
Differential Antimicrobial Effect of Essential Oils and Their Main Components: Insights Based on the Cell Membrane and External Structure

 ,
,  ,
,  ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Extraction of Essential Oils and Characterization
2.2. Antibacterial Activity
2.3. Antifungal Activity
2.4. Antiparasitic Activity
2.5. Statistic Analysis
3. Results
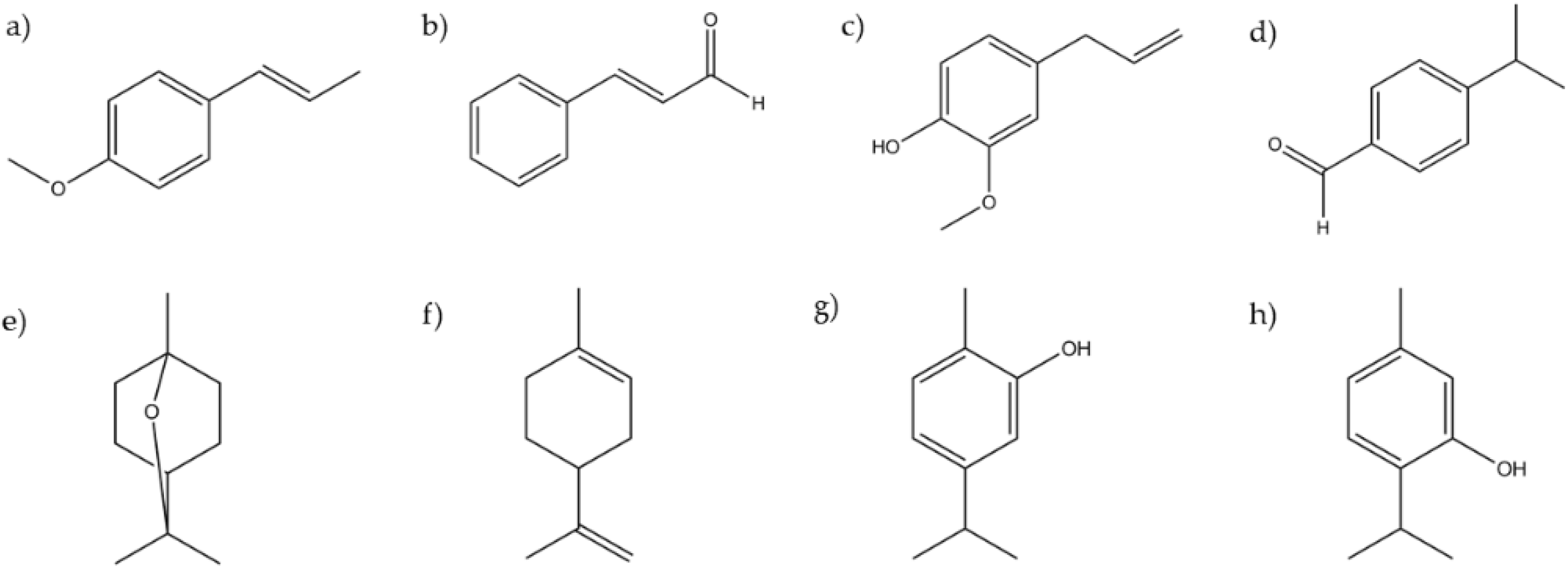
3.1. Essential Oils Characterization
3.2. Antibacterial Activity
3.3. Antifungal Activity
3.4. Antiparasitic Activity
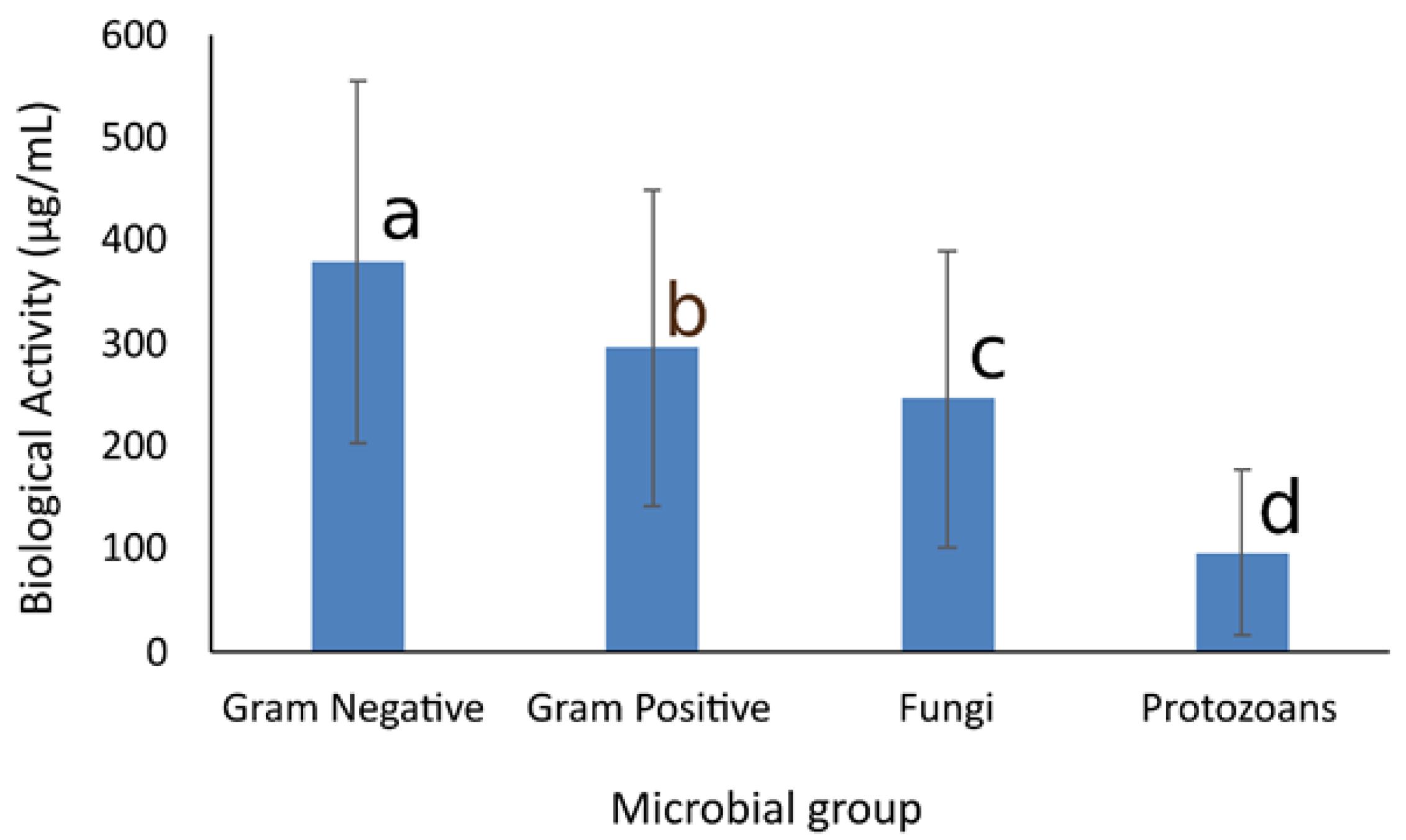
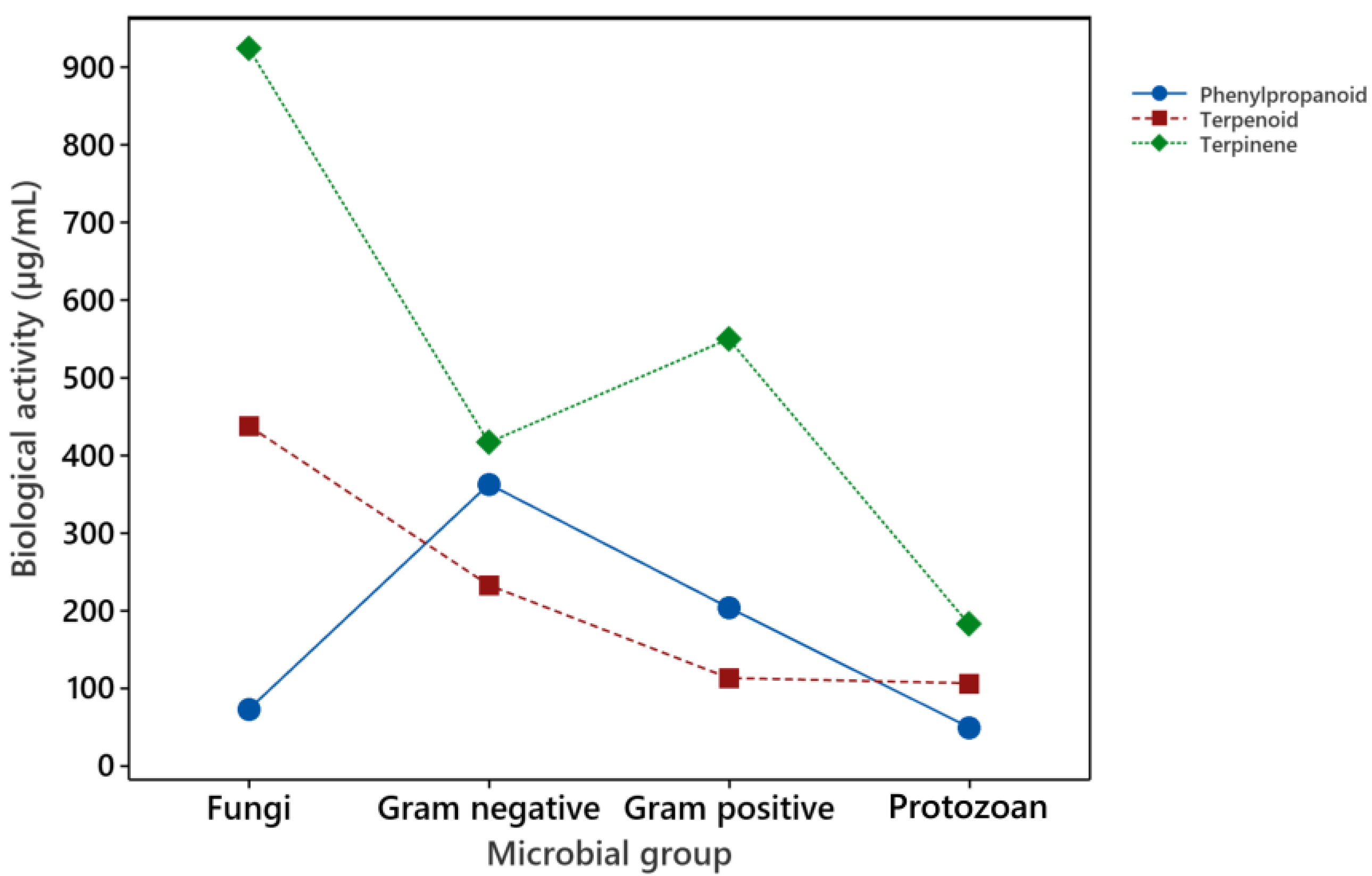
3.5. Analysis of Overall Biological Activity
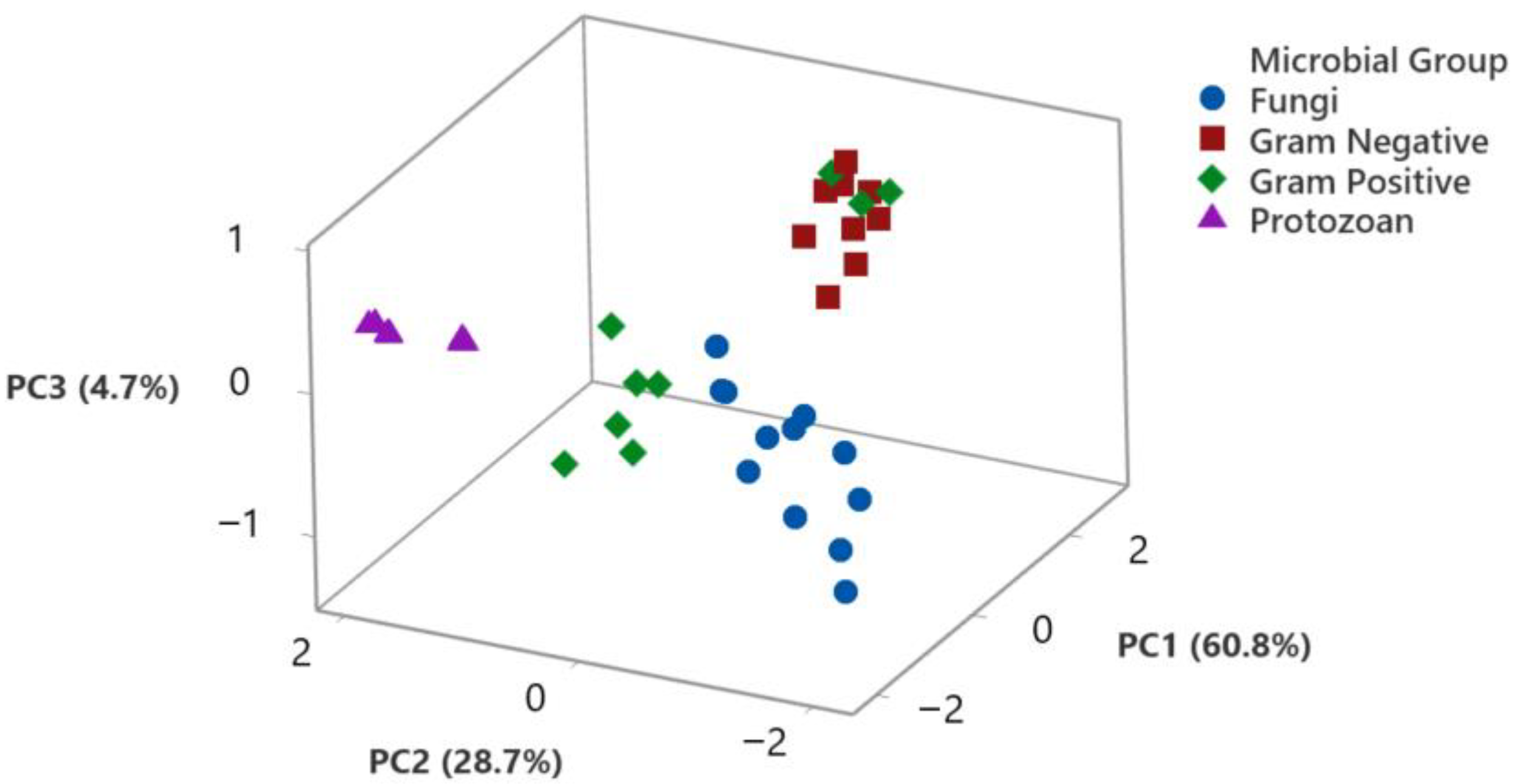
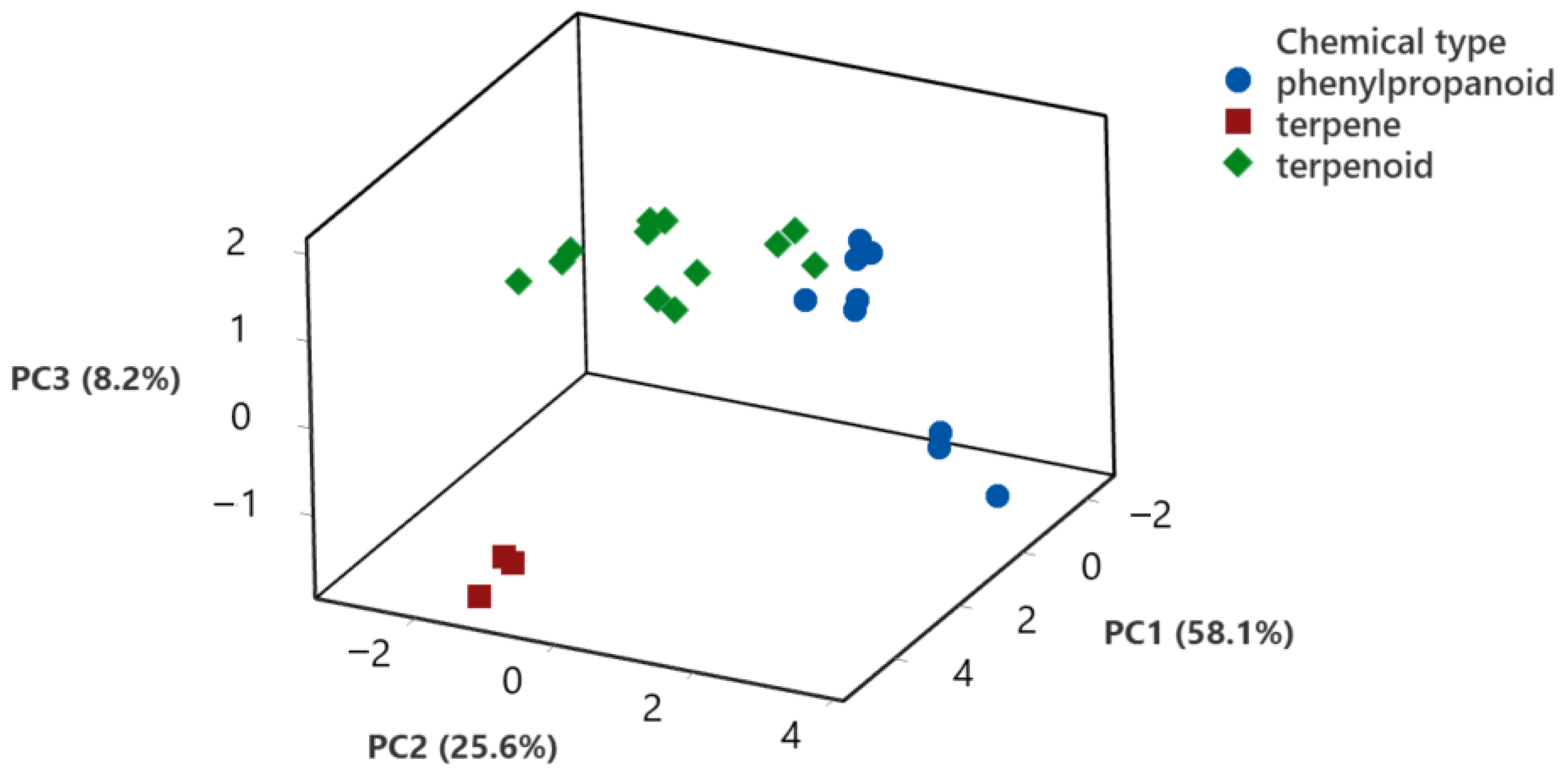
3.6. Principal Component Analysis of the Biological Activity
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lappa, I.K.; Simini, E.; Nychas, G.-J.E.; Panagou, E.Z. In vitro evaluation of essential oils against Aspergillus carbonarius isolates and their effects on Ochratoxin A related gene expression in synthetic grape medium. Food Control. 2017, 73, 71–80. [Google Scholar] [CrossRef]
- Oüzek, G.; Schepetkin, I.A.; Utegenova, G.A.; Kirpotina, L.N.; Andrei, S.R.; Oüzek, T.; Baser, K.H.C.; Abidkulova, K.T.; Kushnarenko, S.V.; Khlebnikov, A.I.; et al. Chemical composition and phagocyte immunomodulatory activity of Ferula iliensis essential oils. J. Leukoc. Biol. 2017, 101, 1361–1371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burt, S. Essential oils: Their antibacterial properties and potential applications in foods—A review. Int. J. Food Microbiol. 2004, 94, 223–253. [Google Scholar] [CrossRef] [PubMed]
- Adame-Gallegos, J.R.; Ochoa, S.A.; Nevarez-Moorillon, G.V. Potential Use of Mexican Oregano Essential Oil against Parasite, Fungal and Bacterial Pathogens. J. Essent. Oil Bear. Plants 2016, 19, 553–567. [Google Scholar] [CrossRef]
- Leyva-López, N.; Gutiérrez-Grijalva, E.P.; Vazquez-Olivo, G.; Heredia, J.B. Essential Oils of Oregano: Biological Activity beyond Their Antimicrobial Properties. Molecules 2017, 22, 989. [Google Scholar] [CrossRef] [Green Version]
- D’Agostino, M.; Tesse, N.; Frippiat, J.P.; Machouart, M.; Debourgogne, A.; Agostino, D. Essential Oils and Their Natural Active Compounds Presenting Antifungal Properties. Molecules 2019, 24, 3713. [Google Scholar] [CrossRef] [Green Version]
- Kong, J.; Zhang, Y.; Ju, J.; Xie, Y.; Guo, Y.; Cheng, Y.; Qian, H.; Quek, S.Y.; Yao, W. Antifungal effects of thymol and salicylic acid on cell membrane and mitochondria of Rhizopus stolonifer and their application in postharvest preservation of tomatoes. Food Chem. 2019, 285, 380–388. [Google Scholar] [CrossRef]
- Xu, Y.; Wei, J.; Wei, Y.; Han, P.; Dai, K.; Zou, X.; Jiang, S.; Xu, F.; Wang, H.; Sun, J.; et al. Tea tree oil controls brown rot in peaches by damaging the cell membrane of Monilinia fructicola. Postharvest Biol. Technol. 2021, 175, 111474. [Google Scholar] [CrossRef]
- André, W.P.P.; Cavalcante, G.S.; Ribeiro, W.L.C.; Dos Santos, J.M.L.; Macedo, I.T.F.; De Paula, H.C.B.; De Morais, S.M.; De Melo, J.V.; Bevilaqua, C.M.L. Anthelmintic effect of thymol and thymol acetate on sheep gastrointestinal nematodes and their toxicity in mice. Rev. Bras. Parasitol. Veterinária 2017, 26, 323–330. [Google Scholar] [CrossRef]
- Andrade-Ochoa, S.; Sánchez-Aldana, D.; Vargas, K.F.C.; Rivera-Chavira, B.E.; Sánchez-Torres, L.E.; Camacho, A.D.; Nogueda-Torres, B.; Nevárez-Moorillón, G.V. Oviposition Deterrent and Larvicidal and Pupaecidal Activity of Seven Essential Oils and their Major Components against Culex quinquefasciatus Say (Diptera: Culicidae): Synergism–antagonism Effects. Insects 2018, 9, 25. [Google Scholar] [CrossRef] [Green Version]
- Tyagi, V.; Patel, R.; Hazarika, H.; Dey, P. Chemical composition and bioefficacy for larvicidal and pupicidal activity of essential oils against two mosquito species. Int. J. Mosq. Res. 2017, 4, 112–118. [Google Scholar]
- Di Pasqua, R.; Betts, G.; Hoskins, N.; Edwards, M.; Ercolini, D.; Mauriello, G. Membrane Toxicity of Antimicrobial Compounds from Essential Oils. J. Agric. Food Chem. 2007, 55, 4863–4870. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-González, L.; Vargas, M.; González-Martínez, C.; Chiralt, A.; Cháfer, M. Use of Essential Oils in Bioactive Edible Coatings: A Review. Food Eng. Rev. 2011, 3, 1–16. [Google Scholar] [CrossRef]
- Trombetta, D.; Castelli, F.; Sarpietro, M.G.; Venuti, V.; Cristani, M.; Daniele, C.; Saija, A.; Mazzanti, G.; Bisignano, G. Mechanisms of Antibacterial Action of Three Monoterpenes. Antimicrob. Agents Chemother. 2005, 49, 2474–2478. [Google Scholar] [CrossRef] [Green Version]
- Van De Vel, E.; Sampers, I.; Raes, K. A review on influencing factors on the minimum inhibitory concentration of essential oils. Crit. Rev. Food Sci. Nutr. 2019, 59, 357–378. [Google Scholar] [CrossRef]
- Sergio, A.-O.; Fabiola, C.-V.; Guadalupe, N.-M.; Blanca, R.-C.; León, H.-O. Evaluation of antimycobacterium activity of the essential oils of cumin (Cuminum cyminum), clove (Eugenia caryophyllata), cinnamon (Cinnamomum verum), laurel (Laurus nobilis) and anis (Pimpinella anisum) against Mycobacterium tuberculosis. Adv. Biol. Chem. 2013, 3, 480–484. [Google Scholar] [CrossRef] [Green Version]
- Aldana, D.S.; Ochoa, S.A.; Aguilar, C.N.; Contreras-Esquivel, J.C.; Nevárez-Moorillón, G.V. Antibacterial activity of pectic-based edible films incorporated with Mexican lime essential oil. Food Control. 2015, 50, 907–912. [Google Scholar] [CrossRef]
- Rasooli, I.; Mirmostafa, S.A. Bacterial Susceptibility to and Chemical Composition of Essential Oils from Thymus kotschyanusand Thymus persicus. J. Agric. Food Chem. 2003, 51, 2200–2205. [Google Scholar] [CrossRef]
- Rasooli, I. Antimycotoxigenic characteristics of Rosmarinus officinalis and Trachyspermum copticum L. essential oils. Int. J. Food Microbiol. 2008, 122, 135–139. [Google Scholar] [CrossRef]
- Sykes, M.L.; Avery, V.M. Development of an Alamar Blue™ Viability Assay in 384-Well Format for High Throughput Whole Cell Screening of Trypanosoma brucei brucei Bloodstream Form Strain 427. Am. J. Trop. Med. Hyg. 2009, 81, 665–674. [Google Scholar] [CrossRef] [Green Version]
- Ordóñez-Gutiérrez, L.; Espada-Fernández, R.; Dea-Ayuela, M.A.; Torrado, J.J.; Bolás-Fernandez, F.; Alunda, J.M. In vitro effect of new formulations of amphotericin B on amastigote and promastigote forms of Leishmania infantum. Int. J. Antimicrob. Agents 2007, 30, 325–329. [Google Scholar] [CrossRef] [PubMed]
- Finney, J. Statistical logic in the monitoring of reactions to therapeutic drugs. Methods Inf. Med. 1971, 10, 237–245. [Google Scholar] [CrossRef] [PubMed]
- Tariq, S.; Wani, S.; Rasool, W.; Shafi, K.; Bhat, M.A.; Prabhakar, A.; Shalla, A.H.; Rather, M.A. A comprehensive review of the antibacterial, antifungal and antiviral potential of essential oils and their chemical constituents against drug-resistant microbial pathogens. Microb. Pathog. 2019, 134, 103580. [Google Scholar] [CrossRef]
- Guimarães, A.C.; Meireles, L.M.; Lemos, M.F.; Guimarães, M.C.C.; Endringer, D.C.; Fronza, M.; Scherer, R. Antibacterial Activity of Terpenes and Terpenoids Present in Essential Oils. Molecules 2019, 24, 2471. [Google Scholar] [CrossRef] [Green Version]
- Nazzaro, F.; Fratianni, F.; De Martino, L.; Coppola, R.; De Feo, V. Effect of Essential Oils on Pathogenic Bacteria. Pharmyst 2013, 6, 1451–1474. [Google Scholar] [CrossRef]
- Tang, C.; Chen, J.; Zhang, L.; Zhang, R.; Zhang, S.; Ye, S.; Zhao, Z.; Yang, D. Exploring the antibacterial mechanism of essential oils by membrane permeability, apoptosis and biofilm formation combination with proteomics analysis against methicillin-resistant staphylococcus aureus. Int. J. Med. Microbiol. 2020, 310, 151435. [Google Scholar] [CrossRef] [PubMed]
- Machado, M.; Dinis, A.; Salgueiro, L.; Custódio, J.B.; Cavaleiro, C.; Sousa, M. Anti-Giardia activity of Syzygium aromaticum essential oil and eugenol: Effects on growth, viability, adherence and ultrastructure. Exp. Parasitol. 2011, 127, 732–739. [Google Scholar] [CrossRef]
- Owen, L.; Laird, K.; Wilson, P.B. Structure-activity modelling of essential oils, their components, and key molecular parameters and descriptors. Mol. Cell. Probes 2018, 38, 25–30. [Google Scholar] [CrossRef] [Green Version]
- Afrin, F.; Chouhan, G.; Islamuddin, M.; Want, M.Y.; Ozbak, H.A.; Hemeg, H.A. Cinnamomum cassia exhibits antileishmanial activity against Leishmania donovani infection in vitro and in vivo. PLoS Negl. Trop. Dis. 2019, 13, e0007227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaroque, G.N.; Sartorelli, P.; Caseli, L. Interfacial vibrational spectroscopy and Brewster angle microscopy distinguishing the interaction of terpineol in cell membrane models at the air-water interface. Biophys. Chem. 2019, 246, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Gow, N.A.R.; Latge, J.-P.; Munro, C.A. The Fungal Cell Wall: Structure, Biosynthesis, and Function. Fungal King. 2017, 2017, 267–292. [Google Scholar] [CrossRef] [Green Version]
- Ochoa, S.A.; Nevárez-Moorillón, G.V.; Sánchez-Torres, L.E.; Villanueva-García, M.; Sánchez-Ramírez, B.E.; Rodríguez-Valdez, L.M.; Rivera-Chavira, B.E. Quantitative structure-activity relationship of molecules constituent of different essential oils with antimycobacterial activity against Mycobacterium tuberculosis and Mycobacterium bovis. BMC Complement. Altern. Med. 2015, 15, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Nazzaro, F.; Fratianni, F.; Coppola, R.; De Feo, V. Essential Oils and Antifungal Activity. Pharmers 2017, 10, 86. [Google Scholar] [CrossRef] [Green Version]
- Yutani, M.; Hashimoto, Y.; Ogita, A.; Kubo, I.; Tanaka, T.; Fujita, K.-I. Morphological Changes of the Filamentous Fungus Mucor mucedo and Inhibition of Chitin Synthase Activity Induced by Anethole. Phytotherapy Res. 2011, 25, 1707–1713. [Google Scholar] [CrossRef]
- Rajput, S.B.; Karuppayil, S.M. Small molecules inhibit growth, viability and ergosterol biosynthesis in Candida albicans. SpringerPlus 2013, 2, 1–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Da Silva, V.D.; Almeida-Souza, F.; Teles, A.M.; Neto, P.A.; Mondego-Oliveira, R.; Filho, N.E.M.; Taniwaki, N.N.; Abreu-Silva, A.L.; Calabrese, K.D.S.; Filho, V.E.M. Chemical composition of Ocimum canum Sims. essential oil and the antimicrobial, antiprotozoal and ultrastructural alterations it induces in Leishmania amazonensis promastigotes. Ind. Crop. Prod. 2018, 119, 201–208. [Google Scholar] [CrossRef]
- Yang, S.-K.; Yusoff, K.; Thomas, W.; Akseer, R.; Alhosani, M.S.; Abushelaibi, A.; Lim, E.; Lai, K.-S. Lavender essential oil induces oxidative stress which modifies the bacterial membrane permeability of carbapenemase producing Klebsiella pneumoniae. Sci. Rep. 2020, 10, 1–14. [Google Scholar] [CrossRef]
- Ogungbe, I.V.; Setzer, W.N. In-silico Leishmania Target Selectivity of Antiparasitic Terpenoids. Molecules 2013, 18, 7761–7847. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andrade-Ochoa, S.; Correa-Basurto, J.; Rodríguez-Valdez, L.M.; Sánchez-Torres, L.E.; Nogueda-Torres, B.; Nevárez-Moorillón, G.V. In vitro and in silico studies of terpenes, terpenoids and related compounds with larvicidal and pupaecidal activity against Culex quinquefasciatus Say (Diptera: Culicidae). Chem. Cent. J. 2018, 12, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Andrade-Ochoa, S.; Sánchez-Torres, L.E.; Rodríguez-Valdéz, L.M.; Nevárez-Moorillón, G.V.; Rivera-Chavira, B.E. Theoretical characterization of carvacrol, thymol and related compounds: Structural requirements for their antibacterial activity. In Advances in Medicine and Biology; Berhard, L.V., Ed.; Nova Science Publishers, Inc.: Hauppauge, NY, USA, 2017; Volume 199, pp. 179–200. ISBN 978-1-53611-105-7. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Content % | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| No. | RI | Compound | Anise | Cinnamon | Clove | Cumin | Laurel | Lime | Oregano |
| 1 | 908 | α-Thujene | - | - | - | 0.2 | - | - | - |
| 2 | 934 | α-Pinene | - | 0.4 | - | 0.5 | 6.1 | 1.2 | - |
| 3 | 950 | Camphene | - | 0.5 | - | - | - | - | - |
| 4 | 964 | Sabinene | - | - | - | - | 8.1 | - | - |
| 5 | 979 | β-Pinene | - | 0.3 | - | 7.8 | - | 1.3 | - |
| 6 | 988 | β-Myrcene | 2.2 | - | - | 0.6 | 0.1 | 0.5 | 1.5 |
| 7 | 1009 | 3-Carene | - | 0.2 | - | 0.1 | - | - | - |
| 8 | 1007 | α-Phellandrene | - | 1.1 | - | 0.1 | - | - | - |
| 9 | 1018 | α-Terpinene | - | 0.8 | - | 0.1 | - | - | 2.1 |
| 10 | 1031 | Limonene | - | 0.6 | - | 0.4 | - | 92.3 | - |
| 11 | 1032 | β-Phellandrene | - | 1.3 | - | - | - | 0.1 | - |
| 12 | 1027 | p-Cymene | - | 0.6 | - | 9.5 | - | - | 24.6 |
| 13 | 1087 | α-Terpinolene | - | 0.2 | - | - | 2.3 | - | - |
| 14 | 964 | Benzaldehyde | - | 1.2 | - | - | - | - | - |
| 15 | 1019 | Eucalyptol | - | - | 0.1 | 0.3 | 70.7 | - | 0.9 |
| 16 | 1055 | γ-Terpinene | - | - | - | 11.1 | 1.4 | 0.1 | - |
| 17 | 1088 | α-Terpinolene | - | - | - | 0.1 | 1.2 | - | - |
| 18 | 1102 | Linalool | - | 1.1 | 0.1 | 0.2 | 0.7 | - | - |
| 19 | 1130 | Pulegone | - | - | - | 0.1 | - | - | - |
| 20 | 1143 | Terpineol-4 | - | - | - | 0.6 | 3.3 | - | 2.4 |
| 21 | 1160 | Isopulegone | - | - | - | 0.4 | - | - | - |
| 22 | 1176 | Myrtenol | - | - | - | 0.1 | - | - | - |
| 23 | 1196 | Estragole | 1.4 | - | - | - | - | - | - |
| 24 | 1200 | p-Anisaldehyde | 1.9 | - | - | - | - | - | - |
| 25 | 1228 | Cuminaldehyde | - | - | - | 41.3 | - | - | - |
| 26 | 1236 | Chavicol | 2.8 | - | - | - | - | - | - |
| 27 | 1255 | Phellandral | - | - | - | 0.2 | - | - | - |
| 28 | 1270 | Safranal | - | - | - | 1.9 | - | - | - |
| 29 | 1273 | Thymol | - | - | - | - | - | - | 58.3 |
| 30 | 1277 | Carvacrol | - | - | - | - | - | - | 3.4 |
| 31 | 1283 | Cuminic alcohol | - | - | - | 16.9 | - | - | - |
| 32 | 1295 | trans-Anethole | 88.9 | - | - | - | - | - | - |
| 33 | 1315 | p-Mentha-1,4-dien-7-ol | - | - | - | 0.3 | - | - | - |
| 34 | 1425 | β-Caryophyllene | - | 3.2 | 24.9 | 0.1 | 0.3 | - | 3.4 |
| 35 | 1463 | Humulene | - | 0.6 | 2.8 | - | - | - | 0.8 |
| 36 | 1200 | α-Terpineol | - | 0.5 | - | - | - | 1.1 | - |
| 37 | 1167 | Hydrocinnamic aldehyde | - | 0.2 | - | - | - | - | - |
| 38 | 1375 | Hydrocinnamyl acetate | - | 0.2 | - | - | - | - | - |
| 39 | 1591 | Caryophyllene oxide | - | 0.5 | - | - | 0.1 | - | - |
| 40 | 1279 | Cinnamaldehyde | - | 73.3 | - | - | - | - | - |
| 41 | 1453 | Cinnamyl acetate | - | 5.1 | - | - | - | - | - |
| 42 | 1357 | Eugenol | - | 2 | 61.3 | - | 0.3 | - | - |
| 43 | 1368 | Methyleugenol | - | - | - | - | 0.7 | - | - |
| 44 | 1313 | Cinnamyl alcohol | - | 0.3 | - | - | - | - | - |
| 45 | 1450 | Cinnamic acid | - | 0.1 | - | - | - | - | - |
| 46 | 1482 | Eugenyl acetate | - | - | 4.2 | - | - | - | - |
| 47 | 1491 | β-Cadinene | - | - | 3.6 | - | - | - | - |
| 48 | 1508 | Epizonarene | - | - | 0.1 | - | - | - | - |
| 49 | 1777 | Benzyl benzoate | - | 0.5 | - | - | - | - | - |
| Total | 97.2 | 94.8 | 97.1 | 92.7 | 95.3 | 96.6 | 97.4 | ||
| Identified components | 5 | 24 | 8 | 24 | 13 | 7 | 9 | ||
| Assays | MIC (µg/mL) | |||||
|---|---|---|---|---|---|---|
| Gram Positive | Gram Negative | |||||
| Essential Oils | B. cereus | S. aureus | L. monocytogenes | E. coli | S. typhimurium | S. sonnei |
| Anise | 383 ± 157.74 a | 316 ± 57.74 ab | 416 ± 57.74 b | 483 ± 28.87 ab | 483 ± 28.87 ab | 466 ± 28.87 ab |
| Cinnamon | 350 ± 0.00 ab | 283 ± 57.74 ab | 416 ± 57.74 b | 416 ± 57.74 bc | 416 ± 57.74 bc | 383 ± 57.74 bc |
| Clove | 283 ± 57.74 abc | 416 ± 157.74 a | 483 ± 28.87 b | 383 ± 57.74 bc | 383 ± 57.74 bc | 550 ± 86.60 ab |
| Cumin | 150 ± 86.60 cd | 200 ± 86.60 bc | 650 ± 0.00 a | 600 ± 86.60 a | 600 ± 86.60 a | 600 ± 86.60 a |
| Laurel | 216 ± 57.74 bcd | 216 ± 57.74 bc | 283 ± 57.74 c | 283 ± 57.74 c | 283 ± 57.74 c | 283 ± 57.74 c |
| Lime | 283 ± 57.74 ab | 216 ± 57.74 bc | 283 ± 57.74 c | 283 ± 57.74 c | 283 ± 57.74 c | 283 ± 57.74 c |
| Oregano | 83 ± 28.87 d | 83 ± 14.43 c | 100 ± 0.00 d | 66 ± 28.87 d | 66 ± 28.87 d | 100 ± 0.00 d |
| Major Compounds | B. cereus | S. aureus | L. monocytogenes | E. coli | S. typhimurium | S. sonnei |
| Anethole | 783± 57.24 a | 150 ± 86.60 a | 133 ± 28.87 d | 216 ± 57.74 b | 716 ± 57.74 a | 483 ± 28.87 a |
| Cinamaldehyde | 91 ± 14.43 c | 58 ± 14.43b | 100 ± 0.00 d | 100 ± 0.00 c | 600 ± 86.60 ab | 283 ± 57.74 b |
| Eugenol | 250 ± 0.00 b | 50 ± 0.00 b | 216 ± 57.74 c | 133 ± 28.87 c | 416 ± 144.34 b | 316 ± 57.74 b |
| Cuminaldehyde | 116 ± 28.87 c | 41 ± 14.43 b | 91 ± 14.43 e | 50 ± 0.00 d | 316 ± 57.74 bc | 383 ± 57.74 ab |
| Eucalyptol | 83 ± 28.87 c | 141 ± 14.44 a | 416 ± 144.34 b | 600 ± 86.60 a | 666 ± 144.34 ab | 466 ± 28.87 a |
| Limonene | 750 ± 0.00 ab | 66 ± 14.43 b | 833 ± 144.34 a | 583 ± 144.34 a | 250 ± 0.00 c | 416 ± 57.74 a |
| Carvacrol | 91 ± 14.43 c | 23 ± 2.89 c | 200 ± 86.60 c | 83 ± 14.43 c | 41 ± 14.43 d | 83 ± 14.43 c |
| Thymol | 33 ± 14.43 d | 23 ± 2.89 c | 91 ± 14.43 e | 41 ± 14.43 d | 33 ± 14.43 d | 25 ± 0.00 d |
| Assays | MIC (µg/mL) | |||
|---|---|---|---|---|
| Essential Oils | A. niger | A. ochraceus | A. alternata | F. oxysporum |
| Anise | 150 ± 86.60 bc | 150 ± 8.60 bc | 200 ± 86.60 bc | 150 ± 86.60 bc |
| Cinnamon | 66 ± 14.43 c | 83 ± 14.43 c | 66 ± 14.43 d | 116 ± 28.87 c |
| Clove | 150 ± 86.60 bc | 100 ± 0.00 c | 100 ± 0.00 cd | 116 ± 28.87 c |
| Cumin | 283 ± 57.74 b | 283 ± 57.74 b | 250 ± 57.74 b | 316 ± 57.74 b |
| Laurel | 483 ± 28.87 a | 500 ± 0.00 a | 450 ± 0.00 a | 450 ± 28.87 a |
| Lime | 316 ± 57.74 ab | 283 ± 57.74 b | 483 ± 28.87 a | 450 ± 28.87 a |
| Oregano | 250 ± 0.00 b | 283 ± 57.74 b | 200 ± 86.60 bc | 316 ± 57.74 b |
| Major Compounds | A. niger | A. ochraceus | A. alternata | F. oxysporum |
| Anethole | 750 ± 0.00 b | 483 ± 28.42 c | 500 ± 0.00 c | 650 ± 0.00 c |
| Cinamaldehyde | 75 ± 0.00 e | 58.3 ± 14.06 e | 41± 14.06 e | 58 ± 14.06 f |
| Eugenol | 100 ± 0.00 d | 66 ± 14.06 e | 41 ± 14.06 e | 66 ± 14.06 f |
| Cuminaldehyde | 283 ± 57.17 cd | 316 ± 57.17 cd | 66 ± 14 e | 91 ± 14.06 e |
| Eucalyptol | 716 ± 57.17 b | 683.3 ± 57.17 b | 716.6 ± 57.17 b | 716 ± 57.17 b |
| Limonene | 950 ± 0.00 a | 1000 ± 0.00 a | 750 ± 0.00 ab | 1000 ± 0.00 a |
| Carvacrol | 466 ± 28.42 bc | 466 ± 28.42 c | 783 ± 57.44 a | 616 ± 202.57 c |
| Thymol | 350 ± 0.00 c | 250 ± 0.00 d | 200 ± 86.23 d | 283 ± 57.44 d |
| Assays | IC50 (µg/mL) | LD50 (µg/mL) | |
|---|---|---|---|
| Essential Oils | T. cruzi | L. mexicana | G. lamblia |
| Anise | 52 ± 36.62 c | 63 ± 0.91 c | 136 ± 32.8 b |
| Cinnamon | 22 ± 0.22 f | 21 ± 25.13 e | 108 ± 22.53 e |
| Clove | 56 ± 7.88 d | 56 ± 8.35 c | 139 ± 11.35 bc |
| Cumin | 131 ± 115.53 e | 130 ± 19.02 d | 175 ± 43.62 d |
| Laurel | 223 ± 24.7 a | 209 ± 27.62 a | 193 ± 54.44 a |
| Lime | 98 ± 36.02 b | 88 ± 26.02 b | 112 ± 54.88 cd |
| Oregano | 23 ± 24.62 g | 59 ± 28.13 e | 60 ± 12.13 f |
| Major Compounds | T. cruzi | L. mexicana | G. lamblia |
| Anethole | 47.17 ± 5.90 d | 47.30 ± 2.32 d | 134.99 ± 0.65 b |
| Cinamaldehyde | 10.45 ± 0.19 g | 8.66 ± 2.18 f | 76.42 ± 2.79 e |
| Eugenol | 14.95 ± 1.11 f | 12.91 ± 12.29 ef | 104.04 ± 3.26 d |
| Cuminaldehyde | 65.92 ± 2.38 c | 55.96 ± 2.40 d | 141.16 ± 1.02 b |
| Eucalyptol | 275.43 ± 3.03 a | 262.09 ± 1.10 a | 265.43 ± 3.02 a |
| Limonene | 211.61± 1.63 b | 208.91 ± 3.13 b | 127.59 ± 316 c |
| Carvacrol | 36.43 ± 4.75 e | 79.77 ± 1.28 c | 31.92 ± 0.56 f |
| Thymol | 16.62 ± 1.71 f | 21.22 ± 1.30 e | 21.44 ± 52.07 g |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Andrade-Ochoa, S.; Chacón-Vargas, K.F.; Sánchez-Torres, L.E.; Rivera-Chavira, B.E.; Nogueda-Torres, B.; Nevárez-Moorillón, G.V. Differential Antimicrobial Effect of Essential Oils and Their Main Components: Insights Based on the Cell Membrane and External Structure. Membranes 2021, 11, 405. https://doi.org/10.3390/membranes11060405
Andrade-Ochoa S, Chacón-Vargas KF, Sánchez-Torres LE, Rivera-Chavira BE, Nogueda-Torres B, Nevárez-Moorillón GV. Differential Antimicrobial Effect of Essential Oils and Their Main Components: Insights Based on the Cell Membrane and External Structure. Membranes. 2021; 11(6):405. https://doi.org/10.3390/membranes11060405
Chicago/Turabian StyleAndrade-Ochoa, Sergio, Karla Fabiola Chacón-Vargas, Luvia Enid Sánchez-Torres, Blanca Estela Rivera-Chavira, Benjamín Nogueda-Torres, and Guadalupe Virginia Nevárez-Moorillón. 2021. "Differential Antimicrobial Effect of Essential Oils and Their Main Components: Insights Based on the Cell Membrane and External Structure" Membranes 11, no. 6: 405. https://doi.org/10.3390/membranes11060405
APA StyleAndrade-Ochoa, S., Chacón-Vargas, K. F., Sánchez-Torres, L. E., Rivera-Chavira, B. E., Nogueda-Torres, B., & Nevárez-Moorillón, G. V. (2021). Differential Antimicrobial Effect of Essential Oils and Their Main Components: Insights Based on the Cell Membrane and External Structure. Membranes, 11(6), 405. https://doi.org/10.3390/membranes11060405