1. Introduction
The spread of food allergies in developed countries has become one of the most serious health issues. Today, about 30% of the world population suffers from allergic-related diseases [
1,
2]. IgE-mediated allergic reaction is one of the most common and severe types of allergic reactions to foodstuffs, and it is characterized by elevated allergen-specific IgE levels in blood of atopic patients [
3]. The IgE-mediated food-allergic reactions result in skin (urticaria, angioedema), respiratory tract (rhinitis, asthma), gastrointestinal tract (abdominal pain, diarrhea), and systemic anaphylactic reactions [
4,
5].
Legumes are a staple food in the diet of many countries due to their high protein content and low price. Widespread consumption of legumes has led to an increase in the prevalence of legume allergy. Additionally, there is a high rate of cross-reactivity between different legume allergens. Most previous studies have been focused on 2 members of the legume food family, peanuts and soybeans, but lentils should also be examined in detail as a basic food in the Mediterranean area, Near East, India, Pakistan and the United States [
6]. It has been shown that allergy to lentil is the most frequently diagnosed legume allergy in children from a Mediterranean area [
7]. Allergy to lentils started early in life and is characterized by predominantly mild allergic symptoms, like oral allergy syndrome and urticaria; however, in some cases, lentils are able to cause severe systemic reactions like anaphylaxis [
8,
9].
Previously, a new allergen Len c 3 belonging to the class of lipid transfer proteins (LTPs) was isolated from the lentil
Lens culinaris seeds [
10]. We have shown that Len c 3 was cross-reacting with the major peach allergen Pru p 3 [
10]. In Len c 3, four conserved disulfide bridges stabilize a compact spatial structure containing four helices and a central hydrophobic cavity [
11]. The protein is able to bind with various effectiveness of such ligands as fatty acids (FAs) and lysolipids in solutions with different pHs [
11,
12]. This allergen is sensitive to heating and duodenal digestion, but the presence of its diverse ligands may increase the protein thermostability, decrease the rate of its gastroduodenal degradation, but not affect the IgE-binding capacity of the allergen [
13].
Site-directed mutagenesis can be used to identify key amino acid residues, either responsible for the ability of LTPs to bind different lipid ligands [
14], or playing an important role in the IgE-binding capacity of the proteins [
15]. In our previous work, we assumed that the same amino acid residues can participate both in the interaction with lipid ligands and specific IgE from the sera of allergic patients [
16]. Therefore, the main goal of this study was to investigate the effect of point amino acid substitutions on the structural and allergenic properties of Len c 3. With this aim in view, we obtained and characterized recombinant analogues of the allergen with single or double substitutions of Thr41, Arg45 and Tyr80, which are located close to each other near the “bottom” entrance to the hydrophobic cavity of Len c 3 and are apparently included in the IgE-binding epitope of the allergen.
2. Materials and Methods
2.1. Materials
2-p-toluidinonaphthalene-6-sulphonate (TNS) was purchased from Sigma-Aldrich (St. Louis, MO, USA). Digestive enzymes (porcine pepsin and trypsin, bovine α-chymotrypsin) were purchased from Sigma-Aldrich (St. Louis, MO, USA).
Len c 3 and its R45A, Y80A and R45A/Y80A analogues were produced as earlier described [
11]. The T41A and T41A/Y80A analogues were obtained by site-directed mutagenesis of the original plasmid pET-His8-TrxL-Len c 3 using full-length inverse PCR amplification with mutagenizing primers (
Supplementary Materials, Table S1). All recombinant proteins were overexpressed in
Escherichia coli and purified as described previously [
11]. Homogeneity and identity of the recombinant protein samples were confirmed by MALDI mass spectrometry (
Supplementary Materials, Figure S1). Rabbit polyclonal anti-Len c 3 antibodies were obtained as previously described [
17].
Sera from patients (n = 100) with legume and other plant food allergy were obtained from the Clinical Diagnostic Center of the G.N. Gabrichevsky Research Institute for Epidemiology and Microbiology. The work with human sera was approved by the local ethical committees of the Institute (FS-99-01-009026, Moscow, Russia). The amount of specific IgE to allergen extracts in the patient sera was determined by using RIDA qLine Allergy Panel 1–4 (R-Biopharm, Pfungstadt, Germany). Screening of sera containing specific IgE (sIgE) to Len c 3 was performed by Enzyme-Linked Immunosorbent Assay (ELISA). Sera samples from non-allergic individuals were used as a negative control. Sera of 14 allergic patients containing sIgE to Len c 3 were selected for experiments on IgE-binding with the recombinant analogues carrying amino acid substitutions. Seven serum samples were used in experiments with the T41A, Y80A and T41A/Y80A analogues, and the others were used in experiments with the R45A, Y80A and R45A/Y80A analogues.
2.2. Circular Dichroism Spectroscopy
Circular dichroism spectra were recorded at 25 °C using a J-810 spectropolarimeter (Jasco) in a 0.1 cm path length quartz cell (Hellma GmbH & Co. KG, Mullheim, Germany) in a wavelength range of 190–250 nm (scan rate 1 nm) using solutions of the recombinant proteins in 10 mM phosphate buffer, pH 7.4, at a concentration of 17 mM.
2.3. Ligand Binding
An ability of Len c 3 and its mutant analogues (T41A, R45A, Y80A, T41A/Y80A, R45A/Y80A) to bind lipids was assessed using the TNS fluorescent probe as previously described [
18]. Fluorescence intensity was measured at 437 nm with excitation at 320 nm using F-2710 spectrofluorometer (Hitachi High Technologies America Inc., Pleasanton, CA, USA). TNS (4 µM) with or without a lipid (4 µM) was incubated for 1 min in a stirred cuvette containing 2 mL of the 10 mM phosphate buffer (pH 7.4) with gentle mixing before the initial fluorescence (F
0) was recorded. Then, Len c 3 or its mutant analogues (4 µM) were added, and 2 min later the fluorescence was recorded at equilibrium (F). Experiments were performed in triplicate. The results were expressed as a percentage of the allergen-TNS complex fluorescence calculated according to the formula [(F − F
0)/F
C] × 100%, where F
C is the fluorescence of the allergen-TNS complex in the absence of a lipid.
2.4. Gastroduodenal Digestion In Vitro
Gastroduodenal digestion of the recombinant Len c 3 and its mutant analogues in vitro was simulated as described with some modifications [
13]. Cleavage mimicking the protein gastric digestion in vitro was performed for 2 h at 37 °C using 50 ng (0.1 U) of pepsin per 1 μg of Len c 3 or its mutant analogues in 0.05 M HCl, pH 2.0 (at the final protein concentration of 0.05 mM). For cleavage mimicking subsequent protein duodenal digestion in vitro, pH of the mixture resulting from gastric digestion was adjusted to 8.0 by addition of 0.1 M ammonium bicarbonate and incubated for 24 h at 37 °C with 2.5 ng (0.03 U) of trypsin and 10 ng (0.4 × 10
−3 U) of α-chymotrypsin per 1 μg of the substrate (Len c 3 or its mutant analogues). Digestion of all the proteins was monitored by sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) [
19]. SDS-PAGE was carried out in Tris-Glycine electrode buffer, pH 8.3, using disk-gels with 3% stacking and 15% separating gels. All samples were mixed with loading buffer containing 2-mercaptoethanol besides SDS. Each experiment was carried out three times. Analysis of the obtained polyacrylamide gels was performed using Gel Doc XR
+ imaging system (Bio-Rad, Hercules, CA, USA) and Image Lab Software.
2.5. Bioinformatic Approaches to Study Structural Properties of Mutant Analogues of Len c 3
Spatial structure of the wild-type lentil Len c 3 has been solved by us previously [PDB ID 2MAL] using heteronuclear NMR in solution [
11]. This structure was used in the current study as a template. The mutant analogues T41A, R45A, Y80A, T41A/Y80A, and R45A/Y80A were generated by means of the mutagenesis tool of the PyMOL 1.8.2.0 software (Schrödinger, LLC, New York, NY, USA) [
20]. The hydrophobic pockets’ volumes were calculated using the MOLE online tool (
https://mole.upol.cz/, accession date 21 October 2021) with a 5 Å probe radius [
21]. Proteins were visualized by the PyMOL 1.8.2.0 software, and hydrophobicity coloring was made by using
color_h script, which was based on a normalized consensus Eisenberg scale [
22].
2.6. Immunoglobulin Binding Assay
Comparative IgE-binding assay was performed by ELISA using sera from allergic patients containing specific IgE to Len c 3. For that, plate wells (Corning Incorporated, Corning, NY, USA) were coated with the recombinant Len c 3 and its mutant analogues (0.5 mg/well) in 0.01 M phosphate-buffered saline (PBS), pH 7.4, for 1 h, at 37 °C; saturated with 2% bovine serum albumin (BSA, SERVA, Heidelberg, Germany) in PBS buffer for 1 h, at 37 °C; and then incubated with sera of allergic patients (a dilution ratio of 1:5) overnight, at 4 °C. sIgE-binding was detected by using the peroxidase-conjugated anti-human IgE from goat (a dilution ratio of 1:2000, Sigma) in PBS with 0.5% BSA, and 3,3′,5,5′-tetramethylbenzidine (TMB) liquid substrate system for ELISA (Sigma). The enzymatic reaction was stopped after 30 min by 2 N H2SO4, and absorbance values were determined at 450 nm. PBS, containing 0.05% Tween-20 (PBS-T), was used as a washing solution on each step. Each experiment was carried out twice.
ELISA assays with the polyclonal anti-Len c 3 rabbit antiserum were performed as described above with some modifications. After blocking free binding sites, the plate wells were incubated with the anti-Len c 3 rabbit antiserum in PBS (from 1:400 to 1:735,306 serial dilutions) for 1 h at 37 °C. The peroxidase-conjugated anti-rabbit antibodies from goat (a dilution ratio of 1:50,000, Sigma) in PBS with 0.5% BSA were used for detection. Each experiment was carried out three times.
4. Discussion
Lipid transfer proteins (LTPs) represent one of the most clinically significant classes of plant allergens. LTPs have a compact structure stabilized by four disulfide bonds, which provide them with a high stability. An important characteristic feature of these allergens is the presence of a hydrophobic cavity capable of accommodating different lipid ligands. The formation of complexes with ligands can affect the structure and allergenic properties of plant LTPs.
Earlier, it was shown that substitutions of such amino acids as Arg39, Thr40 and Arg44 in Pru p 3 structure led to substantial decrease (approximately 5 times) of IgE-binding capacity of this peach allergen [
15]. It was also demonstrated that two protein regions 35–46 and 76–79 comprise major IgE-binding conformational epitope of Pru p 3 [
24]. In the other publications, an important role of such amino acids as Asn35, Arg44 of Pru p 3 [
25] and Arg46, Tyr81 of walnut allergen Jug r 3 and maize allergen Zea m 14 (walnut numbering) in the binding of lipid ligands was shown [
26]. However, in our previous work [
16], we assumed that the same amino acids in the LTP structure may play a key role both in the interaction with lipid ligands and specific IgE from sera of allergic patients.
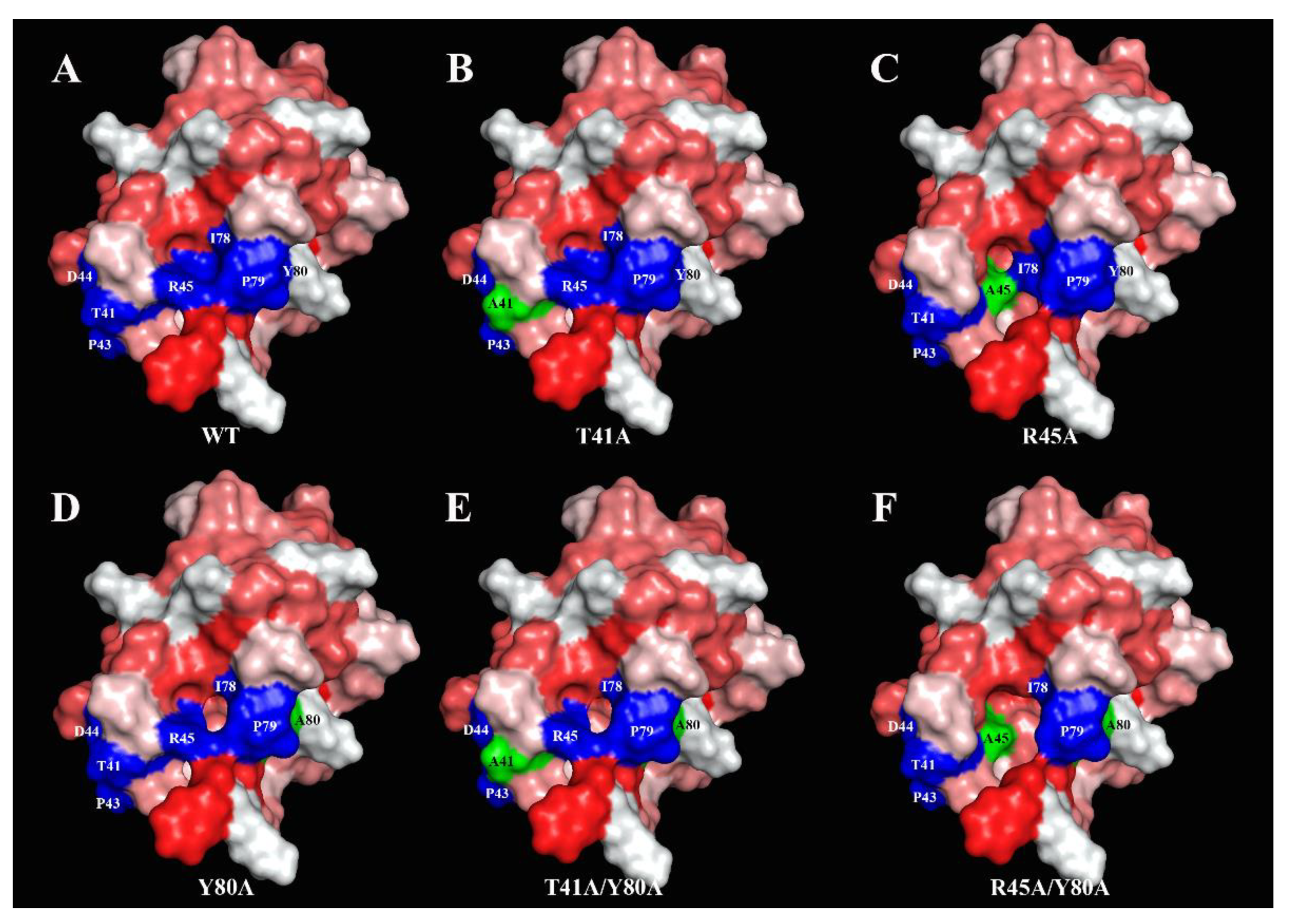
In this work, on the example of lentil Len c 3, we investigated the effect of point amino acid substitutions simultaneously on the structural and allergenic properties of allergen of LTP class as well as on its ligand-binding capacity. For this aim in view, we obtained mutant analogues of the protein with single or double substitutions of three amino acid residues—Thr41, Arg45 and Tyr80 by site-directed mutagenesis. These amino acid residues were selected for several reasons. First, these residues are close to each other and located near the “bottom” entrance to the hydrophobic cavity of Len c 3 (
Figure 3). Second, all of them are included in a presumable conformational epitope of Len c 3 predicted by analogy with Pru p 3 [
15] (
Figure 3). Third, Arg45 and Tyr80 play an important role in the interaction with lipids and stabilization of protein–ligand complexes in various LTPs [
27], including Len c 3 [
18]. Fourth, Tyr80 is one of the key residues for duodenal degradation of plant LTPs [
28], including Len c 3 [
13]. To meet this goal, we investigated structural properties of mutant analogues, their ability to bind lipid ligands and a rate of degradation under conditions mimicking digestion in the human gastrointestinal tract, as well as IgE-binding capacity.
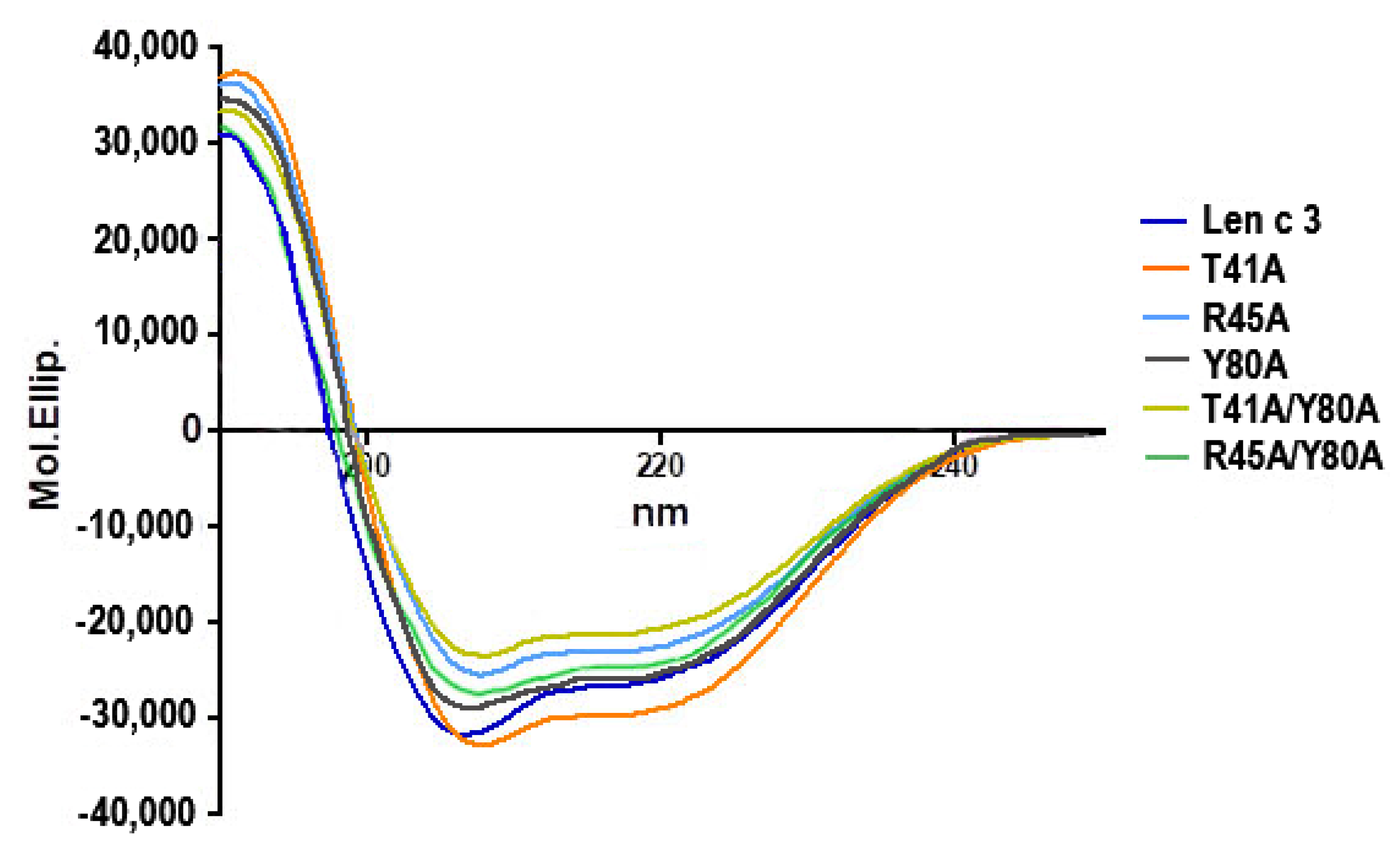
Five mutant analogues of Len c 3 (T41A, R45A, Y80A, T41A/Y80A, R45A/Y80A) were obtained by site-directed mutagenesis and structurally characterized using CD-spectroscopy and computer modeling. The CD spectra revealed that all the mutant analogues did not have significant changes in secondary structures compared to the wild-type protein (
Figure 1). Computer visualization of the mutant analogues demonstrated that some amino acid substitutions substantially affected the volume of the protein hydrophobic cavity. Substitution of Tyr80 (but not Thr41 and Arg45) by alanine led to an increase in the volume of the hydrophobic cavity. The Y80A, T41A/Y80A, and R45A/Y80A analogues had approximately the same sizes of hydrophobic cavities, which were extended compared to Len c 3. In addition, visualization showed that substitutions of selected amino acid residues affected the size and shape of the “bottom” entrance to the hydrophobic cavity. It was also noted that the replacement of Arg45, and to a greater extent of Thr41, but not Tyr80, led to the disruption of continuity of possible conformational IgE-binding epitope of Len c 3 (
Figure 3). Based on these data, we hypothesized that substitutions of all the three amino acids could affect the ability of Len c 3 to bind lipid ligands. At the same time, Thr41 and Arg45 substitutions can alter an ability of the allergen to bind IgE from sera of allergenic patients. Verification of these assumptions was performed by comparative studies of lipid binding in vitro using fluorescence spectroscopy and immunological properties using enzyme-linked immunosorbent assay.
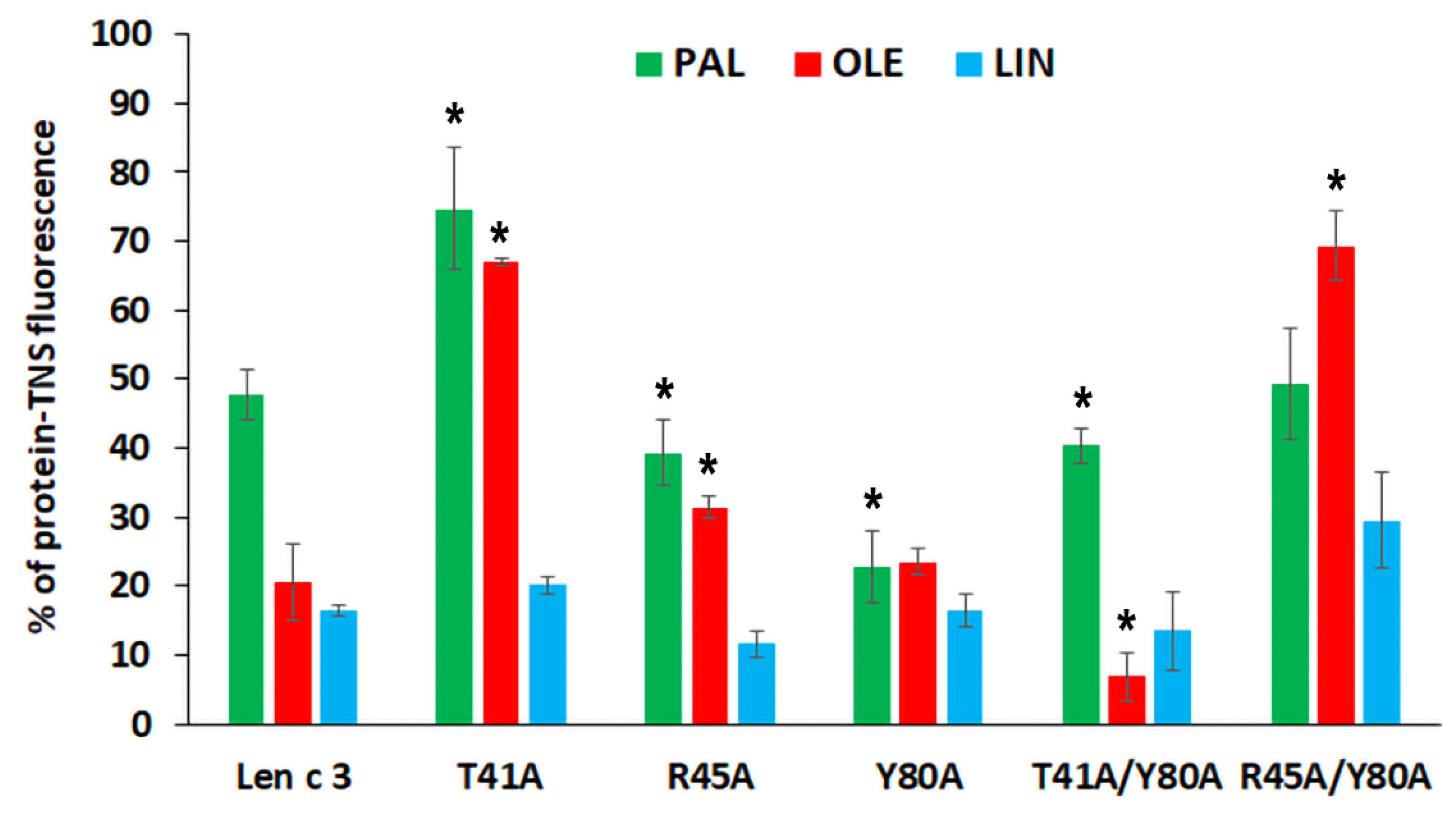
In vitro TNS displacement assay with fatty acids of different chain length and degree of saturation (PAL, OLE, and LIN) demonstrated that the replacement of both Arg45 and Tyr80 did not strongly affect an ability of the proteins to bind FAs. The mutant analogue with double substitution R45A/Y80A bound all these FAs less effectively than Len c 3 (
Figure 2). The results obtained in this work are in good agreement with our previous data on the binding of lauric and stearic acids. An unexpected result was obtained with the mutant analogue T41A, which bound the tested FAs less effectively than Len c 3. Thr41 has not been previously described as an amino acid residue playing an important role in lipid binding. This effect was not observed for the T41A/Y80A analogue. A possible reason for the observed decrease or increase in the ability to bind fatty acids by mutant analogues may be confluence of the following factors: (1) a change in the size and shape of the hydrophobic cavity, which can lead both to spatially more favorable placement of the ligand in the hydrophobic cavity and to constriction in the interactions holding it inside; (2) a change in the size and shape of the “bottom” entrance to the cavity, which may facilitate or prevent the penetration of structurally different ligands into the cavity; (3) changes in the distribution of charges at the “bottom” entrance to the cavity, which play an important role in initiating interactions with ligands.
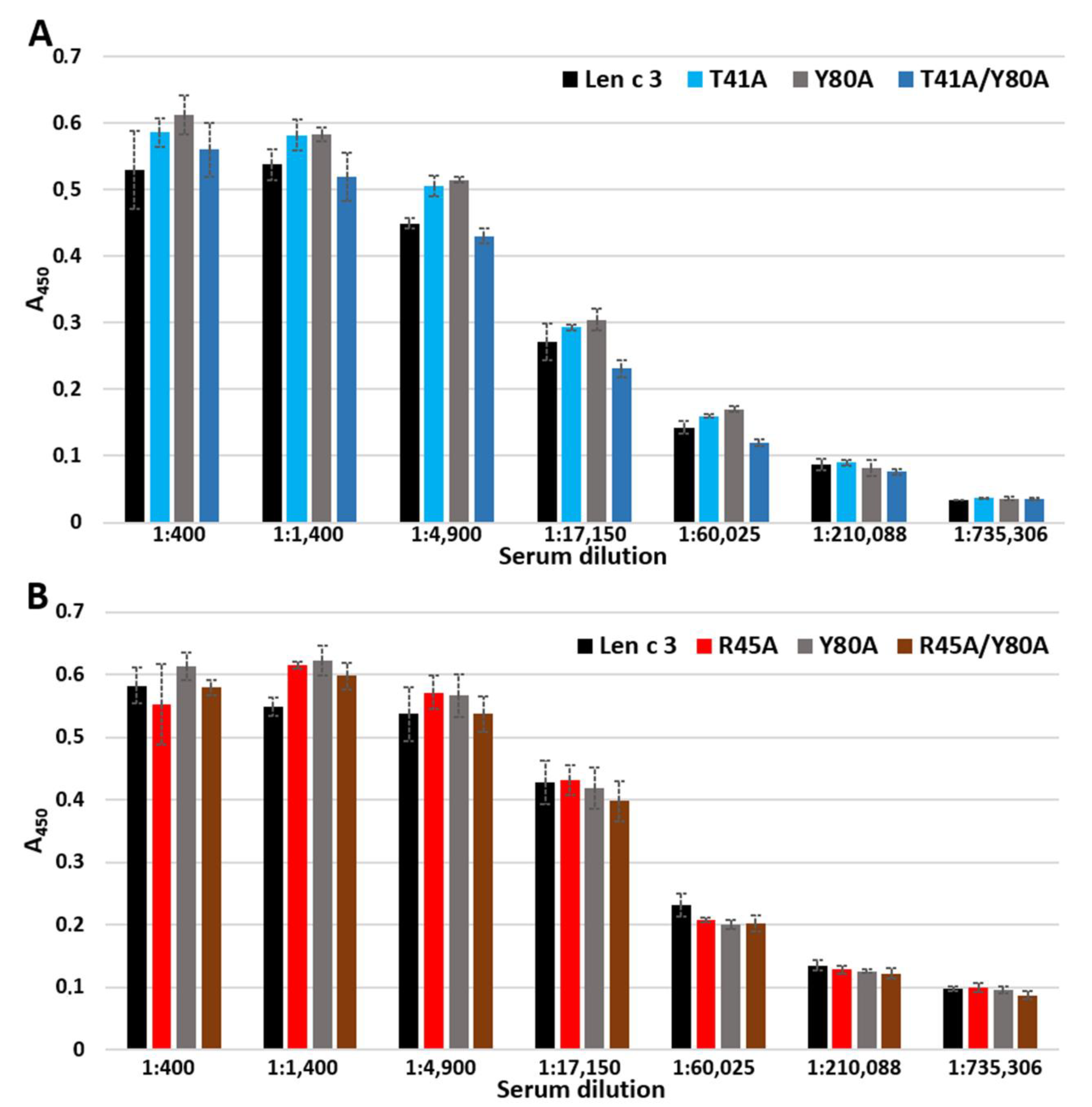
In ELISA experiments, changes in the ability of mutant analogues to bind polyclonal rabbit anti-Len c 3 IgG antibodies were not observed (
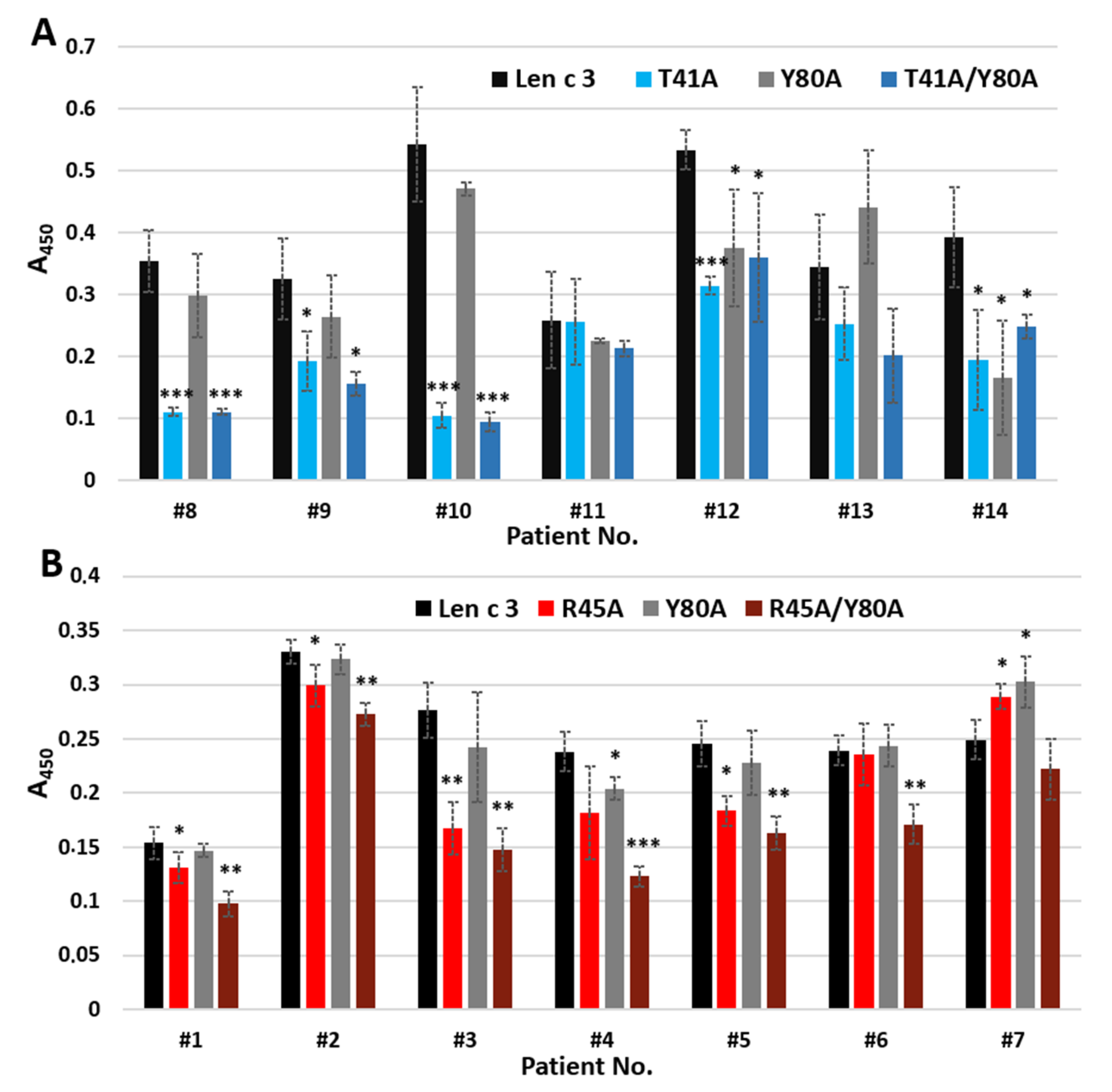
Figure 5). However, when using sera from allergic patients containing specific IgE to Len c 3, we found that substitution of Arg45 and especially Thr41, but not Tyr80, by alanine resulted in a decrease in the IgE-binding capacity of mutant analogues (
Figure 6). The IgE-reactivity of the T41A and T41A/Y80A analogues was similarly reduced. At the same time, the ability of R45A/Y80A to bind sIgE was reduced to a greater extent than in the case of the R45A analogue. We assumed that Thr41 plays the most important role in the interaction with sIgE among the three substituted amino acid residues apparently included in the conformational epitope of Len c 3. At the same time, amino acid substitutions leading to a significant, as in the case of the R45A/Y80A analogue, disruption of the structure of this epitope, can also crucially affect the IgE-binding capacity and the allergenic potential of this protein.
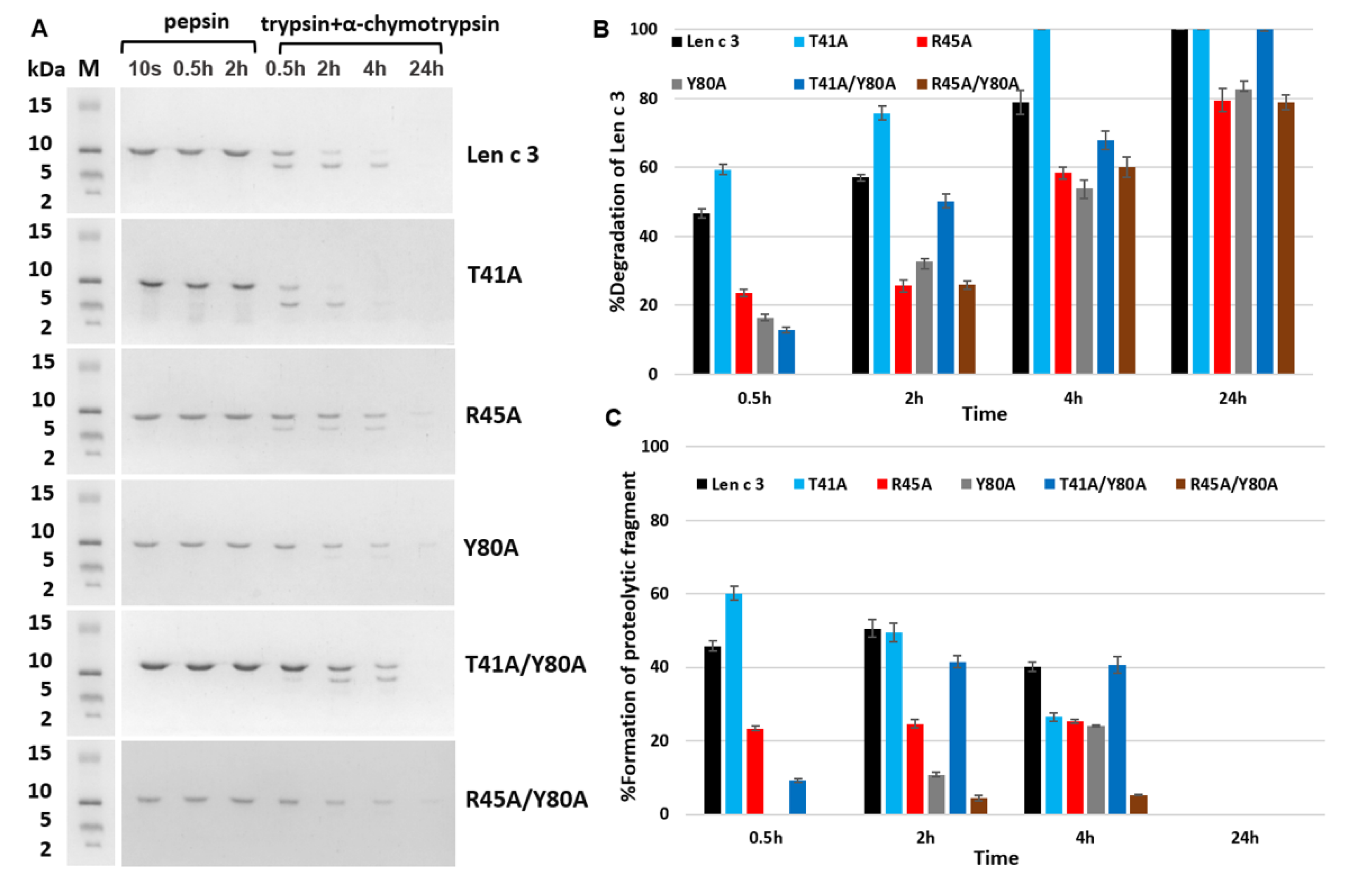
In the last part of the work, we investigated the effects of selected amino acid substitutions on the degradation rate of Len c 3 in experiments simulating its digestion in the human gastrointestinal tract in vitro. For the R45A and Y80A, the expected decrease in the rate of duodenal protein degradation was observed, since both Arg45 and Tyr80 form the cleavage recognition sites for such serine proteases as trypsin and α-chymotrypsin, respectively. At the same time, a cleavage rate of the R45A/Y80A analogue was not further reduced and was approximately the same as for the R45A and Y80A analogues. We hypothesized that a possible reason for this could be a significant change in the compactness of the structure of the R45A/Y80A analogue. This was in good agreement with data obtained by computer modeling of the largest cavity size for this mutant analogue. An unpredictable result was obtained for the T41A mutant analogue that was digested faster than wild-type protein. At the same time, cleavage of the double-mutant analogue T41A/Y80A was slower than that of Len c 3, but faster than cleavage of the Y80A analogue. We suggested that the substitution of Thr41 led to significant changes in the allergen conformation and an increase in the availability of cleavage sites for digesting enzymes. These results for Thr41 were in good agreement with the data on the decrease in the ability of the T41A analogue to bind lipids, as well as sIgE from sera of allergic patients.
5. Conclusions
In this study, using lentil Len c 3 and its mutant analogues (T41A, R45A, Y80A, T41A/Y80A and R45A/Y80A), we investigated the effects of substitutions of three amino acids—Thr41, Arg45 and Tyr80, spaced close to each other and located at the “bottom” entrance to the hydrophobic cavity of the protein, on its structural and allergenic properties as well as on its ligand-binding capacity. We showed that substitutions of three selected amino acids significantly affected the allergen structural organization and protein packing density, the size and shape of the hydrophobic cavity and the “bottom” entrance to it, as well as the structure of a possible conformational IgE-binding epitope. This, in turn, led to a change in the ability of Len c 3 to bind various ligands, in a rate of its proteolytic cleavage, and in the IgE-binding capacity of the allergen. Altogether, this led to a change in the allergenic potential of the protein. Taking into account all the data obtained, we conclude that site-directed mutagenesis of the region, participating in interactions of plant LTPs both with lipids and IgE, may in the future become a promising approach for modeling the structural and biological properties of allergens of this class and creating their hypoallergenic analogues.


 ,
, 


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}





