
Surface Properties of Synaptosomes in the Presence of L-Glutamic and Kainic Acids: In Vitro Alteration of the ATPase and Acetylcholinesterase Activities


Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Isolation of Synaptosomes
2.3. Determination of Acetylcholinesterase Activity
2.4. Determination of the Enzymatic Activity of Brain ATPases
2.5. Microelectrophoresis
2.6. Preparation of Giant Unilamellar Vesicles (GUVs)
2.7. Thermal Shape Fluctuation Analysis of GUVs
2.8. Statistical Analysis
3. Results
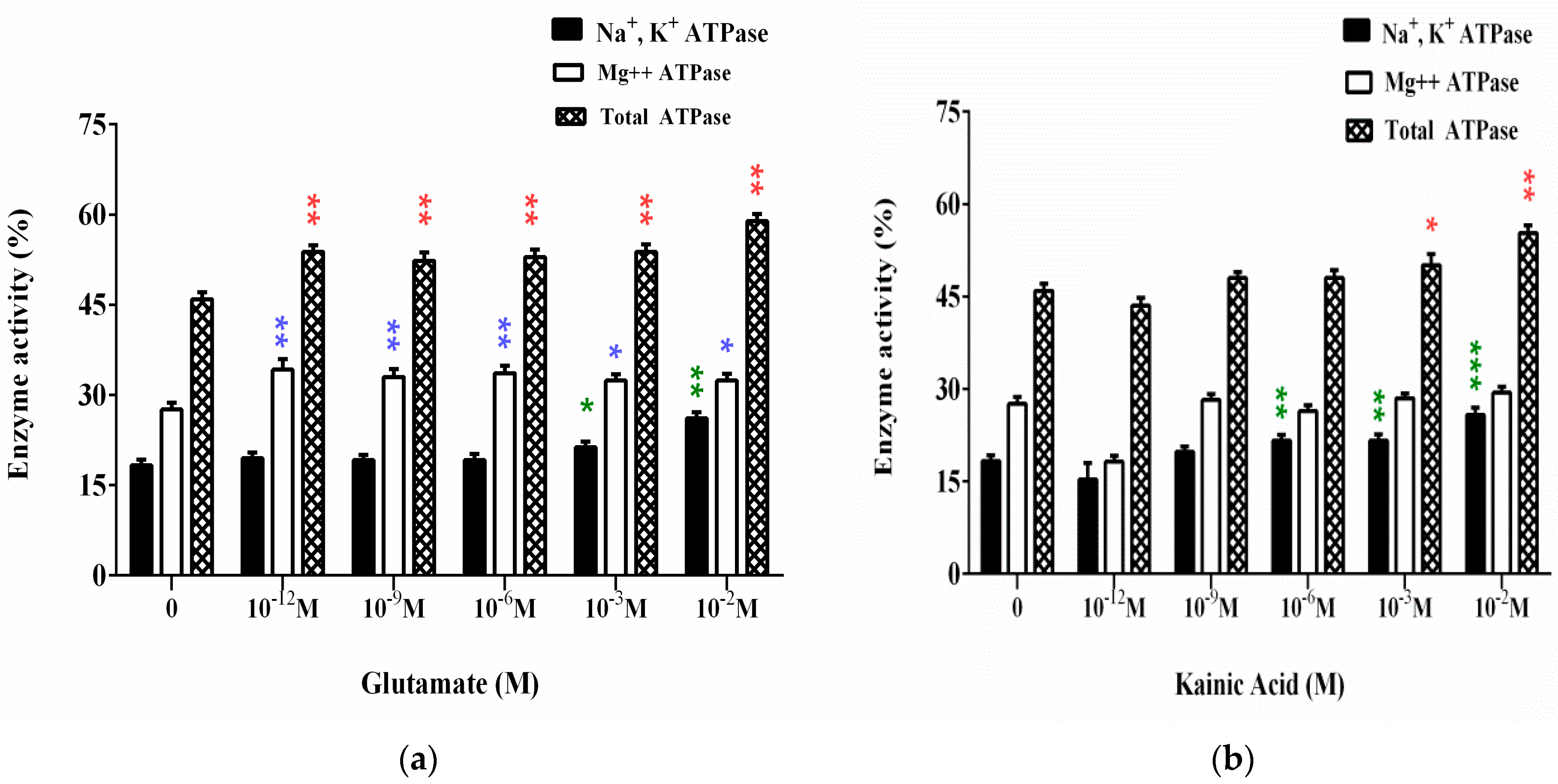
3.1. Effect of L-Glutamate and Kainic Acid on the Adenosine Triphosphatase Activities on Synaptic Membranes (ATPase Assay)
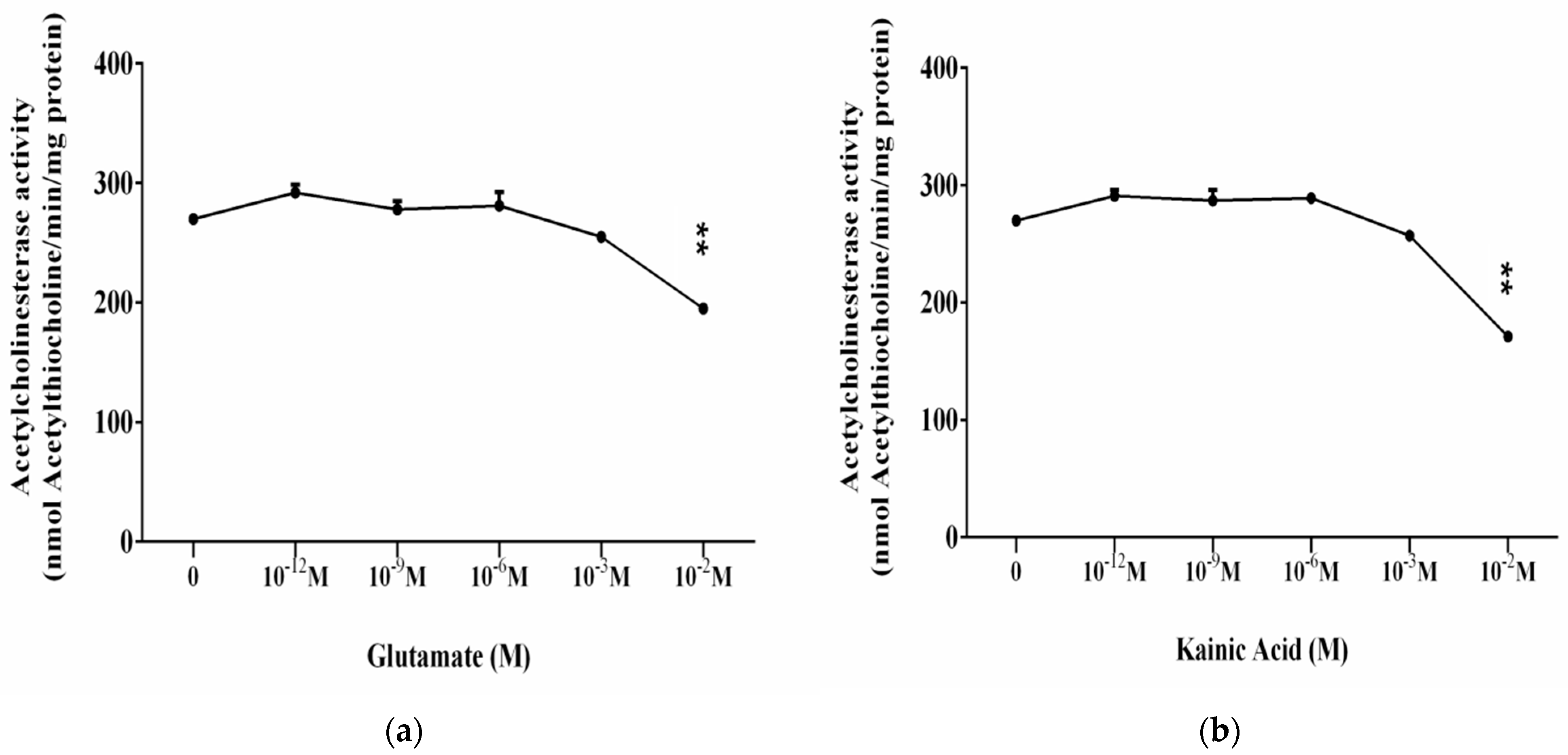
3.2. Influence of L-Glutamate and Kainic Acid on the Enzymatic Activity of Brain Acetylcholinesterase (AChE)
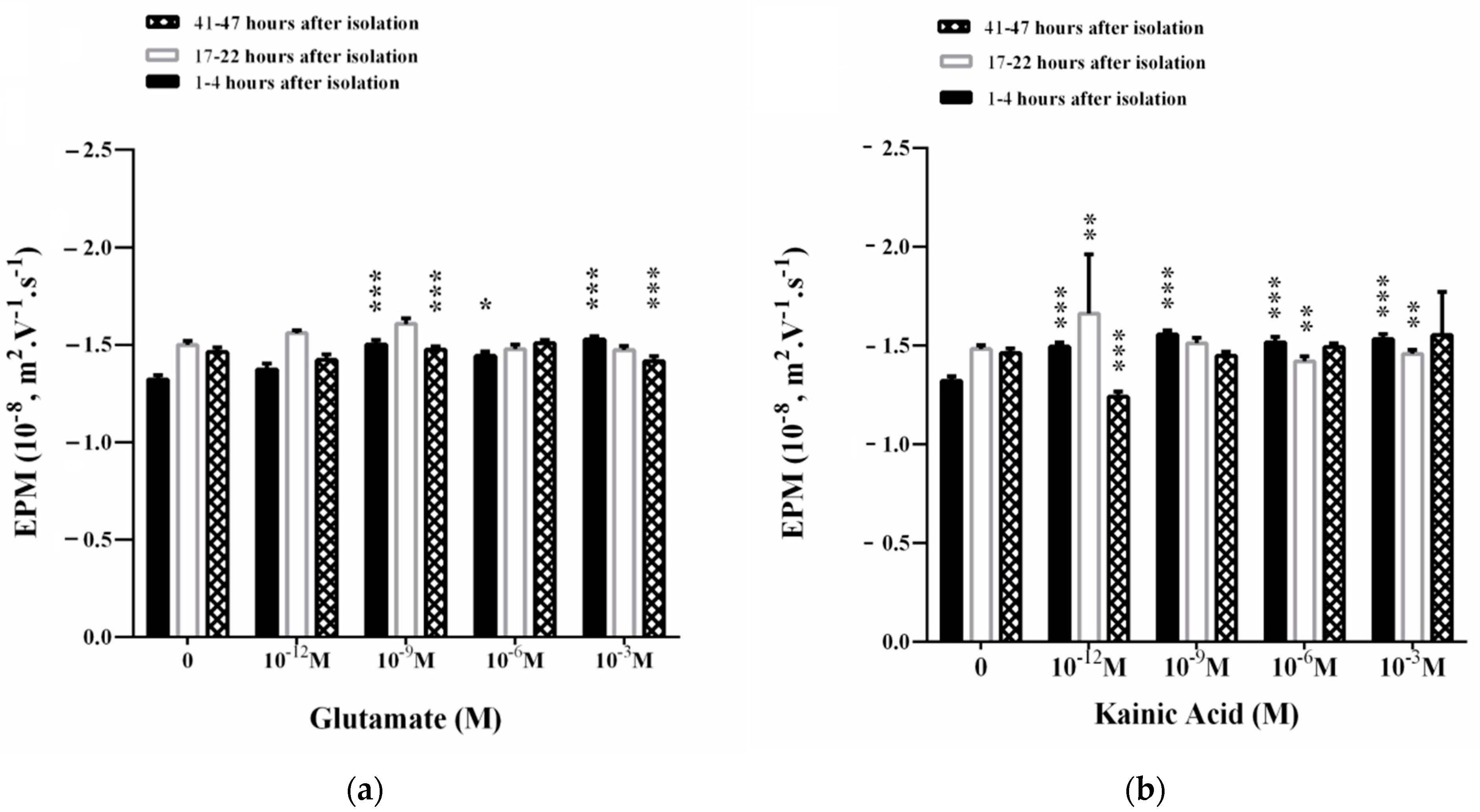
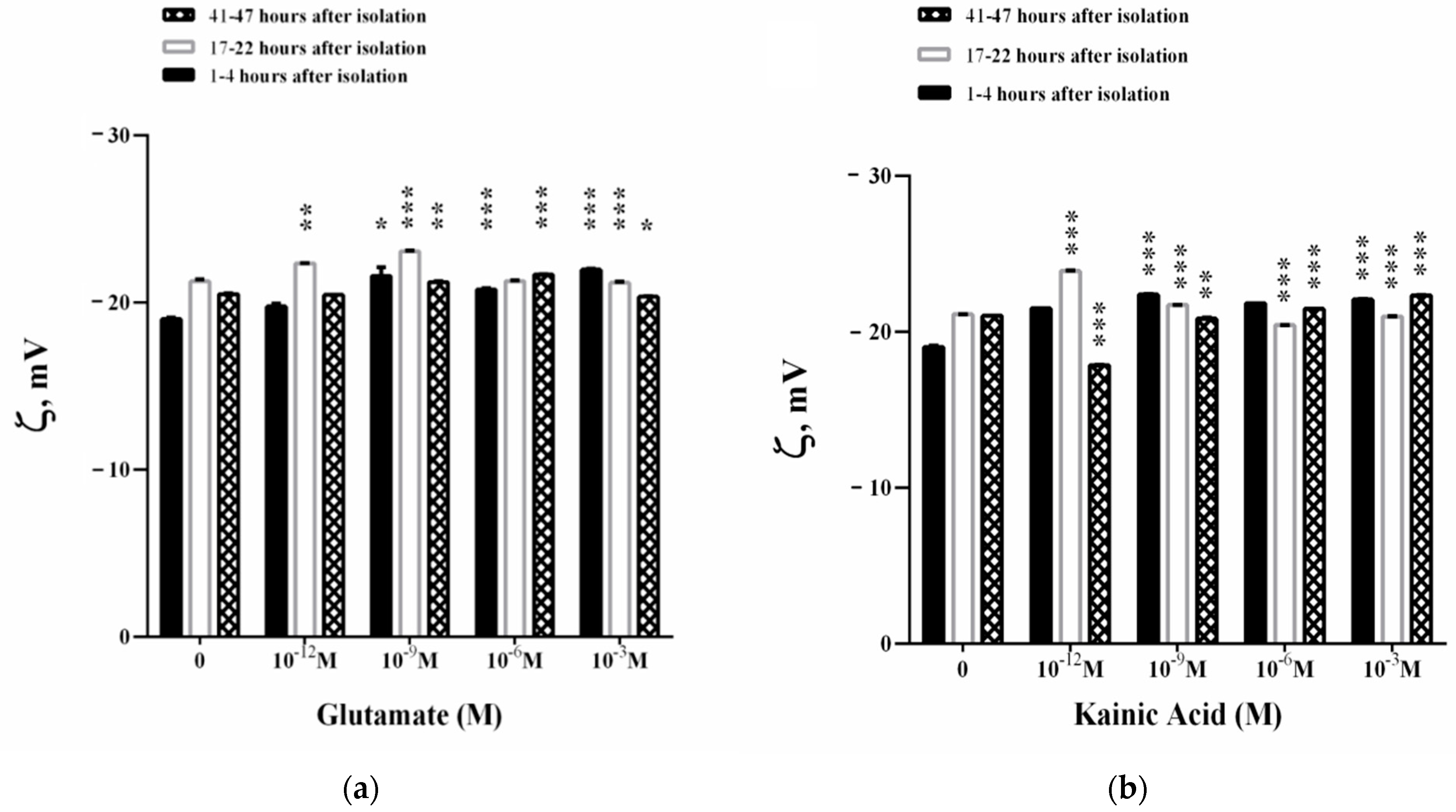
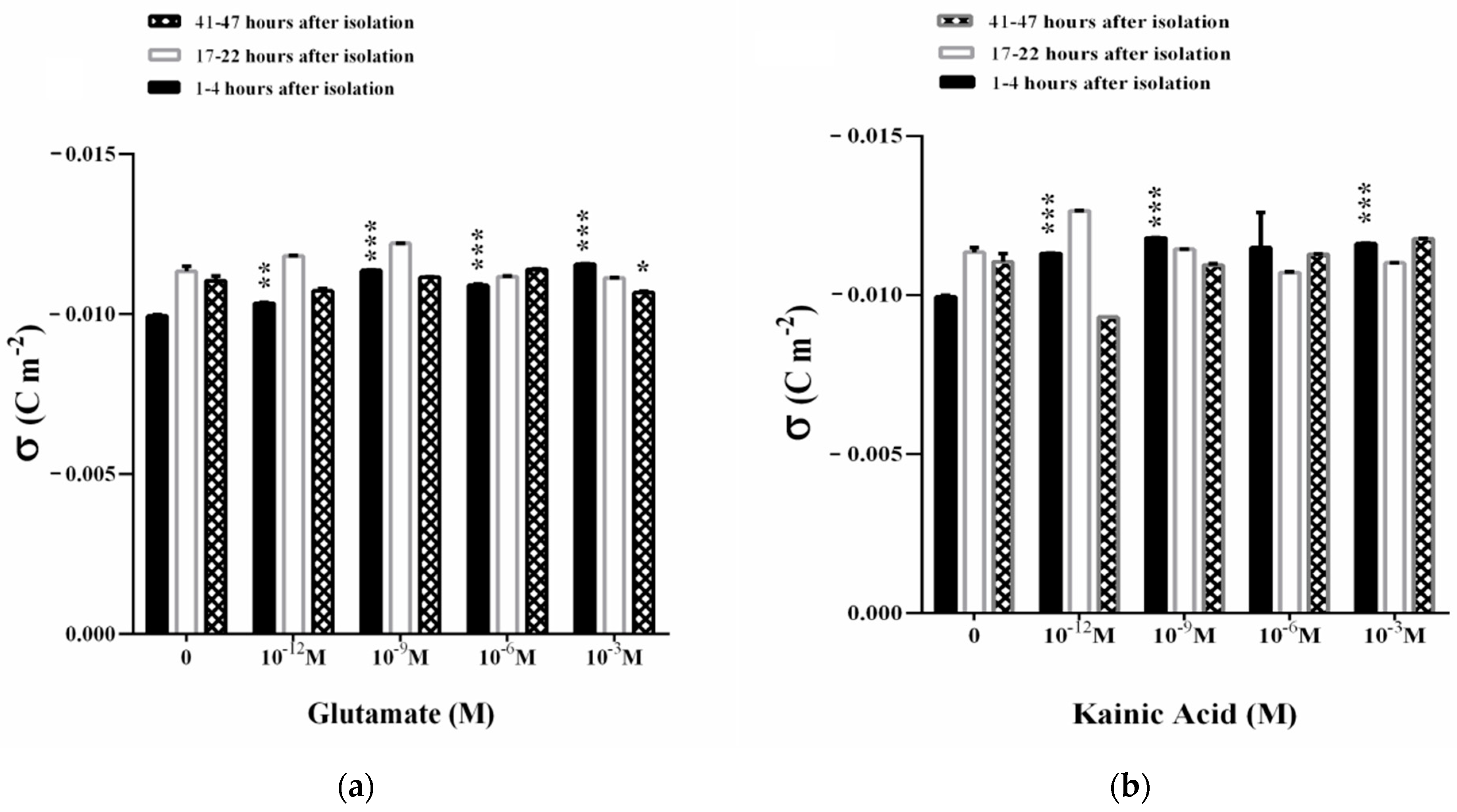
3.3. Influence of L-Glutamate and Kainic Acid on the Electrokinetic Parameters of Synaptosomes
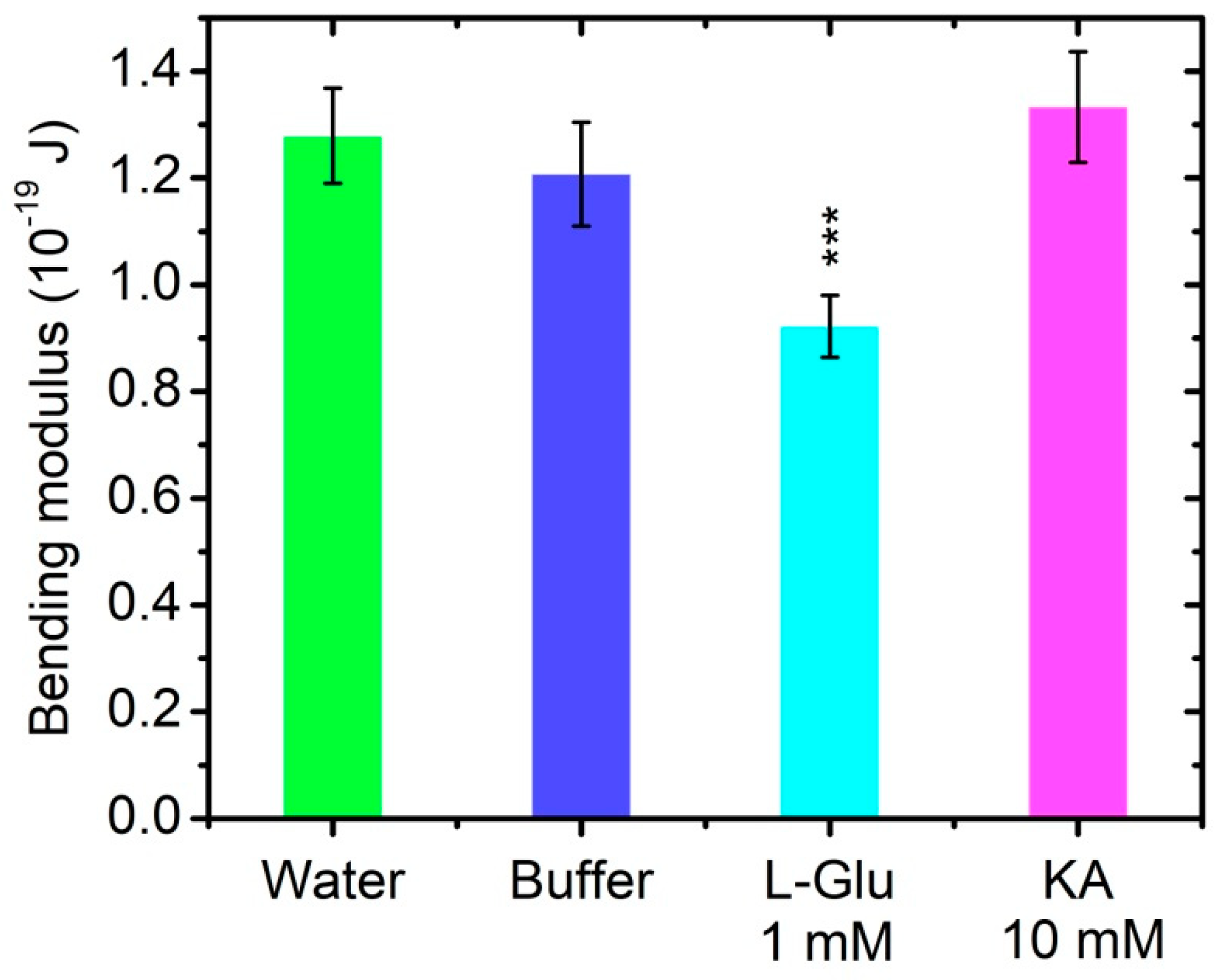
3.4. Bending Elasticity of Model Lipid Membranes in the Presence of L-Glutamate and Kainic Acid
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| AChE | acetylcholinesterase |
| CNS | central nervous system |
| GUV | giant unilamellar vesicles |
| EPM | electrophoretic mobility |
| ITO | indium thin oxide |
| KA | kainic acid |
| L-Glu | L-glutamic acid (L-glutamate) |
| Mg2+-ATPase | magnesium adenosine 5′-triphosphatase |
| NT | neurotransmitter |
| Na+,K+-ATPase | Sodium, potassium-adenosine 5′-triphosphatase |
| PDMS | polydimethylsiloxane |
| POPC | 1-palmitoyl-2-oleoyl-sn-glycero-3-phosphocholine |
| TCA | Trichloroacetic acid |
| Tris-HCl | Trizma Hydrochloride, Tris [hydroxymethyl] aminomethan hydrochloride |
| SD | standard deviation |
| SE | standard error |
| SM | supplementary material |
| σ | surface electrical charge (surface charge density) |
| TSFA | thermal shape fluctuation analysis |
| ζ | zeta (electrokinetic) potential |
References
- Pereda, A.E. Neurobiology: All synapses are created equal. Curr. Biol. 2015, 25, R38–R41. [Google Scholar] [CrossRef] [Green Version]
- Luo, L. Signaling across synapses. In Principles in Neurobiology, 2nd ed.; CRC Press: Boca Raton, FL, USA, 2020; pp. 69–120. ISBN 9780815346050. [Google Scholar] [CrossRef]
- Erecińska, M.; Nelson, D.; Silver, I.A. Metabolic and energetic properties of isolated nerve ending particles (Synaptosomes). Biochim. Biophys. Acta 1996, 127, 13–34. [Google Scholar] [CrossRef] [Green Version]
- Tauskela, J.S.; Fang, H.; Hewitt, M.; Brunette, E.; Ahuja, T.; Thivierge, J.-P.; Comas, T.; Mealing, G.A.R. Elevated synaptic activity preconditions neurons against an in vitro model of Ischemia. J. Biol. Chem. 2008, 283, 34667–34676. [Google Scholar] [CrossRef] [Green Version]
- Hübel, N.; Hosseini-Zare, M.S.; Žiburkus, J.; Ullah, G. The role of glutamate in neuronal ion homeostasis: A case study of spreading depolarization. PLoS Comput. Biol. 2017, 13, e1005804. [Google Scholar] [CrossRef] [Green Version]
- Zou, S.; Chisholm, R.; Tauskela, J.S.; Mealing, G.A.; Johnston, L.J.; Morris, C.E. Force spectroscopy measurements show that cortical neurons exposed to excitotoxic agonists stiffen before showing evidence of bleb damage. PLoS ONE 2013, 8, e73499. [Google Scholar] [CrossRef] [Green Version]
- Kretschmer, B.D.; Schmidt, W.J.; Kostrzewa, R.M.; Herrera-Marschitz, M. Amino acids in neurobiology: Neuroprotective and neurotoxic aspects of amino acids involved in neurotransmission and neuromodulation—General Introduction. Amino Acids 2002, 23, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Whittaker, V.P. The synaptosome. In Handbook of Neurochemistry, 2nd ed.; Lajtha, A., Ed.; Plenum: New York, NY, USA, 1984; Volume 2, pp. 327–364. [Google Scholar]
- Evans, G.J.O. The synaptosome as a model system for studying synaptic physiology. Cold Spring Harb. Protoc. 2015, 5, 421–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arvízu-Santamaría, A.G.; Navarro, R.E.; Soberanes, Y.; Velázquez, E.F.; Santacruz, H.; Inoue, M. Complexation of neurotransmitters—Dopamine, serotonin and melatonin—With a DTPA-based cyclophane of high rigidity: 1H NMR shift and line-broadening. Supramol. Chem. 2017, 29, 658–667. [Google Scholar] [CrossRef]
- Doltchinkova, V.; Shkodrova, M.; Mouleshkova, N.; Stoylov, S. Effect of L-Glutamic acid, gamma-aminobutiric acid and kainic acid on the surface electrical charge and enzyme activity of rat brain synaptosomes. In 10th Jubilee International Conference of the Balkan Physical Union, Proceedings of the AIP Conference, Sofia, Bulgaria, 26–30 August 2018; AIP Publishing: College Park, MA, USA, 2019; Volume 2075, pp. 170022-1–170022-4. [Google Scholar] [CrossRef]
- Galassi, V.V.; Wilke, N. On the coupling between mechanical properties and electrostatics in biological membranes. Membranes 2021, 11, 478. [Google Scholar] [CrossRef] [PubMed]
- Pontes, Y.; Ayala, Y.; Fonseca, A.C.C.; Romão, L.F.; Amaral, R.F.; Salgalo, L.T.; Lima, F.R.; Farina, M.; Vianna, N.B.; Moura-Neto, V.; et al. Membrane elastic properties and cell function. PLoS ONE 2013, 8, e67708. [Google Scholar] [CrossRef] [Green Version]
- Vianallo, A.; Dell’Antone, P.; Macri, F. ATP-ionophore-induced proton translocation in pea stem microsomal vesicles. Biochim. Biophys. Acta 1982, 689, 89–96. [Google Scholar] [CrossRef]
- Rezin, G.T.; Scaini, G.; Gonçalves, C.L.; Ferreira, G.K.; Cardoso, M.R.; Ferreira, A.G.K.; Cunha, M.J.; Schmitz, F.; Varela, R.B.; Quevedo, J.; et al. Evaluation of Na+,K+-ATPase activity in the brain of young rats after acute administration of fenproporex. Braz. J. Psychiatry 2014, 36, 138–142. [Google Scholar] [CrossRef] [Green Version]
- De Lores Arnaiz, G.R.; López Ordieres, M.G. Brain Na+,K+-ATPase activity in aging and disease. Int. J. Biomed. Sci. 2014, 10, 85–102. [Google Scholar]
- Thang, A.; Khandhar, P.B. Physiology, acetylcholinesterase. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2021. Available online: https://www.ncbi.nlm.nih.gov/books/NBK539735/ (accessed on 12 November 2021).
- Vukova, T.; Atanassov, A.; Ivanov, R.; Radicheva, N. Intensity-dependent effects of microwave electromagnetic fields on acetylcholinesterase activity and protein conformation in frog skeletal muscles. Med. Sci. Monit. Int. Med. J. Exp. Clin. Res. 2005, 11, BR50–BR56. [Google Scholar]
- Institute of Medicine (US). Forum on Neuroscience and Nervous System Disorders. 1. Introduction. In Glutamate-Related Biomarkers in Drug Development for Disorders of the Nervous System; Workshop Summary; National Academic Press: Washington, DC, USA, 2011. [Google Scholar]
- Slater, C.R. Neuromuscular Junction (NMJ): Postsynaptic Events in Neuromuscular Transmission. Encycl. Neurosci. 2009, 601–609. [Google Scholar] [CrossRef]
- McKay, M.J.; Marr, K.A.; Price, J.; Greathouse, D.V.; Koeppe, R.E., II. Light-dependent titration of glutamic acid at a bilayer membrane. ACS Omega 2021, 6, 8488–8494. [Google Scholar] [CrossRef] [PubMed]
- Achilonu, S.; Fannucchi, S.; Cross, M.; Fernandes, M.; Dirr, H.W. Role of individual histidines in the ph-dependent global stability of human chloride intracellular Channel 1. Biochemistry 2012, 51, 995–1004. [Google Scholar] [CrossRef]
- Wang, Y.; Fathali, H.; Mishra, D.; Olsson, T.; Keighron, J.D.; Skibicka, K.P.; Cans, A.-S. Counting the number of glutamate molecules in single synaptic vesicles. J. Am. Chem. Soc. 2019, 141, 17507–17511. [Google Scholar] [CrossRef]
- Lakshmann, J.; Padmanaban, G. Effect of some “strong” excitants of central neurons on the uptake of L-glutamate and L-aspartate by synaptosomes. Biochem. Biophys. Res. Comm. 1974, 58, 690–698. [Google Scholar] [CrossRef]
- Johnston, J.A.; Kennedy, S.M.; Twitchin, B. Action of the neurotoxin kainic acid on high affinity uptake of L-glutamic acid in rat brain slices. J. Neurochem. 1979, 32, 121–127. [Google Scholar] [CrossRef]
- Sairazi, N.S.M.; Sirajudeen, K.N.S.; Asari, M.A.; Muzaimi, M.; Mummedy, S.; Sulaiman, S.A. Evidence-based complementary and alternative medicine. Evid. Based Complement.Altern. Med. 2015, 12, 972623. [Google Scholar]
- Tzvetanova, E.; Pavlova, A.; Alexandrova, A.; Nenkova, G.; Petrov, L.; Kirkova, M.; Girchev, R.; Naydenova, E. Are nociceptin (1-13)NH2 and its structural analogue [ORN9]nociceptin(1-13)NH2 able to affect brain antioxidant status in control and kainic acid-treated rats? Cell Biochem. Funct. 2009, 27, 243–250. [Google Scholar] [CrossRef]
- Bossman, H.B.; Carlson, W. Identification of sialic acid at the nerve ending periphery and electrophoretic mobility of isolated synaptosomes. Exp. Cell Res. 1972, 72, 436–440. [Google Scholar] [CrossRef]
- Stoylova, S.S.; Kantcheva, M.R. Microelectrophoretic properties of rat brain membrane structures-synaptosomes. Studia Biophys. 1986, 113, 27–30. [Google Scholar]
- McLaughlin, S. Salicylates and phospholipid bilayer membranes. Nature 1973, 243, 234–236. [Google Scholar] [CrossRef] [PubMed]
- Mironov, S.L.; Dolgaya, E.V. Surface charge of mammalian neurones as revealed by microelectrophoresis. J. Membr. Biol. 1985, 86, 197–202. [Google Scholar] [CrossRef] [PubMed]
- Binotti, B.; Jahn, R.; Pérez-Lara, Á. An overview of the synaptic vesicle lipid composition. Arch. Biochem. Biophys. 2021, 709, 108966. [Google Scholar] [CrossRef] [PubMed]
- Takamori, S.; Holt, M.; Stenius, K.; Lemke, E.A.; Gronborg, M.; Riedel, D.; Urlaub, H.; Schenck, S.; Brugger, B.; Ringler, P.; et al. Molecular anatomy of a trafficking organelle. Cell 2006, 127, 831–846. [Google Scholar] [CrossRef] [Green Version]
- Callenberg, K.M. Membrane Bending Is Critical for Assessing the Thermodynamic Stability of Proteins in the Membrane. Ph.D. Thesis, University of Pittsburgh, Pittsburgh, PA, USA, 2013. [Google Scholar]
- Faizi, H.A.; Frey, S.L.; Steinkühler, J.; Dimova, R.; Vlahovska, P.M. Bending rigidity of charged lipid bilayer membranes. Soft Matter 2019, 15, 6006–6013. [Google Scholar] [CrossRef] [Green Version]
- Bivas, I.; Ermakov, Y.A. Elasticity and electrostatics of amphiphilic layers: Current state of the theory and the experiment. In Advances in Planar Lipid bilayers and Liposomes, 1st ed.; Leitmannova Liu, A., Ed.; Academic Press: Cambridge, MA, USA, 2006; Volume 5, pp. 313–343. [Google Scholar] [CrossRef]
- Luisi, P.L.; Walde, P. Giant Vesicles; John Wiley & Sons, Ltd.: Chichester, UK, 2000. [Google Scholar]
- Dimova, R.; Aranda, S.; Bezlyepkina, N.; Nikolov, V.; Riske, K.A.; Lipowsky, R. A practical guide to giant vesicles: Probing the membrane nanoregime via optical microscopy. J. Phys. Condens. Matter 2006, 18, S1151–S1176. [Google Scholar] [CrossRef] [Green Version]
- Bagatolli, L.A. To see or not to see: Lateral organization of biological membranes and fluorescence microscopy. Biochim. Biophys. Acta (BBA)-Biomembr. 2006, 1758, 1541–1556. [Google Scholar] [CrossRef] [Green Version]
- Available online: https://eur-lex.europa.eu/legal-content/BG/TXT/HTML/?uri=CELEX:32010L0063&from=EN (accessed on 12 November 2021).
- Ellman, G.L.; Courtney, D.K.; Andreas, V.; Featherstone, R.M. A new and rapid colorimetric determination of acetylcholinesterase activity. Biochem. Pharmacol. 1961, 7, 88–95. [Google Scholar] [CrossRef]
- Rozhanets, V.V.; Kozlova, V.P.; Rodina, R.I.; Shvets, V.I.; Glebov, R.N. Effects of anticonvulsive substances on Na,K-ATPase from the synaptic membranes of animal brain. Biokhimia SSSR 1978, 43, 892–898, (In Russian with English). [Google Scholar]
- Fieske, C.H.; Subbarow, Y. The colorimetric determination of phosphorus. J. Biol. Chem. 1925, 66, 375–400. [Google Scholar] [CrossRef]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the folin reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef]
- Delgado, A.V.; González-Caballero, F.; Hunter, R.J.; Koopal, K.L.; Lyklema, J. Measurement and interpretation of electrokinetic phenomena. J. Colloid Interface Sci. 2007, 309, 194–224. [Google Scholar] [CrossRef]
- McLaughlin, S. Electrostatic Potentials at membrane-solution interfaces. Curr. Top. Membr. Transp. 1977, 9, 71–144. [Google Scholar]
- Angelova, M.I.; Dimitrov, D.S. Liposome electroformation. Faraday Discuss. Chem. Soc. 1986, 81, 303–311. [Google Scholar] [CrossRef]
- Vitkova, V.; Antonova, K.; Popkirov, G.; Mitov, M.D.; Ermakov, Y.A.; Bivas, I. Electrical resistivity of the liquid phase of vesicular suspensions prepared by different methods. J. Phys. Conf. Ser. 2010, 253, 012059. [Google Scholar] [CrossRef] [Green Version]
- Méléard, P.; Bagatolli, L.A.; Pott, T. Giant unilamellar vesicle electroformation: From lipid mixtures to native membranes under physiological conditions. Methods Enzymol. 2009, 465, 161–176. [Google Scholar] [CrossRef]
- Helfrich, W. Elastic properties of lipid bilayers: Theory and possible experiments. Z. Naturforsch. C 1973, 28, 693–703. [Google Scholar] [CrossRef]
- Evans, E.A. A new material concept for the red cell membrane. Biophys. J. 1973, 13, 926–940. [Google Scholar] [CrossRef] [Green Version]
- Faucon, J.F.; Mitov, M.D.; Méléard, P.; Bivas, I.; Bothorel, P. Bending elasticity and thermal fluctuations of lipid membranes. Theoretical and experimental requirements. J. Physique 1989, 50, 2389–2414. [Google Scholar] [CrossRef] [Green Version]
- Mitov, M.D.; Faucon, J.F.; Méléard, P.; Bothorel, P. Thermal fluctuations of membranes. In Advances in Supramolecular Chemistry; Gokel, G.W., Ed.; JAI Press Inc.: Greenwich, CT, USA, 1992; pp. 93–139. [Google Scholar]
- Vitkova, V.; Misbah, C. Dynamics of lipid vesicles—From thermal fluctuations to rheology. In Advances in Planar Lipid Bilayers and Liposomes; Iglič, A., Ed.; Academic Press: Burlington, MA, USA, 2011; pp. 258–292. [Google Scholar]
- Genova, J.; Vitkova, V.; Aladgem, L.; Meleard, P.; Mitov, M.D. Using stroboscopic illumination to improve the precision of the bending modulus measured by the analysis of thermally induced shape fluctuations of giant vesicles. Bulg. J. Phys. 2004, 31, 68–75. [Google Scholar]
- Genova, J.; Vitkova, V.; Bivas, I. Registration and analysis of the shape fluctuations of nearly spherical lipid vesicles. Phys. Rev. E Stat. Nonlinear Soft Matter Phys. 2013, 88, 022707. [Google Scholar] [CrossRef]
- Milner, S.T.; Safran, S.A. Dynamical fluctuations of droplet microemulsions and vesicles. Phys. Rev. A 1987, 36, 4371–4379. [Google Scholar] [CrossRef]
- Bivas, I.; Hanusse, P.; Bothorel, P.; Lalanne, J.; Aguerre-Chariol, O. An application of the optical microscopy to the determination of the curvature elastic modulus of biological and model membranes. J. Phys. 1987, 48, 855–867. [Google Scholar] [CrossRef]
- Genova, J.; Vitkova, V.; Aladgem, L.; Mitov, M.D. The stroboscopic illumination gives new opportunities and improves the precision of the bending elastic modulus measurement. J. Optoelectron. Adv. Mater. 2005, 7, 257–260. [Google Scholar]
- Maestú, F.; de Haan, W.; Busche, M.A.; DeFelipe, J. Neuronal excitation/inhibition imbalance: Core element of a translational perspective on Alzheimer pathophysiology. Ageing Res. Rev. 2021, 69, 101372. [Google Scholar] [CrossRef]
- Szkudlarek, J.; Lachowicz, L.; Wojtkowiak, R. Effects in vitro of L-Glutamate and Kainic Acid on the ATPase activities of synaptosomal membtanes from different areas of rat brain. Neurosci. Lett. 1986, 65, 304–310. [Google Scholar] [CrossRef]
- Kimelberg, H.K.; Papahadjopoulos, D. Phospholipid requirements for (Na+ + K+)–ATPase activity: Head-group specificity and fatty acid fluidity. Biochim. Biophys. Acta 1972, 282, 277–2926. [Google Scholar] [CrossRef]
- Yoneda, J.S.; Sebinelli, H.G.; Itri, R.; Ciancaglini, P. Overview on solubilization and lipid reconstitution of Na,K-ATPase: Enzyme kinetic and biophysical characterization. Biophys. Rev. 2020, 12, 49–64. [Google Scholar] [CrossRef]
- Jebali, J.; Khedher, S.B.; Sabbagh, M.; Kamel, N.; Banni, M.; Boussetta, H. Cholinesterase activity as biomarker of neurotoxicity: Utility in the assessment of aquatic environmental condition. J. Integr. Coast. Zone Manag. 2013, 13, 525–537. [Google Scholar] [CrossRef] [Green Version]
- Seaman, G.V.F.; Heard, D.H. The surface of a washed human erythrocyte as a polyanion. J. Gen. Physiol. 1960, 44, 251–272. [Google Scholar] [CrossRef] [PubMed]
- Hsu, J.-P.; Huang, S.-W.; Hsieh, T.-S.; Young, T.-H.; Hu, W.-W. Electrophoretic behavior of cerebellar granule neurons. Electrophoresis 2002, 23, 2001–2006. [Google Scholar] [CrossRef]
- Angelova, P.R. Sources and triggers of oxidative damage in neurodegeneration. Free Radic. Biol. Med. 2021, 173, 52–63. [Google Scholar] [CrossRef]
- Konev, S.V. Structural Lability of Biological Membranes and Regulatory Processes; Nauka i Tecknika Publish: Minsk, Belarus, 1987. (In Russian) [Google Scholar]
- Brown, W.H.; Iverson, B.L.; Anslyn, E.V.; Foote, C.S.; Novak, B.N. Organic Chemistry, 8th ed.; Cengage Learning: Boston, MA, USA, 2017; p. 1312. ISBN 10:1-305-58035-4. [Google Scholar]
- Bivas, I.; Méléard, P. Bending elasticity and bending fluctuations of lipid bilayer containing additive. Phys. Rev. E 2003, 67, 012901. [Google Scholar] [CrossRef]
- Pinigin, K.V.; Kuzmin, P.I.; Akimov, S.A.; Galimzyanov, T.R. Additional contributions to elastic energy of lipid membranes: Tilt-curvature coupling and curvature gradient. Phys. Rev. E 2020, 102, 042406. [Google Scholar] [CrossRef] [PubMed]
- Crans, R.A.J.; Dalemans, S.; Raedt, R.; Ciruela, F.; Stove, C.P. Kainic acid-induced Status epilepticus decreases mGlu5 receptor and phase-specifically downregulates Homer1b/c expression. Brain Res. 2020, 1730, 146640. [Google Scholar] [CrossRef]
- Joseph, C.; Tseng, C.Y.; Zocchi, G.; Tlusty, T. Asymmetric effect of mechanical stress on the forward and reverse reaction catalyzed by an enzyme. PLoS ONE 2014, 9, e101442. [Google Scholar] [CrossRef]
- Ma, Y.; Poole, K.; Goyette, J.; Gaus, K. Introducing membrane charge and membrane potential to T cell signaling. Front. Immunol. 2017, 8, 1513. [Google Scholar] [CrossRef] [PubMed]
- Dencheva, E.; Ivanov, R. The Cholinesterases as biomonitoring markers in cases of poisoning of organisms in ecotoxicological environment. In 10th Anniversary Seminar of Ecology with International Participation-2017 Proceedings; IBER BAS: Sofia, Bulgaria, 2017; pp. 125–134. ISBN 979-853-476-132-4. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | NT, mmol/L | Bending Modulus, 10−en J | Number of Vesicles | GF |
|---|---|---|---|---|
| (i) Control, H2O | 0 | 1.28 ± 0.09 | 17 | 0.59 |
| (ii) Control, buffer | 0 | 1.21 ± 0.10 | 8 | 0.31 |
| L-Glutamate | 1 | 0.92 ± 0.06 | 9 | 0.62 |
| Kainic acid | 10 | 1.33 ± 0.10 | 5 | 0.77 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Doltchinkova, V.; Mouleshkova, N.; Vitkova, V. Surface Properties of Synaptosomes in the Presence of L-Glutamic and Kainic Acids: In Vitro Alteration of the ATPase and Acetylcholinesterase Activities. Membranes 2021, 11, 987. https://doi.org/10.3390/membranes11120987
Doltchinkova V, Mouleshkova N, Vitkova V. Surface Properties of Synaptosomes in the Presence of L-Glutamic and Kainic Acids: In Vitro Alteration of the ATPase and Acetylcholinesterase Activities. Membranes. 2021; 11(12):987. https://doi.org/10.3390/membranes11120987
Chicago/Turabian StyleDoltchinkova, Virjinia, Nevena Mouleshkova, and Victoria Vitkova. 2021. "Surface Properties of Synaptosomes in the Presence of L-Glutamic and Kainic Acids: In Vitro Alteration of the ATPase and Acetylcholinesterase Activities" Membranes 11, no. 12: 987. https://doi.org/10.3390/membranes11120987
APA StyleDoltchinkova, V., Mouleshkova, N., & Vitkova, V. (2021). Surface Properties of Synaptosomes in the Presence of L-Glutamic and Kainic Acids: In Vitro Alteration of the ATPase and Acetylcholinesterase Activities. Membranes, 11(12), 987. https://doi.org/10.3390/membranes11120987