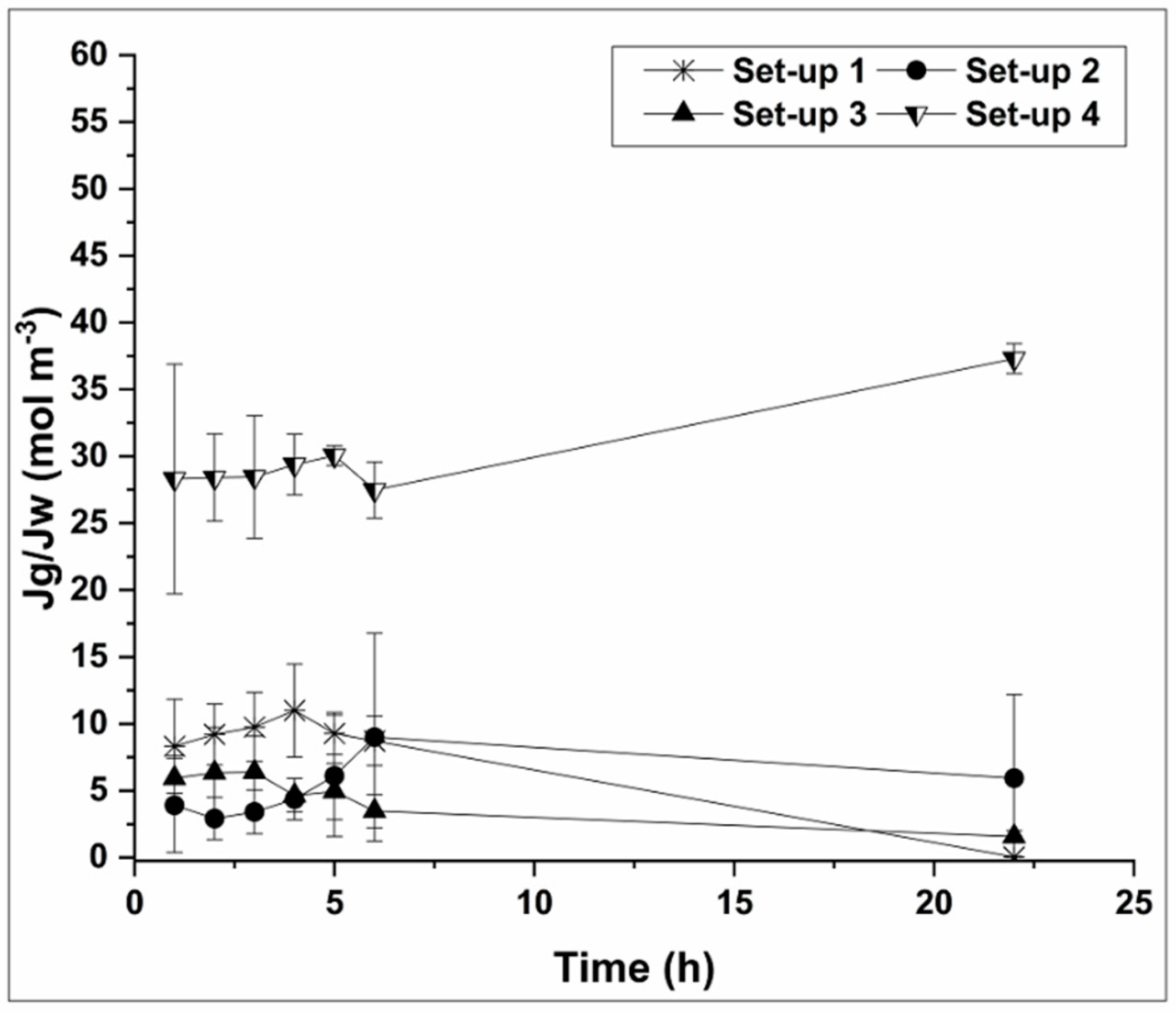
3.3. Specific Glycerol Reverse Flux and Process Butanol Rejection
4. Conclusions
- Water recovery can be directly linked with Total-ATP accumulation on the membrane. In the case of low pH other factors can affect the membrane blockage and water recovery, e.g., electrostatic repulsions/attractions between the membrane, the cells, and extracellular material in the feed solution.
- ATP analysis revealed that cells on the exponential growth face are more prone to attach on the membrane surface resulting in higher degree of biofouling.
- As revealed by ATR-FTIR analysis of the support layer of the membrane did not suffer from severe fouling (ATR-FTIR), or biofouling as demonstrated by the SEM study.
- Total-ATP was higher at the inlet than at the membrane outlet. This knowledge can be used when considering switching flow direction as a biofouling mitigation approach.
- Low pH has the greatest impact on cellular accumulation on the FO membrane surface. However, the highest specific glycerol reverse flux was obtained. This can be explained by the lack of biofilm development on the membrane surface and lower glycerol consumption by microorganisms.
- Process butanol rejection was found to be between 73 and 88% during 22 h filtration experiments.
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| A | membrane area (m−2) |
| AL-FS | active-side feed solution |
| ATP | adenosine tri-phosphate |
| ATR | attenuated total reflection |
| CFt | final solute concentration (M) |
| CFt0 | Initial solute concentration (M) |
| CFV | cross-flow velocity |
| CP | concentration polarization |
| DS | draw solution |
| ECP | external concentration polarization |
| EPS | extracellular polymeric substances |
| FO | forward osmosis |
| FS | feed solution |
| FT-IR | Fourier transform infrared spectroscopy |
| ICP | internal concentration polarization |
| Js | reverse solute flux (mol m−2 s−1) |
| Jsaver | average reverse solute flux (mol m−2 s−1) |
| Jg | reverse glycerol flux (mol m−2 s−1) |
| Js/Jw | specific reverse glycerol flux (mol m−3) |
| JWaver | average water flux (L m−2 s−1) |
| NF | nanofiltration |
| OMBR | osmotic pressure bioreactor |
| PA | polyamide |
| PES | polyethersulfone |
| RBuOH | Process butanol rejection (%) |
| RO | reverse osmosis |
| SEM | scanning electron microscopy |
| SRSF | specific reverse solute flux |
| TSS | total suspended solids |
| VFt | Final feed solution volume (L) |
| VFt0 | initial feed solution volume (L) |
| Greek letters | |
| Δt | time interval (s) |
| ΔV | volume change f feed solution (L) |
References
- Cath, T.Y.; Childress, A.E.; Elimelech, M. Forward osmosis: Principles, applications, and recent developments. J. Membr. Sci. 2006, 281, 70–87. [Google Scholar] [CrossRef]
- Hey, T.; Bajraktari, N.; Davidsson, Å.; Vogel, J.; Madsen, H.T.; Hélix-Nielsen, C.; Jansen, J.L.C.; Jönsson, K. Evaluation of direct membrane filtration and direct forward osmosis as concepts for compact and energy-positive municipal wastewater treatment. Environ. Technol. 2018, 39, 264–276. [Google Scholar] [CrossRef] [PubMed]
- Hey, T.; Zarebska, A.; Bajraktari, N.; Vogel, J.; Hélix-Nielsen, C.; la Cour Jansen, J.; Jönsson, K. Influences of mechanical pretreatment on the non-biological treatment of municipal wastewater by forward osmosis. Environ. Technol. 2017, 38, 2295–2304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valladares Linares, R.; Li, Z.; Sarp, S.; Bucs, S.S.; Amy, G.; Vrouwenvelder, J.S. Forward osmosis niches in seawater desalination and wastewater reuse. Water Res. 2014, 66, 122–139. [Google Scholar] [CrossRef] [PubMed]
- Kalafatakis, S.; Braekevelt, S.; Vilhelmsen Carlsen, N.S.; Lange, L.; Skiadas, I.V.; Gavala, H.N. On a novel strategy for water recovery and recirculation in biorefineries through application of forward osmosis membranes. Chem. Eng. J. 2017, 311, 209–216. [Google Scholar] [CrossRef] [Green Version]
- Chatzifragkou, A.; Papanikolaou, S. Effect of impurities in biodiesel-derived waste glycerol on the performance and feasibility of biotechnological processes. Appl. Microbiol. Biotechnol. 2012, 95, 13–27. [Google Scholar] [CrossRef]
- Santibáñez, C.; Varnero, M.T.; Bustamante, M. Residual Glycerol from Biodiesel Manufacturing, Waste or Potential Source of Bioenergy: A Review. Chil. J. Agric. Res. 2011, 71, 469–475. [Google Scholar] [CrossRef]
- Clomburg, J.M.; Gonzalez, R. Anaerobic fermentation of glycerol: A platform for renewable fuels and chemicals. Trends Biotechnol. 2013, 31, 20–28. [Google Scholar] [CrossRef] [Green Version]
- Johnson, E.; Sarchami, T.; Kießlich, S.; Munch, G.; Rehmann, L. Consolidating biofuel platforms through the fermentative bioconversion of crude glycerol to butanol. World J. Microbiol. Biotechnol. 2016, 32, 103. [Google Scholar] [CrossRef]
- Kalafatakis, S.; Braekevelt, S.; Lymperatou, A.; Zarebska, A.; Hélix-Nielsen, C.; Lange, L.; Skiadas, I.V.; Gavala, H.N. Use of forward osmsosis technology in crude glycerol fermentation biorefinery-Potential and Challenges. Bioprocess Biosyst. Eng. 2018, 41, 1089–1101. [Google Scholar] [CrossRef]
- She, Q.; Jin, X.; Li, Q.; Tang, C.Y. Relating reverse and forward solute diffusion to membrane fouling in osmotically driven membrane processes. Water Res. 2012, 46, 2478–2486. [Google Scholar] [CrossRef] [PubMed]
- Zhao, P.; Gao, B.; Yue, Q.; Liu, P.; Shon, H.K. Fatty acid fouling of forward osmosis membrane: Effects of pH, calcium, membrane orientation, initial permeate flux and foulant composition. J. Environ. Sci. 2015, 46, 55–62. [Google Scholar] [CrossRef] [PubMed]
- Bucs, S.S.; Valladares Linares, R.; Vrouwenvelder, J.S.; Picioreanu, C. Biofouling in forward osmosis systems: An experimental and numerical study. Water Res. 2016, 106, 86–97. [Google Scholar] [CrossRef] [PubMed]
- Yoon, H.; Baek, Y.; Yu, J.; Yoon, J. Biofouling occurrence process and its control in the forward osmosis. Desalination 2013, 325, 30–36. [Google Scholar] [CrossRef]
- Kwan, S.E.; Bar-Zeev, E.; Elimelech, M. Biofouling in forward osmosis and reverse osmosis: Measurements and mechanisms. J. Membr. Sci. 2015, 493, 703–708. [Google Scholar] [CrossRef]
- Cath, T.Y.; Hancock, N.T.; Lundin, C.D.; Hoppe-Jones, C.; Drewes, J.E. A multi-barrier osmotic dilution process for simultaneous desalination and purification of impaired water. J. Membr. Sci. 2010, 362, 417–426. [Google Scholar] [CrossRef]
- Coday, B.D.; Yaffe, B.G.M.; Xu, P.; Cath, T.Y. Rejection of trace organic compounds by forward osmosis membranes: A literature review. Environ. Sci. Technol. 2014, 48, 3612–3624. [Google Scholar] [CrossRef]
- Wang, Q.; Hu, M.; Wang, Z.; Hu, W.; Cao, J.; Wu, Z.-C. Uniqueness of biofouling in forward osmosis systems: Mechanisms and control. Crit. Rev. Environ. Sci. Technol. 2018, 48, 1031–1066. [Google Scholar] [CrossRef]
- Chun, Y.; Mulcahy, D.; Zou, L.; Kim, I.S. A Short Review of Membrane Fouling in Forward Osmosis Processes. Membranes 2017, 7, 30. [Google Scholar] [CrossRef] [Green Version]
- Lee, C.; Nguyen, T.-T.; Adha, R.S.; Shon, H.K.; Kim, I. Influence of hydrodynamic operating conditions on organic fouling of spiral-wound forward osmosis membranes: Fouling-induced performance deterioration in FO-RO hybrid system. Water Res. 2020, 185, 116154. [Google Scholar] [CrossRef]
- Vrouwenvelder, J.S.; Beyer, F.; Dahmani, K.; Hasan, N.; Galjaard, G.; Kruithof, J.C.; Van Loosdrecht, M.C.M. Phosphate limitation to control biofouling. Water Res. 2010, 44, 3454–3466. [Google Scholar] [CrossRef]
- She, Q.; Wang, R.; Fane, A.G.; Tang, C.Y. Membrane fouling in osmotically driven membrane processes: A review. J. Membr. Sci. 2016, 499, 201–233. [Google Scholar] [CrossRef]
- Zarebska, A.; Petrinic, I.; Korenak, J.; Buksek, H.; Ciszewska-Kaluzka, A.; Hélix-Nielsen, C. Influence of mechanical wastewater pretreatment on membrane fouling during municipal wastewater treatment by forward osmosis. In Proceedings of the 9th International Membrane Science & Technology Conference, Adelaide, Australia, 5–8 December 2016; pp. 467–470. [Google Scholar]
- Mi, B.; Elimelech, M. Chemical and physical aspects of organic fouling of forward osmosis membranes. J. Membr. Sci. 2008, 320, 292–302. [Google Scholar] [CrossRef]
- Bucs, S.S.; Radu, A.I.; Lavric, V.; Vrouwenvelder, J.S.; Picioreanu, C. Effect of different commercial feed spacers on biofouling of reverse osmosis membrane systems: A numerical study. Desalination 2014, 343, 26–37. [Google Scholar] [CrossRef]
- Varrone, C.; Heggeset, T.M.B.; Le, S.B.; Haugen, T.; Markussen, S.; Skiadas, I.V.; Gavala, H.N. Comparison of Different Strategies for Selection/Adaptation of Mixed Microbial Cultures Able to Ferment Crude Glycerol Derived from Second-Generation Biodiesel. Biomed. Res. Int. 2015, 2015, 14. [Google Scholar] [CrossRef] [Green Version]
- Anand, P.; Saxena, R.K. A comparative study of solvent-assisted pretreatment of biodiesel derived crude glycerol on growth and 1,3-propanediol production from Citrobacter freundii. N. Biotechnol. 2012, 29, 199–205. [Google Scholar] [CrossRef]
- APHA American Public Health Association. Standard Methods for Examination of Water and Wastewater, 21st ed.; APHA, AWWA, WPCF: Washington, DC, USA, 2005. [Google Scholar]
- Biebl, H. Fermentation of glycerol by Clostridium pasteurianum-batch and continuous culture studies. J. Ind. Microbiol. Biotechnol. 2001, 27, 18–26. [Google Scholar] [CrossRef] [PubMed]
- Dabrock, B.; Bahl, H.; Gottschalk, G. Parameters affecting solvent production by Clostridium pasteurianum. Appl. Environ. Microbiol. 1992, 58, 1233–1239. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Qiu, C.; Li, X.; Vararattanavech, A.; Shen, W.; Torres, J.; Hélix-Nielsen, C.; Wang, R.; Hu, X.; Fane, A.G.; et al. Synthesis of robust and high-performance aquaporin-based biomimetic membranes by interfacial polymerization-membrane preparation and RO performance characterization. J. Membr. Sci. 2012, 423–424, 422–428. [Google Scholar] [CrossRef]
- Blandin, G.; Verliefde, A.R.D.; Comas, J.; Rodriguez-Roda, I.; Le-Clech, P. Efficiently combining water reuse and desalination through forward osmosis-reverse osmosis (FO-RO) hybrids: A critical review. Membranes 2016, 6, 37. [Google Scholar] [CrossRef] [Green Version]
- Vrouwenvelder, J.S.; Graf von der Schulenburg, D.A.; Kruithof, J.C.; Johns, M.L.; van Loosdrecht, M.C.M. Biofouling of spiral-wound nanofiltration and reverse osmosis membranes: A feed spacer problem. Water Res. 2009, 43, 583–594. [Google Scholar] [CrossRef] [PubMed]
- Vang, Ó.K.; Corfitzen, C.B.; Smith, C.; Albrechtsen, H.-J. Evaluation of ATP measurements to detect microbial ingress by wastewater and surface water in drinking water. Water Res. 2014, 64, 309–320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Ning, Z.; Wang, D.K.; Diniz da Costa, J.C. A novel ethanol dehydration process by forward osmosis. Chem. Eng. J. 2013, 232, 397–404. [Google Scholar] [CrossRef]
- Zarebska, A.; Amor, Á.C.; Ciurkot, K.; Karring, H.; Thygesen, O.; Andersen, T.P.; Hägg, M.B.; Christensen, K.V.; Norddahl, B. Fouling mitigation in membrane distillation processes during ammonia stripping from pig manure. J. Membr. Sci. 2015, 484, 119–132. [Google Scholar] [CrossRef]
- Borgnia, M.J.; Kozono, D.; Calamita, G.; Maloney, P.C.; Agre, P. Functional reconstitution and characterization of AqpZ, the E. coli water channel protein. J. Mol. Biol. 1999, 291, 1169–1179. [Google Scholar] [CrossRef]
- Hammes, F.; Goldschmidt, F.; Vital, M.; Wang, Y.; Egli, T. Measurement and interpretation of microbial adenosine tri-phosphate (ATP) in aquatic environments. Water Res. 2010, 44, 3915–3923. [Google Scholar] [CrossRef]
- Stanley, P.E. Extraction of Adenosine triphosphate form microbial and somatic cells. Methods Enzym. 1986, 133, 14–22. [Google Scholar]
- Bucs, S.S.; Valladares Linares, R.; van Loosdrecht, M.C.M.; Kruithof, J.C.; Vrouwenvelder, J.S. Impact of organic nutrient load on biomass accumulation, feed channel pressure drop increase and permeate flux decline in membrane systems. Water Res. 2014, 67, 227–242. [Google Scholar] [CrossRef]
- D’Haese, A.; Le-Clech, P.; Van Nevel, S.; Verbeken, K.; Cornelissen, E.R.; Khan, S.J.; Verliefde, A.R.D. Trace organic solutes in closed-loop forward osmosis applications: Influence of membrane fouling and modeling of solute build-up. Water Res. 2013, 47, 5232–5244. [Google Scholar] [CrossRef]
- Vrouwenvelder, J.S.; van Paassen, J.A.M.; Wessels, L.P.; van Dam, A.F.; Bakker, S.M. The Membrane Fouling Simulator: A practical tool for fouling prediction and control. J. Membr. Sci. 2006, 281, 316–324. [Google Scholar] [CrossRef]
- Nguyen, T.; Roddick, F.A.; Fan, L. Biofouling of water treatment membranes: A review of the underlying causes, monitoring techniques and control measures. Membranes 2012, 2, 804–840. [Google Scholar] [CrossRef] [Green Version]
- Subramani, A.; Hoek, E.M. V Direct observation of initial microbial deposition onto reverse osmosis and nanofiltration membranes. J. Membr. Sci. 2008, 319, 111–125. [Google Scholar] [CrossRef]
- Yuan, B.; Wang, X.; Tang, C.; Li, X.; Yu, G. In situ observation of the growth of biofouling layer in osmotic membrane bioreactors by multiple fluorescence labeling and confocal laser scanning microscopy. Water Res. 2015, 75, 188–200. [Google Scholar] [CrossRef]
- Freger, V.; Gilron, J.; Belfer, S. TFC polyamide membranes modified by grafting of hydrophilic polymers: An FT-IR/AFM/TEM study. J. Membr. Sci. 2002, 209, 283–292. [Google Scholar] [CrossRef]
- Kwon, Y.N.; Leckie, J.O. Hypochlorite degradation of crosslinked polyamide membranes. II. Changes in hydrogen bonding behavior and performance. J. Membr. Sci. 2006, 282, 456–464. [Google Scholar] [CrossRef]
- Tang, C.Y.; Kwon, Y.-N.; Leckie, J.O. Effect of membrane chemistry and coating layer on physiochemical properties of thin film composite polyamide RO and NF membranes. Desalination 2009, 242, 168–182. [Google Scholar] [CrossRef]
- McCutcheon, J.; Hoek, E.; Bui, N.; Lind Mary, L. Nanostructured Membranes for Engineered Osmosis Applications. U.S. Patent Application 13/508,892, 2 May 2013. Available online: https://patents.google.com/patent/US20130105395A1/en (accessed on 1 October 2020).
- Maruyama, T.; Katoh, S.; Nakajima, M.; Nabetani, H.; Abbott, T.P.; Shono, A.; Satoh, K. FT-IR analysis of BSA fouled on ultrafiltration and microfiltration membranes. J. Membr. Sci. 2001, 192, 201–207. [Google Scholar] [CrossRef]
- Xu, P.; Drewes, J.E.; Kim, T.U.; Bellona, C.; Amy, G. Effect of membrane fouling on transport of organic contaminants in NF/RO membrane applications. J. Membr. Sci. 2006, 279, 165–175. [Google Scholar] [CrossRef]
- Hurwitz, G.; Guillen, G.R.; Hoek, E.M. V Probing polyamide membrane surface charge, zeta potential, wettability, and hydrophilicity with contact angle measurements. J. Membr. Sci. 2010, 349, 349–357. [Google Scholar] [CrossRef]
- Boo, C.; Elimelech, M.; Hong, S. Fouling control in a forward osmosis process integrating seawater desalination and wastewater reclamation. J. Membr. Sci. 2013, 444, 148–156. [Google Scholar] [CrossRef]
- Habash, M.B.; Van Der Mei, H.C.; Reid, G.; Busscher, H.J. Adhesion of Pseudomonas aeruginosa to silicone rubber in a parallel plate flow chamber in the absence and presence of nutrient broth. Microbiology 1997, 143, 2569–2574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walker, S.L.; Hill, J.E.; Redman, J.A.; Elimelech, M. Influence of growth phase on adhesion kinetics of Escherichia coli D21g. Appl. Environ. Microbiol. 2005, 71, 3093–3099. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Araújo, P.A.; Miller, D.J.; Correia, P.B.; Van Loosdrecht, M.C.M.; Kruithof, J.C.; Freeman, B.D.; Paul, D.R.; Vrouwenvelder, J.S. Impact of feed spacer and membrane modification by hydrophilic, bactericidal and biocidal coating on biofouling control. Desalination 2012, 295, 1–10. [Google Scholar] [CrossRef]
- Wang, X.; Zhao, Y.; Yuan, B.; Wang, Z.; Li, X.; Ren, Y. Comparison of biofouling mechanisms between cellulose triacetate (CTA) and thin-film composite (TFC) polyamide forward osmosis membranes in osmotic membrane bioreactors. Bioresour. Technol. 2016, 202, 50–58. [Google Scholar] [CrossRef]
- Dreszer, C.; Wexler, A.D.; Drusová, S.; Overdijk, T.; Zwijnenburg, A.; Flemming, H.C.; Kruithof, J.C.; Vrouwenvelder, J.S. In-situ biofilm characterization in membrane systems using Optical Coherence Tomography: Formation, structure, detachment and impact of flux change. Water Res. 2014, 67, 243–254. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Baseline Setup | CFV (cm s−1) | JWaver (L m−2 h−1) | JSaver (g m−2 h−1) | JS/JW (g L−1) |
|---|---|---|---|---|
| 1A | 4.7 | 10.04 | 0.82 | 0.08 |
| 1B | 4.7 | 9.81 | 0.87 | 0.09 |
| 2A | 4.7 | 8.80 | 2.30 | 0.26 |
| 2B | 4.7 | 10.61 | 1.82 | 0.17 |
| 3A | 19.4 | 10.36 | 2.74 | 0.26 |
| 3B | 19.4 | 10.38 | 2.64 | 0.25 |
| 4A | 19.4 | 12.67 | 2.84 | 0.22 |
| 4B | 19.4 | 10.95 | 2.67 | 0.24 |
| 5A | 19.4 | 11.14 | 3.98 | 0.36 |
| 5B | 19.4 | 10.61 | 2.92 | 0.28 |
| Setup | pH Feedini | pH Feedend |
|---|---|---|
| 1A | 6.9 | 6.5 |
| 1B | 6.9 | 6.5 |
| 2A | 6.6 | 6.5 |
| 2B | 6.45 | 6.45 |
| 3A | 6.4 | 6.55 |
| 3B | 6.4 | 6.5 |
| 4A | 4.00 | 3.97 |
| 4B | 4 | 4.1 |
| 5A | 8.42 | 5.8 |
| 5B | 8.50 | 6.10 |
| Setup | Intracellular ATP/Total ATP |
|---|---|
| 1A | 0.95 |
| 1B | 0.79 |
| 2A | 0.25 |
| 2B | 0.42 |
| 3A | 0.26 |
| 3B | 0.47 |
| 4A | 0.00 1 |
| 4B | 0.00 1 |
| 5A | 0.42 |
| 5B | 0.42 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kalafatakis, S.; Zarebska, A.; Lange, L.; Hélix-Nielsen, C.; Skiadas, I.V.; Gavala, H.N. Biofouling Mitigation Approaches during Water Recovery from Fermented Broth via Forward Osmosis. Membranes 2020, 10, 307. https://doi.org/10.3390/membranes10110307
Kalafatakis S, Zarebska A, Lange L, Hélix-Nielsen C, Skiadas IV, Gavala HN. Biofouling Mitigation Approaches during Water Recovery from Fermented Broth via Forward Osmosis. Membranes. 2020; 10(11):307. https://doi.org/10.3390/membranes10110307
Chicago/Turabian StyleKalafatakis, Stavros, Agata Zarebska, Lene Lange, Claus Hélix-Nielsen, Ioannis V. Skiadas, and Hariklia N. Gavala. 2020. "Biofouling Mitigation Approaches during Water Recovery from Fermented Broth via Forward Osmosis" Membranes 10, no. 11: 307. https://doi.org/10.3390/membranes10110307
APA StyleKalafatakis, S., Zarebska, A., Lange, L., Hélix-Nielsen, C., Skiadas, I. V., & Gavala, H. N. (2020). Biofouling Mitigation Approaches during Water Recovery from Fermented Broth via Forward Osmosis. Membranes, 10(11), 307. https://doi.org/10.3390/membranes10110307