
Modeling and Optimization of Gas Sparging-Assisted Bacterial Cultivation Broth Microfiltration by Response Surface Methodology and Genetic Algorithm


 ,
,  , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Production of Bacillus velezensis Cultivation Broth
2.2. Microfiltration Experimental Setup
2.3. Experimental Data Analysis—Modeling and Optimization
3. Results
3.1. Modeling of Gas Sparging-Assisted Microfiltration of Bacillus velezensis IP22 Cultivation Broth
3.2. Optimization of Gas Sparging-Assisted Microfiltration of Bacillus velezensis IP22 Cultivation Broth
4. Discussion
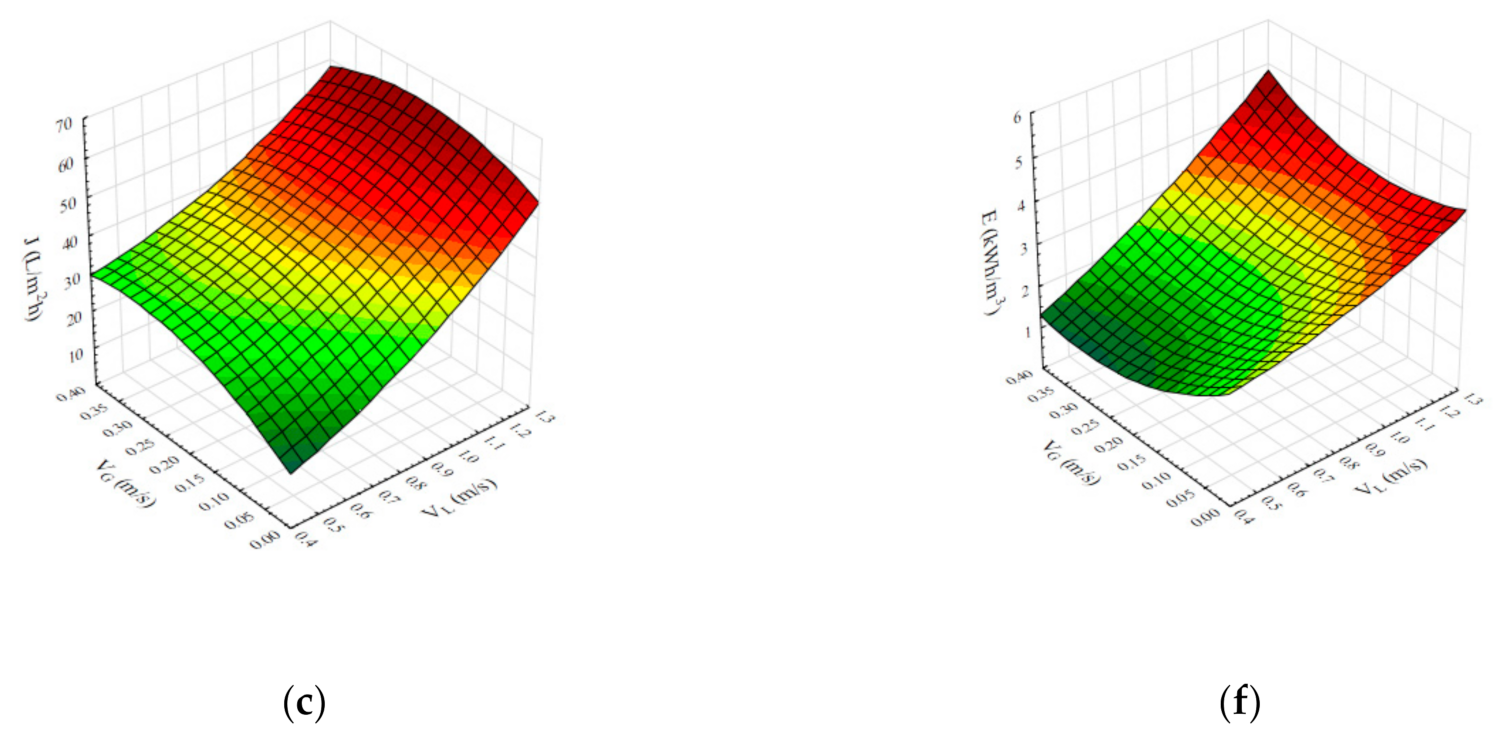
4.1. The Effects of Operational Conditions on Steady State Permeate Flux during Air Sparging-Assisted Microfiltration of Bacillus velezensis IP22 Cultivation Broth
4.2. The Effects of Operational Conditions on Specific Energy Consumption during Air Sparging-Assisted Microfiltration of Bacillus velezensis Cultivation Broth
4.3. Optimization of Operational Conditions for Air Sparging-Assisted Microfiltration of Bacillus velezensis Cultivation Broth
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Cidón, C.F.; Figueiró, P.S.; Schreiber, D. Benefits of organic agriculture under the perspective of the bioeconomy: A systematic review. Sustainability 2021, 13, 6852. [Google Scholar] [CrossRef]
- Seiber, J.N.; Coats, J.; Duke, S.O.; Gross, A.D. Biopesticides: State of the art and future opportunities. J. Agric. Food Chem. 2014, 62, 11613–11619. [Google Scholar] [CrossRef] [Green Version]
- Ruiu, L. Microbial biopesticides in agroecosystems. Agronomy 2018, 8, 235. [Google Scholar] [CrossRef] [Green Version]
- Reganold, J.; Wachter, J. Organic agriculture in the twenty-first century. Nat. Plants 2016, 2, 15221. [Google Scholar] [CrossRef]
- Muller, A.; Schader, C.; El-Hage Scialabba, N.; Brüggemann, J.; Isensee, A.; Erb, K.; Smith, P.; Klocke, P.; Leiber, F.; Stolze, M.; et al. Strategies for feeding the world more sustainably with organic agriculture. Nat. Commun. 2017, 8, 1290. [Google Scholar] [CrossRef] [Green Version]
- Shafi, J.; Tian, H.; Ji, M. Bacillus species as versatile weapons for plant pathogens: A review. Biotechnol. Biotechnol. Equip. 2017, 31, 446–459. [Google Scholar] [CrossRef] [Green Version]
- Fira, Đ.; Dimkić, I.; Berić, T.; Lozo, J.; Stanković, S. Biological control of plant pathogens by Bacillus species. J. Bitechnol. 2018, 285, 44–55. [Google Scholar] [CrossRef]
- Villarreal-Delgado, M.F.; Villa-Rodríguez, E.D.; Cira-Chávez, L.A.; Estrada-Alvarado, M.I.; Parra-Cota, F.I.; de los Santos-Villalobos, S. The genus Bacillus as a biological control agent and its implications in the agricultural biosecurity. Rev. Mex. Fitopatol. 2018, 36, 95–130. [Google Scholar] [CrossRef]
- Rabbee, M.F.; Ali, M.S.; Choi, J.; Hwang, B.S.; Jeong, S.C.; Baek, K.-h. Bacillus velezensis: A valuable member of bioactive molecules within plant microbiomes. Molecules 2019, 24, 1046. [Google Scholar] [CrossRef] [Green Version]
- Ngalimat, M.S.; Yahaya, R.S.R.; Baharudin, M.M.A.-a.; Yaminudin, S.M.; Karim, M.; Ahmad, S.A.; Sabri, S. A review on the biotechnological applications of the operational group Bacillus amyloliquefaciens. Microorganisms 2021, 9, 614. [Google Scholar] [CrossRef] [PubMed]
- Pajčin, I.; Vlajkov, V.; Frohme, M.; Grebinyk, S.; Grahovac, M.; Mojićević, M.; Grahovac, J. Pepper bacterial spot control by Bacillus velezensis: Bioprocess solution. Microorganisms 2020, 8, 1463. [Google Scholar] [CrossRef]
- Grahovac, J.; Pajčin, I.; Vlajkov, V.; Rončević, Z.; Dodić, J.; Cvetković, D.; Jokić, A. Xanthomonas campestris biocontrol agent: Selection, medium formulation and bioprocess kinetic analysis. Chem. Ind. Chem. Eng. Q. 2021, in press. [Google Scholar] [CrossRef]
- Prabakaran, G.; Hoti, S.L. Application of different downstream processing methods and their comparison for the large-scale preparation of Bacillus thuringiensis var. israelensis after fermentation for mosquito control. Biologicals 2008, 36, 412–415. [Google Scholar] [CrossRef]
- Brar, S.K.; Verma, M.; Tyagi, R.D.; Valéro, J.R. Recent advances in downstream processing and formulations of Bacillus thuringiensis based biopesticides. Process Biochem. 2006, 41, 323–342. [Google Scholar] [CrossRef]
- Qi, L.; Hu, Y.; Chai, Q.; Wang, Q. Enhanced filtration performance and anti-biofouling properties of antibacterial polyethersulfone membrane for fermentation broth concentration. J. Ind. Eng. Chem. 2019, 72, 346–353. [Google Scholar] [CrossRef]
- Shimizu, Y.; Matsushita, K.; Watanabe, A. Influence of shear breakage of microbial cells on cross-flow microfiltration flux. J. Ferment. Bioeng. 1994, 78, 170–174. [Google Scholar] [CrossRef]
- Jana, A.; Ghosh, S.; Majumdar, S. Energy efficient harvesting of Arthrospira sp. using ceramic membranes: Analyzing the effect of membrane pore size and incorporation of flocculant as fouling control strategy. J. Chem. Technol. Biotechnol. 2018, 93, 1085–1096. [Google Scholar] [CrossRef]
- Aspelund, M.T.; Rozeboom, G.; Heng, M.; Glatz, C.E. Improving permeate flux and product transmission in the microfiltration of a bacterial cell suspension by flocculation with cationic polyelectrolytes. J. Membr. Sci. 2008, 324, 198–208. [Google Scholar] [CrossRef]
- Salama, A. Modeling of flux decline behavior during the filtration of oily-water systems using porous membranes: Effect of pinning of nonpermeating oil droplets. Sep. Purif. Technol. 2018, 207, 240–254. [Google Scholar] [CrossRef]
- Fan, R.; Ebrahimi, M.; Quitmann, H.; Czermak, P. Lactic acid production in a membrane bioreactor system with thermophilic Bacillus coagulans: Fouling analysis of the used ceramic membranes. Sep. Sci. Technol. 2015, 50, 2177–2189. [Google Scholar] [CrossRef]
- Zhang, Y.; Fu, Q. Algal fouling of microfiltration and ultrafiltration membranes and control strategies: A review. Sep. Purif. Technol. 2018, 203, 193–208. [Google Scholar] [CrossRef]
- Chang, Y.-R.; Lee, D.J. Coagulation–membrane filtration of Chlorella vulgaris at different growth phases. Dry. Technol. 2012, 30, 1317–1322. [Google Scholar] [CrossRef]
- Marzban, R.; Saberi, F.; Shirazi, M.M.A. Microfiltration and ultrafiltration of Bacillus thuringiensis fermentation broth: Membrane performance and spore crystal recovery approaches. Braz. J. Chem. Eng. 2016, 33, 783–791. [Google Scholar] [CrossRef] [Green Version]
- Jokić, A.; Zavargo, Z.; Šereš, Z.; Tekić, M. The effect of turbulence promoter on cross-flow microfiltration of yeast suspensions: A response surface methodology approach. J. Membr. Sci. 2010, 350, 269–278. [Google Scholar] [CrossRef]
- Jokić, A.; Pajčin, I.; Grahovac, J.; Lukić, N.; Dodić, J.; Rončević, Z.; Šereš, Z. Improving energy efficiency of Bacillus velezensis broth microfiltration in tubular ceramic membrane by air sparging and turbulence promoter. J. Chem. Technol. Biotechnol. 2020, 95, 1110–1115. [Google Scholar] [CrossRef]
- Jokić, A.; Pajčin, I.; Grahovac, J.; Lukić, N.; Ikonić, B.; Nikolić, N.; Vlajkov, V. Dynamic modeling using artificial neural network of Bacillus Velezensis broth cross-flow microfiltration enhanced by air-sparging and turbulence promoter. Membranes 2020, 10, 372. [Google Scholar] [CrossRef]
- Tanaka, T.; Usui, K.; Kouda, K.; Nakanishi, K. Filtration behaviors of rod-shaped bacterial broths in unsteady-state phase of cross-flow filtration. J. Chem. Eng. Jpn. 1996, 29, 973–981. [Google Scholar] [CrossRef] [Green Version]
- Guerra, A.; Jonsson, G.; Rasmussen, A.; Waagner Nielsen, E.; Edelsten, D. Low cross-flow velocity microfiltration of skim milk for removal of bacterial spores. Int. Dairy J. 1997, 7, 849–861. [Google Scholar] [CrossRef]
- Tomasula, P.M.; Mukhopadhyay, S.; Datta, N.; Porto-Fett, A.; Call, J.E.; Luchansky, J.B.; Renye, J.; Tunick, M. Pilot-scale crossflow-microfiltration and pasteurization to remove spores of Bacillus anthracis (Sterne) from milk. J. Dairy Sci. 2011, 94, 4277–4291. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.-H.; Min, C.-S. Fouling reduction using the resonance vibration in membrane separation of whole milk. J. Ind. Eng. Chem. 2019, 75, 123–129. [Google Scholar] [CrossRef]
- Jiang, B.; Hu, B.; Yang, N.; Zhang, L.; Sun, Y.; Xiao, X. Study of turbulence promoters in prolonging membrane life. Membranes 2021, 11, 268. [Google Scholar] [CrossRef]
- Cabassud, C.; Laborie, S.; Durand-Bourlier, L.; Lainé, J.M. Air sparging in ultrafiltration hollow fibers: Relationship between flux enhancement, cake characteristics and hydrodynamic parameters. J. Membr. Sci. 2001, 181, 57–69. [Google Scholar] [CrossRef]
- Hwang, K.-J.; Wu, Y.J. Flux enhancement and cake formation in air-sparged cross-flow microfiltration. Chem. Eng. J. 2008, 139, 296–303. [Google Scholar] [CrossRef]
- Hwang, K.-J.; Hsu, C.-E. Effect of gas–liquid flow pattern on air-sparged cross-flow microfiltration of yeast suspension. Chem. Eng. J. 2009, 151, 160–167. [Google Scholar] [CrossRef]
- Cui, Z.F.; Wright, K.I.T. Flux enhancements with gas sparging in downwards crossflow ultrafiltration: Performances and mechanisms. J. Membr. Sci. 1996, 117, 109–116. [Google Scholar] [CrossRef]
- Mercier, M.; Fonade, C.; Lafforque-Delorme, C. How slug flow can enhance the ultrafiltration flux in mineral tubular membranes. J. Membr. Sci. 1997, 128, 103–113. [Google Scholar] [CrossRef]
- Mercier-Bonin, M.; Lagane, C.; Fonade, C. Influence of a gas/liquid two-phase flow on the ultrafiltration and microfiltration performances: Case of a ceramic flat sheet membrane. J. Membr. Sci. 2000, 180, 93–102. [Google Scholar] [CrossRef]
- Gupta, B.S.; Hasan, S.; Hashim, M.A.; Cui, Z.F. Effects of colloidal fouling and gas sparging on microfiltration of yeast suspension. Bioprocess. Biosyst. Eng. 2005, 27, 407–413. [Google Scholar] [CrossRef]
- Sur, H.W.; Cui, Z.F. Enhancement of microfiltration of yeast suspensions using gas sparging—Effect of feed conditions. Sep. Purif. Technol. 2005, 41, 313–319. [Google Scholar] [CrossRef]
- Hwang, K.-J.; Chen, L. Effect of air-sparging on the cross-flow microfiltration of microbe/protein bio-suspension. J. Taiwan Inst. Chem. Eng. 2010, 41, 564–569. [Google Scholar] [CrossRef]
- Mikulášek, P.; Pospišil, P.; Dolecek, P.; Cakl, J. Gas–liquid two-phase flow in microfiltration mineral tubular membranes: Relationship between flux enhancement and hydrodynamic parameters. Desalination 2002, 146, 103–109. [Google Scholar] [CrossRef]
- Mercier-Bonin, M.; Gésan-Guiziou, G.; Fonade, C. Application of gas/liquid two-phase flows during cross flow microfiltration of skimmed milk under constant transmembrane pressure conditions. J. Membr. Sci. 2003, 218, 93–105. [Google Scholar] [CrossRef]
- Fouladitajar, A.; Ashtiani, F.Z.; Rezaei, H.; Haghmoradi, A.; Kargari, A. Gas sparging to enhance permeate flux and reduce fouling resistances in cross flow microfiltration. J. Ind. Eng. Chem. 2014, 20, 624–632. [Google Scholar] [CrossRef]
- Javadi, N.; Ashtiani, F.Z.; Fouladitajar, A.; Zenooz, A.M. Experimental studies and statistical analysis of membrane fouling behavior and performance in microfiltration of microalgae by a gas sparging assisted process. Bioresour. Technol. 2014, 162, 350–357. [Google Scholar] [CrossRef] [PubMed]
- Hyder, M.N.; Huang, R.Y.M.; Chen, P. Pervaporation dehydration of alcohol–water mixtures: Optimization for permeate flux and selectivity by central composite rotatable design. J. Membr. Sci. 2009, 326, 343–353. [Google Scholar] [CrossRef]
- Han, H.; Yu, R.; Li, B.; Zhang, Y.; Wang, W.; Chen, X. Multi-objective optimization of corrugated tube with loose-fit twisted tape using RSM and NSGA-II. Int. J. Heat Mass Transf. 2019, 131, 781–794. [Google Scholar] [CrossRef]
- Mercier, M.; Fonade, C.; Lafforgue-Delorme, C. Influence of the flow regime on the efficiency of a gas-liquid two-phase medium filtration. Biotechnol. Tech. 1995, 9, 853–858. [Google Scholar] [CrossRef]
- Tanaka, T.; Abe, K.-I.; Asakawa, H.; Yoshida, H.; Nakanishi, K. Filtration characteristics and structure of cake in crossflow filtration of bacterial suspension. J. Ferment. Bioeng. 1994, 78, 455–461. [Google Scholar] [CrossRef]
- Huisman, I.H.; Trägårdh, C. Particle transport in crossflow microfiltration—I. Effects of hydrodynamics and diffusion. Chem. Eng. Sci. 1999, 54, 271–280. [Google Scholar] [CrossRef]
- Mota, M.; Teixeira, J.A.; Yelshin, A. Influence of cell-shape on the cake resistance in dead-end and cross-flow filtrations. Sep. Purif. Technol. 2002, 27, 137–144. [Google Scholar] [CrossRef] [Green Version]
- Hwang, K.-J.; Yu, Y.-H.; Lu, W.-M. Cross-flow microfiltration of submicron microbial suspension. J. Membr. Sci. 2001, 194, 229–243. [Google Scholar] [CrossRef]
- Tanaka, T.; Abe, K.-I.; Nakanishi, K. Shear-induced arrangement of cells in cake during crossflow filtration of Escherichia coli cells. Biotechnol. Tech. 1994, 8, 57–60. [Google Scholar] [CrossRef]
- Goel, T.; Vaidyanathan, R.; Haftka, R.T.; Shyy, W.; Queipo, N.V.; Tucker, K. Response surface approximation of Pareto optimal front in multi-objective optimization. Comput. Methods Appl. Mech. Eng. 2007, 196, 879–893. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Experiment | Factors—Independent Variables | Responses—Dependent Variables | |||
|---|---|---|---|---|---|
| TMP (bar) | VL (m∙s−1) | VG (m∙s−1) | J (L∙m−2·h−1) | E (kW·h·m−3) | |
| 1 | 0.2 (−1) | 0.43 (−1) | 0.2 (0) | 31.06 | 1.1 |
| 2 | 1.0 (1) | 0.43 (−1) | 0.2 (0) | 22.95 | 2.3 |
| 3 | 0.2 (−1) | 1.30 (1) | 0.2 (0) | 55.89 | 4.4 |
| 4 | 1.0 (1) | 1.30 (1) | 0.2 (0) | 70.00 | 3.9 |
| 5 | 0.2 (−1) | 0.87 (0) | 0.0 (−1) | 30.57 | 2.4 |
| 6 | 1.0 (1) | 0.87 (0) | 0.0 (−1) | 29.00 | 4.7 |
| 7 | 0.2 (−1) | 0.87 (0) | 0.4 (1) | 36.67 | 3.8 |
| 8 | 1.0 (1) | 0.87 (0) | 0.4 (1) | 41.47 | 2.5 |
| 9 | 0.6 (0) | 0.43 (−1) | 0.0 (−1) | 17.50 | 2.3 |
| 10 | 0.6 (0) | 1.30 (1) | 0.0 (−1) | 53.87 | 4.0 |
| 11 | 0.6 (0) | 0.43 (−1) | 0.4 (1) | 32.64 | 1.1 |
| 12 | 0.6 (0) | 1.30 (1) | 0.4 (1) | 58.05 | 4.6 |
| 13 | 0.6 (0) | 0.87 (0) | 0.2 (0) | 43.45 | 2.1 |
| 14 | 0.6 (0) | 0.87 (0) | 0.2 (0) | 42.80 | 2.1 |
| 15 | 0.6 (0) | 0.87 (0) | 0.2 (0) | 45.00 | 2.0 |
| Effects | Steady State Permeate Flux (L∙m−2·h−1) | Specific Energy Consumption (kW·h·m−3) | ||||
|---|---|---|---|---|---|---|
| Coefficient | p-Value | Coefficient | p-Value | |||
| Actual | Coded | Actual | Coded | |||
| Intercept | ||||||
| b0 | 20.25 | 43.56 | 0.0141 | 0.78 | 2.05 | 0.0302 |
| Linear | ||||||
| b1 | −10.54 | 1.12 | 0.1410 | 0.37 | 0.21 | 0.0009 |
| b2 | −9.62 | 16.71 | <0.0001 | 0.88 | 1.26 | <0.0001 |
| b3 | 107.98 | 4.75 | 0.0007 | −5.40 | −0.18 | 0.0021 |
| Quadratic | ||||||
| b11 | −15.20 | −2.43 | 0.0017 | 3.78 | 0.60 | 0.0002 |
| b22 | 20.34 | 3.85 | 0.0096 | 1.42 | 0.27 | <0.0001 |
| b33 | −172.28 | −6.89 | 0.0295 | 16.98 | 0.68 | 0.0001 |
| Interaction | ||||||
| b12 | 31.90 | 5.55 | 0.0499 | −2.45 | −0.42 | <0.0001 |
| b13 | 19.91 | 1.59 | 0.1400 | −11.25 | −0.90 | 0.0018 |
| b23 | −31.51 | −2.74 | 0.0008 | 5.17 | 0.45 | <0.0001 |
| Source | Response | DF | SS | MS | F-Value | p-Value | R2 |
|---|---|---|---|---|---|---|---|
| Model | J (L∙m−2·h−1) | 9 | 2845.19 | 316.13 | 95.80 | 0.000046 | 0.984 |
| E (kW·h∙m−3) | 9 | 21.06 | 2.34 | 318.23 | 0.000002 | 0.995 | |
| Residual | J (L∙m−2·h−1) | 5 | 16.50 | 3.30 | |||
| E (kW·h∙m−3) | 5 | 0.04 | 0.01 | ||||
| Lack-of-fit | J (L∙m−2·h−1) | 3 | 13.94 | 4.65 | 3.64 | 0.22 | |
| E (kW·h∙m−3) | 3 | 0.03 | 0.01 | 3.01 | 0.26 | ||
| Pure error | J (L∙m−2·h−1) | 2 | 2.56 | 1.28 | |||
| E (kW·h∙m−3) | 2 | 0.01 | 0.00 | ||||
| Total | J (L∙m−2·h−1) | 14 | 2861.68 | ||||
| E (kW·h∙m−3) | 14 | 21.10 |
| Factors—independent variables | Goal | Optimized value |
| Transmembrane pressure, TMP (bar) | in range | 0.68 |
| Superficial feed velocity, VL (m∙s−1) | in range | 0.96 |
| Superficial air velocity, VG (m∙s−1) | in range | 0.25 |
| Responses—dependent variables | Goal | Predicted value |
| Steady state permeate flux, J (L∙m−2·h−1) | maximize | 48.57 |
| Specific energy consumption, E (kW·h∙m−3) | minimize | 2.37 |
| Desirability function | 0.62 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jokić, A.; Pajčin, I.; Lukić, N.; Vlajkov, V.; Kiralj, A.; Dmitrović, S.; Grahovac, J. Modeling and Optimization of Gas Sparging-Assisted Bacterial Cultivation Broth Microfiltration by Response Surface Methodology and Genetic Algorithm. Membranes 2021, 11, 681. https://doi.org/10.3390/membranes11090681
Jokić A, Pajčin I, Lukić N, Vlajkov V, Kiralj A, Dmitrović S, Grahovac J. Modeling and Optimization of Gas Sparging-Assisted Bacterial Cultivation Broth Microfiltration by Response Surface Methodology and Genetic Algorithm. Membranes. 2021; 11(9):681. https://doi.org/10.3390/membranes11090681
Chicago/Turabian StyleJokić, Aleksandar, Ivana Pajčin, Nataša Lukić, Vanja Vlajkov, Arpad Kiralj, Selena Dmitrović, and Jovana Grahovac. 2021. "Modeling and Optimization of Gas Sparging-Assisted Bacterial Cultivation Broth Microfiltration by Response Surface Methodology and Genetic Algorithm" Membranes 11, no. 9: 681. https://doi.org/10.3390/membranes11090681
APA StyleJokić, A., Pajčin, I., Lukić, N., Vlajkov, V., Kiralj, A., Dmitrović, S., & Grahovac, J. (2021). Modeling and Optimization of Gas Sparging-Assisted Bacterial Cultivation Broth Microfiltration by Response Surface Methodology and Genetic Algorithm. Membranes, 11(9), 681. https://doi.org/10.3390/membranes11090681