

The “www” of Xenopus laevis Oocytes: The Why, When, What of Xenopus laevis Oocytes in Membrane Transporters Research

 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
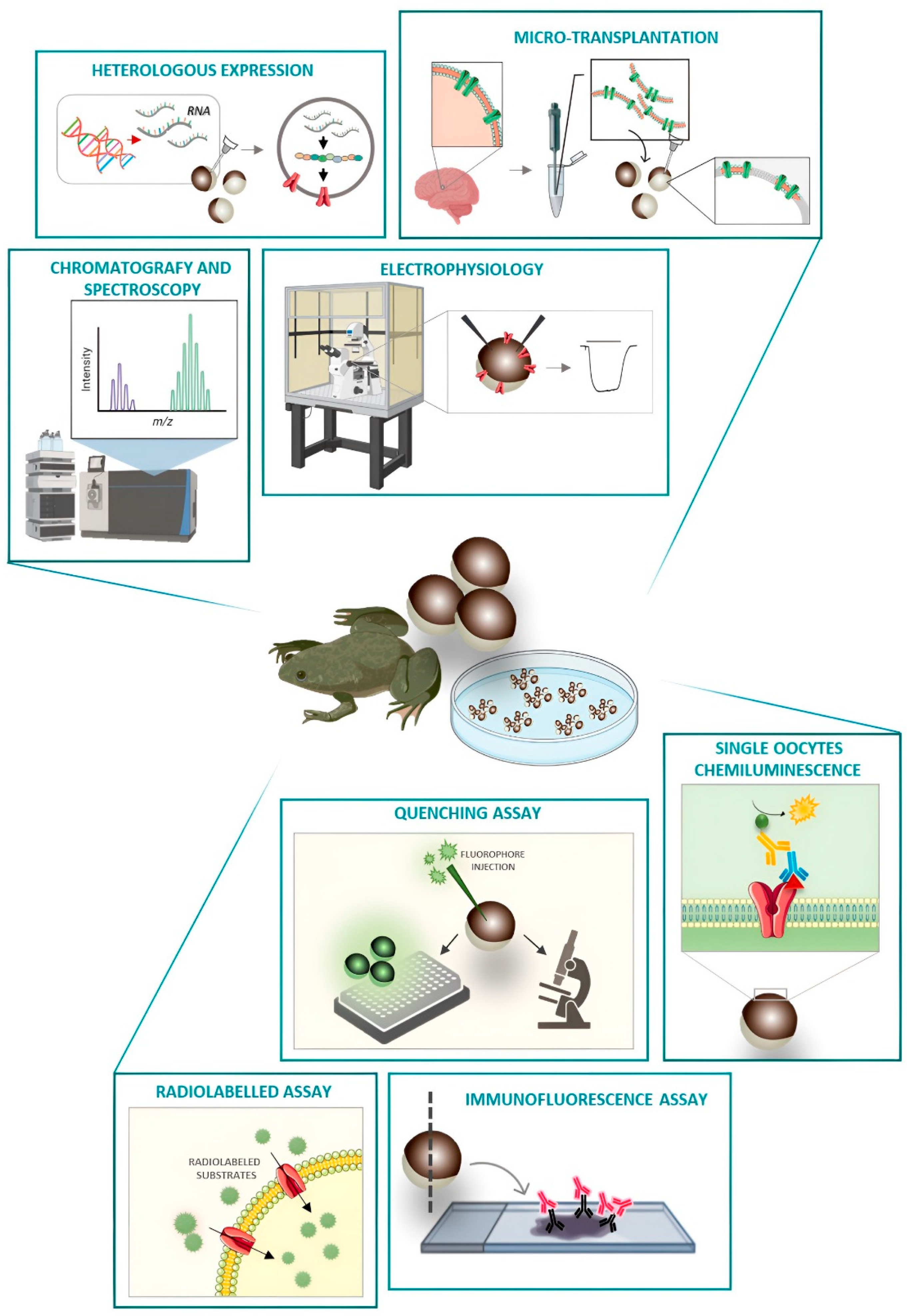
2. Xenopus laevis Oocytes as a Heterologous System of Expression
2.1. mRNA Injection
2.2. Membrane Microtransplantation from Native Tissues
3. Testing the Function of Membrane Proteins
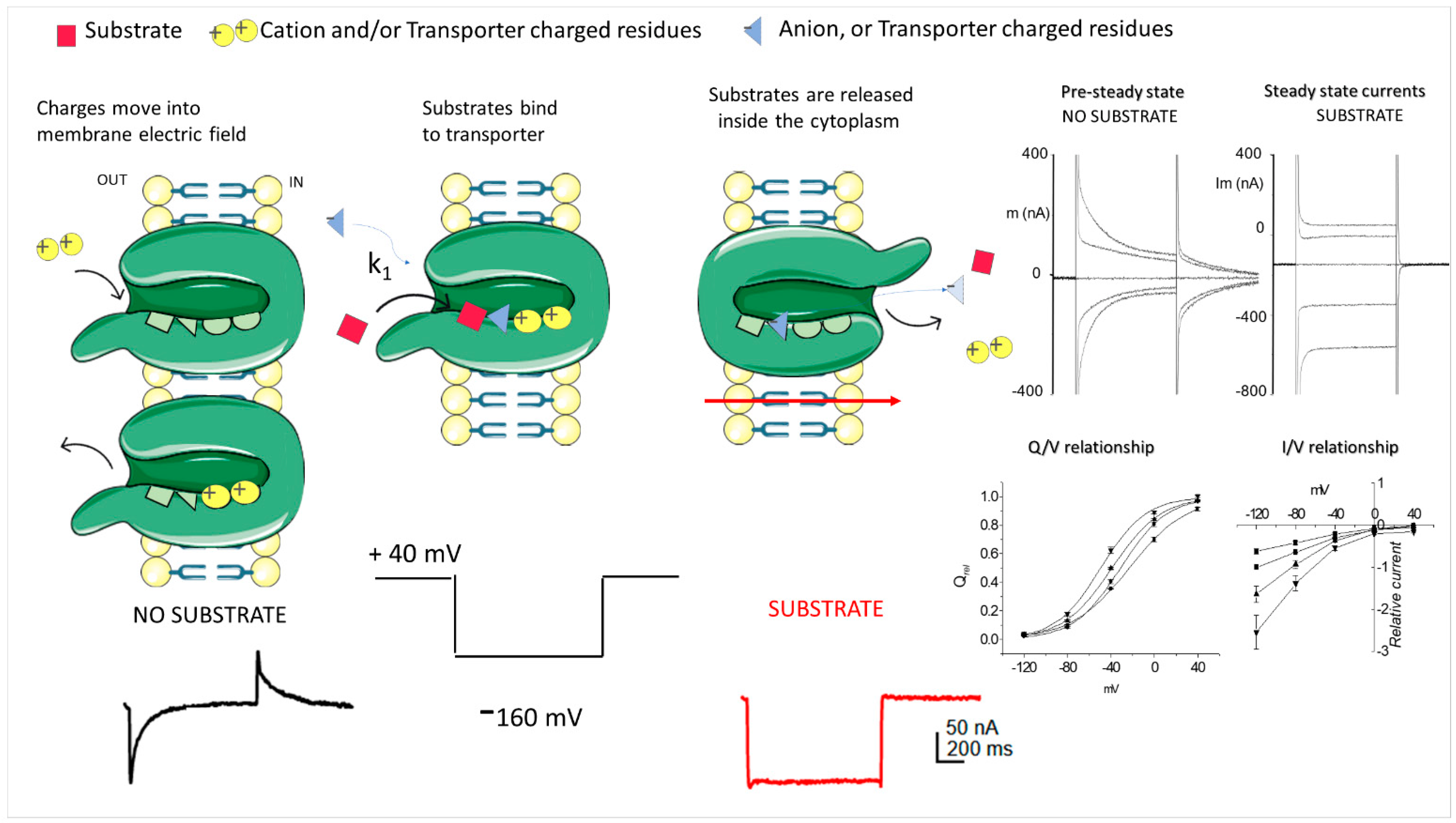
3.1. Classical TEVC
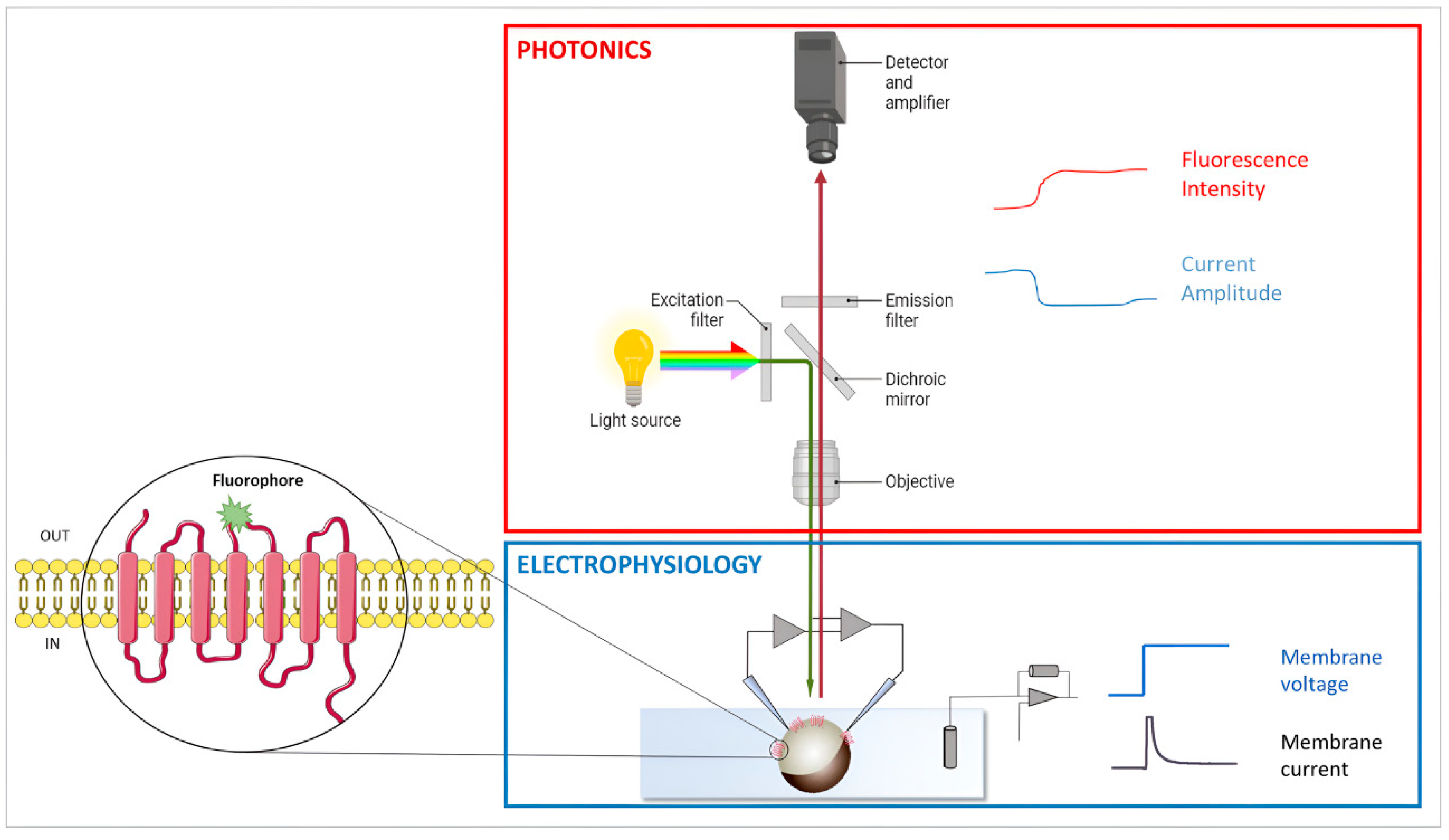
3.2. Voltage Clamp Fluorimetry (VCF)
4. Testing the Expression of Membrane Proteins
SOC and Immunochemistry
5. Alternative Methods to Quantify the Functionality
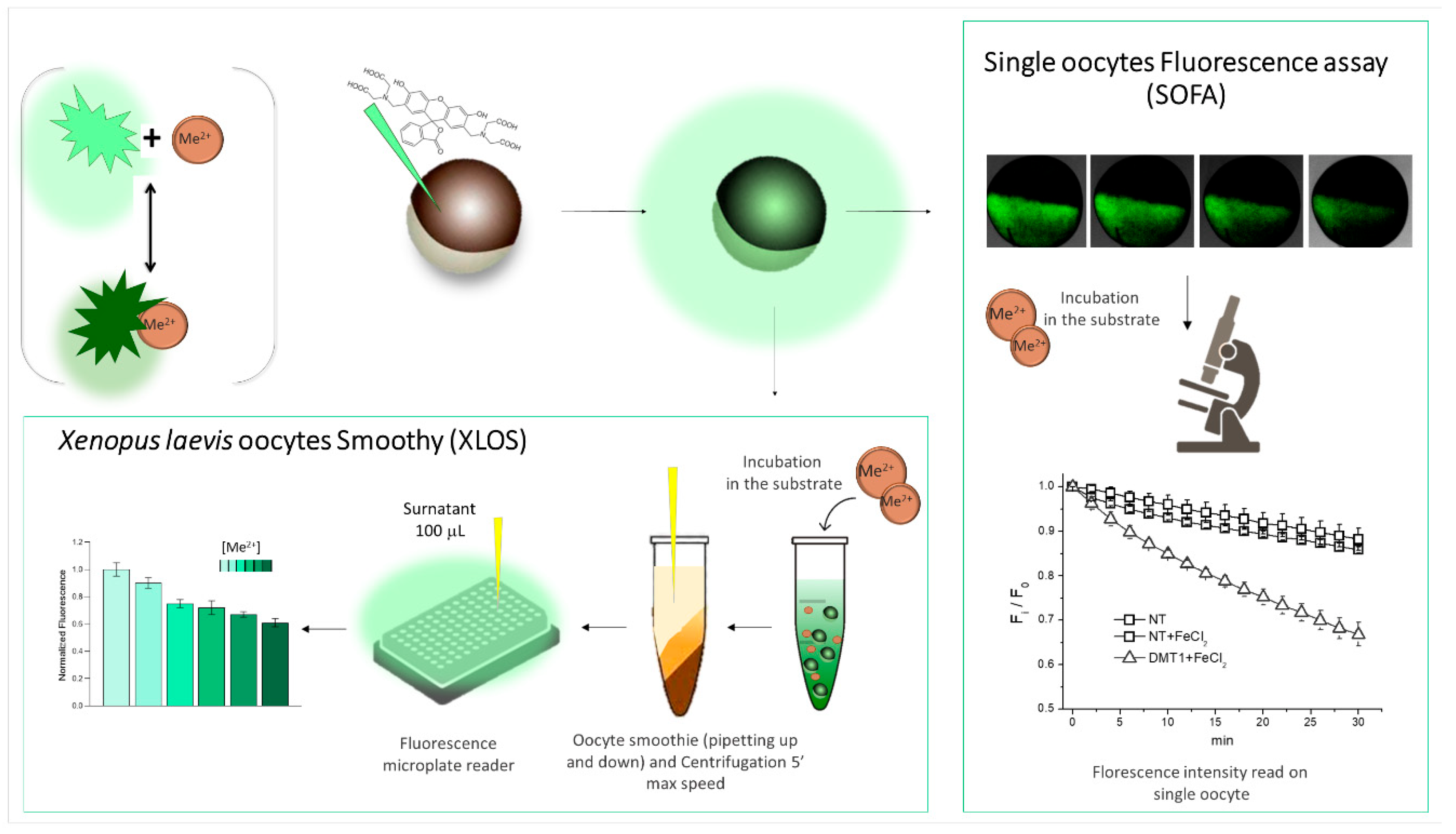
5.1. Fluorescence Monitoring
5.2. Monitoring the Transport by HPLC, GC-MS
5.3. High-Throughput Systems
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
Appendix A
Appendix A.1. Oocytes Collection Procedure
- -
- Anesthetize the frog by immersion in 500 mL of tricaine methane sulfonate, check the pH, and, in case, adjust with bicarbonate solution (750 mg−1g) to pH 8.
- -
- Sterilize the frog’s abdomen with an antiseptic agent (povidone iodine, 0.8%);
- -
- Uplift the skin in the lower abdominal quadrant and make a 10 mm incision using a pair of sterilized, small scissors. This exposes the body wall, which is then cut (0.5 cm) using the same technique.
- -
- Remove the portion of the ovary and collect the oocytes in a tube containing ND96 Ø Ca2+;
- -
- Suture both the incisions in the body wall muscle and the skin using stitches of absorbable synthetic thread.
- -
- Leave the frog for one/two hours post-surgery in a small tank with 3 liters of FETAX solution until complete recovery from anesthesia.
Appendix A.2. Oocytes Preparation Procedure
- -
- Using sterilized tweezers, split the ovary into small pieces: groups of 8–10 oocytes in a Petri dish containing ND96 Ø Ca2+.
- -
- Wash the cluster of oocytes twice in clean ND96 Ø Ca2+ and move the oocytes into a 15 mL tube containing 5 mL of collagenase solution.
- -
- Treat the oocyte groups in the collagenase solution with continuous agitation for about an hour at 18 °C to separate the single oocytes and remove the follicular membrane.
- -
- Wash the oocytes three times with ND96 Ø Ca2+ and three times with NDE solutions.
- -
- Transfer oocytes to a 90 mm Petri dish containing NDE medium solution and incubate for at least 90 min at 18 °C;
- -
- Select the healthy and fully grown oocytes (stage V and VI) under a stereomicroscope and transfer them to a fresh Petri dish containing NDE medium.
Appendix A.3. Selection Criteria
- -
- Well-composed and spherical;
- -
- The distinct partition between animal and vegetal poles;
- -
- Homogenous pigmentation in animal and vegetal poles;
- -
- Absence of follicular membrane.
Appendix A.4. Membrane Tissue Extraction Procedure
- -
- Homogenize the frozen tissue sample (about 0.5 g) using a Teflon glass homogenizer in 2 mL of glycine buffer at pH 9 (200 mM glycine/150 mM NaCl/50 mM EGTA/50 mM EDTA/300 mM sucrose), adding 20 μL of protease inhibitors (Sigma 2714, St. Louis, MO, USA);
- -
- Centrifuge for 15 min at 9500× g at 4 °C (Beckmann centrifuge, C1015 rotor);
- -
- Centrifuge the obtained supernatant for 2 h at 100,000× g a 4 °C (Beckmann centrifuge, SW40 rotor).
- -
- Wash the final pellet with H2O and resuspend it in 5 mM glycine.
- -
- Store the aliquots at −80 °C or use them directly.
- -
- Inject the membrane samples (50–100 nL/ 0.2–10 mg protein/mL) into oocytes using a manual microinjection system (Drummond Scientific Company, Broomall, PA, USA).
Appendix A.5. Expression Vector
- -
- The presence of a promoter site for RNA polymerase (T7 or SP6) upstream of the open reading frame (ORF). It is better to use a T7 or SP6 promoter since the T3 polymer has generally been shown to give rise to an unstable transcript (in our experience, T7 always works better).
- -
- A polyA tail is needed to stabilize the construct.
- -
- A multiple cloning site allows subcloning of the transporter cDNA downstream of the promoter and upstream of the polyA tail.
- -
- It is also important that the untranslated region between the RNA polymerase promoter (T7, T3, or SP6) is as short as possible or has the presence of 5′ and 3′ untranslated regions derived from the Xenopus laevis protein, usually the β-globin, which is efficiently translated in microinjected oocytes.
- -
- The sequences flanking the initiation codon within the cDNA can be modified by site-directed mutagenesis so that they conform to the optimal translation initiation sequence, GCCGCCA/GCCmG. The presence of both a purine at position −3 and a G at the position at +4 is sufficient to direct optimal translation.
- -
- A unique restriction site downstream of the transporter ORF that allows linearization of the transporter. Restriction enzymes that generate blunt ends or 5’ overhangs are preferred.
Appendix A.6. RNA Preparation Procedure
- -
- Transform the recombinant plasmids expressing the gene of interest into the JM109 strain of E. Coli by performing a heat-shock procedure and letting the colony grow at 37 °C overnight.
- -
- Inoculate a single colony in a liquid selective medium specific for plasmid resistance and incubate at 37 °C overnight under constant agitation.
- -
- Extract the plasmid DNAs using Wizard® Plus SV Miniprep (Promega Italia, Milan, Italy).
- -
- To achieve an efficient in vitro transcription, linearize the recombinant plasmid with a restriction enzyme which generates blunt ends or 5’ overhangs.
- -
- Purify with Wizard SV Gel and PCR clean-up system (Promega Italia).
- -
- Compose the in vitro transcription reaction as follow: 18 μL of transcription buffer 5X, 8 μL of DTT 100 mmol/L, 2 μL of RNasin® ribonuclease inhibitors 40 U/μL, 13 μL rNTPs mix (rATP, rCTP, rUTP 5 mmol/L and rGTP 0.5 mmol/L), 6.5 μL of cap analog 10 mmol/L, 10 μL T7 RNA polymerase 20 U/μL (final volume 90 μl). After 10, 20, and 40 min from the beginning of the incubation, add 1 μL of rGTP 25 mmoL/L. After 1 h from the start of the transcription, add a mix of 4 μL of transcription buffer 5X, 1 μL of DTT 100 mmol/L, 1 μL of RNasin® ribonuclease inhibitors 40 U/μL, 5 μL of rNTPs mix, 1 μL of T7 RNA polymerase 20 U/μL, 1 μL of GTP 25 mmoL/L, 4 μL of nuclease-free water. Wait 2 h. Add 101 μL of nuclease-free and 150 μL of LiCl 8 mol/L and store at −80 °C overnight. The next day, precipitate and wash the transcribed cRNA with EtOH 70% to obtain the purified cRNA.
- -
- Quantify the cRNA by NanoDrop™ 2000 Spectrophotometer (Thermo Fisher Scientific, Waltham, MA, USA).
- -
- Store the samples at −80 °C.
- -
- Inject 50 nL of the cRNA (for the solute carrier, the concentration is always between 0.25 and 0.5 µg/µ) in each oocyte using a manual microinjection system (Drummond Scientific Company, Broomall, PA, USA).
Appendix A.7. SOC (Single-Oocyte Chemiluminescence)
- -
- Transfer oocytes (5–10 in each well) into a 24-well plate with 500 μL of ND96;
- -
- Wash using 500 μL ND96 cold in mild agitation (5 min at RT, three times);
- -
- Fix with 500 μL of 4% paraformaldehyde in ND96 and shake the multiwell at 4 °C for 15 min;
- -
- Wash using 500 μL ND96 cold in mild agitation (5 min at RT, three times);
- -
- Block with 500 μL blocking solution and shake at 4 °C for 1 h.
- -
- Incubate oocytes with 300–600 μL of AB I°, diluted, according to the kind of AB at defined concentration, in blocking solution and shake at 4 °C for 1 h.
- -
- Wash using 500 μL blocking solution in mild agitation (5 min at 4 °C, three times).
- -
- Incubate oocytes with 300–600 μL of AB II° conjugated to Horseradish Peroxidase (HPR) diluted, according to the manufacturing indication, in blocking solution and shake at 4 °C for 1 h.
- -
- Wash using 500 μL blocking solution in mild agitation (5 min at 4 °C, three times).
- -
- Wash the oocytes for the last time with ND96 in mild agitation at RT for 10 min.
- -
- In a 96-well costar white flat bottom plate, put 50 μL of SuperSignal (Thermofisher); Transfer the oocytes to the wells with LUMINOL and incubate for 1 h before reading the luminescence in the plate reader.
Appendix A.8. Immunohistochemistry
- -
- Transfer oocytes (5–10 in each well) into a 24-well plate with 500 μL of ND96.
- -
- Wash using 500 μL ND96 cold in mild agitation (5 min at RT, three times);
- -
- Fix with 500 μL of 4% paraformaldehyde in ND96 and shake the multiwell at 4 °C for 15 min.
- -
- Wash using 500 μL ND96 cold in mild agitation (5 min at RT, three times).
- -
- Include the oocyte in Polyfreeze tissue-freezing medium and then freeze in liquid nitrogen.
- -
- Use a cryostat to obtain oocyte cryosections (7 μm thickness) and store at −20 °C until immunostaining.
Appendix A.9. Solutions
- -
- Anesthetic solution: Dissolve 500 mg tricaine (ethyl 3 aminobenzoate methane sulfonate) in 500 mL MilliQ water. Add sodium bicarbonate (750 mg−1 g) to the anesthetic solution until pH > 8;
- -
- FETAX: NaCl 10.69, NaHCO3 1.14, KCl 0.4, CaCl2 0.1, CaSO4 0.35, MgSO4 0.62 (in mM);
- -
- Collagenase solution: dissolve collagenase from Clostridium histolyticum (collagenase type IA from Clostridium histolyticum, C0130 from Sigma-Aldrich) in ND96 Ø Ca2+. Use a concentration of 1–2 mg/mL according to the FALGPA value stated in the production batch specifications.
- -
- ND96Ø: NaCl 96, KCl 2, MgCl2 1, and HEPES 5 (in mM), to pH 7.6 with NaOH.
- -
- ND96: NaCl 96, KCl 2, CaCl2 1.8, MgCl2 1, and HEPES 5 (in mM) to pH 7.6 with NaOH.
- -
- NDE: ND96 with pyruvate 2.5 mM, and 50 μg/mL gentamycin sulphate.
- -
- Blocking solution: 50 mL ND96 + 0.5 g BSA.
References
- Dascal, N.; Lotan, I. Expression of exogenus ion channel and neurotrasmitter receptor in RNA-injected Xenopus oocytes. In Methods in Molecular Biology, Vol 13: Protocols in Molecular Neurobiology; Bausch, S., Prevest, A., Eds.; The Humana Press: Totowa NJ, USA, 1992; Volume 13, pp. 205–225. [Google Scholar]
- Richter, J.D.; Smith, L.D. Differential capacity for translation and lack of competition between mRNAs that segregate to free and membrane-bound polysomes. Cell 1981, 27, 183–191. [Google Scholar] [CrossRef]
- Meric, F.; Searfoss, A.M.; Wormington, M.; Wolffe, A.P. Masking and unmasking maternal mRNA: The role of polyadenylation, transcription, splicing, and nuclear history. J. Biol. Chem. 1996, 271, 30804–30810. [Google Scholar] [CrossRef]
- Kusano, K.; Miledi, R.; Stinnakre, J. Cholinergic and catecholaminergic receptors in the Xenopus oocyte membrane. J. Physiol. 1982, 328, 143–170. [Google Scholar] [CrossRef] [PubMed]
- Terhag, J.; Cavara, N.A.; Hollmann, M. Cave Canalem: How endogenous ion channels may interfere with heterologous expression in Xenopus oocytes. Methods 2010, 51, 66–74. [Google Scholar] [CrossRef] [PubMed]
- Sobczak, K.; Bangel-Ruland, N.; Leier, G.; Weber, W.M. Endogenous transport systems in the Xenopus laevis oocyte plasma membrane. Methods 2010, 51, 183–189. [Google Scholar] [CrossRef]
- Tosco, M.; Faelli, A.; Gastaldi, G.; Paulmichl, M.; Orsenigo, M.N. Endogenous lactate transport in Xenopus laevis oocyte: Dependence on cytoskeleton and regulation by protein kinases. J. Comp. Physiol. B 2008, 178, 457–463. [Google Scholar] [CrossRef]
- Tzounopoulos, T.; Maylie, J.; Adelman, J.P. Induction of endogenous channels by high levels of heterologous membrane proteins in Xenopus oocytes. Biophys. J. 1995, 69, 904–908. [Google Scholar] [CrossRef]
- Van Winkle, L.J. Endogenous amino acid transport systems and expression of mammalian amino acid transport proteins in Xenopus oocytes. Biochim. Biophys. Acta 1993, 1154, 157–172. [Google Scholar] [CrossRef]
- Krafte, D.S.; Volberg, W.A. Properties of endogenous voltage-dependent sodium currents in Xenopus laevis oocytes. J. Neurosci. Methods 1992, 43, 189–193. [Google Scholar] [CrossRef]
- Bossi, E.; Centinaio, E.; Moriondo, A.; Peres, A. Ca2+-dependence of the depolarization-inducible Na+ current of Xenopus oocytes. J. Cell Physiol. 1998, 174, 154–159. [Google Scholar] [CrossRef]
- Centinaio, E.; Bossi, E.; Peres, A. Properties of the Ca(2+)-activated Cl- current of Xenopus oocytes. Cell Mol. Life Sci. 1997, 53, 604–610. [Google Scholar] [CrossRef] [PubMed]
- Weber, W.M. Endogenous ion channels in oocytes of xenopus laevis: Recent developments. J. Membr. Biol. 1999, 170, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, T.; Ikeda, K.; Kumanishi, T. Functional characterization of an endogenous Xenopus oocyte adenosine receptor. Br. J. Pharmacol. 2002, 135, 313–322. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.L.; Hoshi, T.; Wu, C.F. Heteromultimeric Interaction among K+ Channel Subunits from Shaker and eag Families in Xenopus Oocytes. Neuron 1996, 17, 535–542. [Google Scholar] [CrossRef]
- Wongsamitkul, N.; Nache, V.; Eick, T.; Hummert, S.; Schulz, E.; Schmauder, R.; Schirmeyer, J.; Zimmer, T.; Benndorf, K. Quantifying the cooperative subunit action in a multimeric membrane receptor. Sci. Rep. 2016, 6, 20974. [Google Scholar] [CrossRef] [PubMed]
- Isacoff, E.Y.; Jan, Y.N.; Jan, L.Y. Evidence for the formation of heteromultimeric potassium channels in Xenopus oocytes. Nature 1990, 345, 530–534. [Google Scholar] [CrossRef]
- Shistik, E.; Ivanina, T.; Puri, T.; Hosey, M.; Dascal, N. Ca2+ current enhancement by alpha 2/delta and beta subunits in Xenopus oocytes: Contribution of changes in channel gating and alpha 1 protein level. J. Physiol. 1995, 489 Pt 1, 55–62. [Google Scholar] [CrossRef] [PubMed]
- Bröer, S.; Rahman, B.; Pellegri, G.; Pellerin, L.; Martin, J.-L.; Verleysdonk, S.; Hamprecht, B.; Magistretti, P.J. Comparison of Lactate Transport in Astroglial Cells and Monocarboxylate Transporter 1 (MCT 1) Expressing Xenopus laevis Oocytes: EXPRESSION OF TWO DIFFERENT MONOCARBOXYLATE TRANSPORTERS IN ASTROGLIAL CELLS AND NEURONS*. J. Biol. Chem. 1997, 272, 30096–30102. [Google Scholar] [CrossRef] [PubMed]
- Niu, L.; Vazquez, R.W.; Nagel, G.; Friedrich, T.; Bamberg, E.; Oswald, R.E.; Hess, G.P. Rapid chemical kinetic techniques for investigations of neurotransmitter receptors expressed in Xenopus oocytes. Proc. Natl. Acad. Sci. USA 1996, 93, 12964–12968. [Google Scholar] [CrossRef]
- Yang, J.; Cheng, Q.; Takahashi, A.; Goubaeva, F. Kinetic properties of GABA rho1 homomeric receptors expressed in HEK293 cells. Biophys. J. 2006, 91, 2155–2162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keller, K.; Strube, M.; Mueckler, M. Functional expression of the human HepG2 and rat adipocyte glucose transporters in Xenopus oocytes. Comparison of kinetic parameters. J. Biol. Chem. 1989, 264, 18884–18889. [Google Scholar] [CrossRef]
- Gurdon, J.B.; Lingrel, J.B.; Marbaix, G. Message stability in injected frog oocytes: Long life of mammalian alpha and beta globin messages. J. Mol. Biol. 1973, 80, 539–551. [Google Scholar] [CrossRef]
- Gurdon, J.B.; Lane, C.D.; Woodland, H.R.; Marbaix, G. Use of frog eggs and oocytes for the study of messenger RNA and its translation in living cells. Nature 1971, 233, 177–182. [Google Scholar] [CrossRef] [PubMed]
- Gurdon, J.B. The translation of messenger RNA injected in living oocytes of Xenopus laevis. Acta Endocrinol. Suppl. 1973, 180, 225–243. [Google Scholar] [CrossRef]
- Lane, C.D.; Marbaix, G.; Gurdon, J.B. Rabbit haemoglobin synthesis in frog cells: The translation of reticulocyte 9 s RNA in frog oocytes. J. Mol. Biol. 1971, 61, 73–91. [Google Scholar] [CrossRef]
- Sumikawa, K.; Houghton, M.; Emtage, J.S.; Richards, B.M.; Barnard, E.A. Active multi-subunit ACh receptor assembled by translation of heterologous mRNA in Xenopus oocytes. Nature 1981, 292, 862–864. [Google Scholar] [CrossRef]
- Miledi, R.; Parker, I.; Sumikawa, K. Properties of acetylcholine receptors translated by cat muscle mRNA in Xenopus oocytes. Embo J. 1982, 1, 1307–1312. [Google Scholar] [CrossRef]
- Barnard, E.A.; Miledi, R.; Sumikawa, K. Translation of exogenous messenger RNA coding for nicotinic acetylcholine receptors produces functional receptors in Xenopus oocytes. Proc. R. Soc. Lond. B. Biol. Sci. 1982, 215, 241–246. [Google Scholar] [CrossRef]
- Miledi, R.; Parker, I.; Sumikawa, K. Synthesis of chick brain GABA receptors by frog oocytes. Proc. R. Soc. Lond. B. Biol. Sci. 1982, 216, 509–515. [Google Scholar] [CrossRef]
- Nargeot, J.; Dascal, N.; Lester, H.A. Heterologous expression of calcium channels. J. Membr. Biol. 1992, 126, 97–108. [Google Scholar] [CrossRef]
- Miledi, R.; Eusebi, F.; Martinez-Torres, A.; Palma, E.; Trettel, F. Expression of functional neurotransmitter receptors in Xenopus oocytes after injection of human brain membranes. Proc. Natl. Acad. Sci. USA 2002, 99, 13238–13242. [Google Scholar] [CrossRef] [PubMed]
- Eusebi, F.; Palma, E.; Amici, M.; Miledi, R. Microtransplantation of ligand-gated receptor-channels from fresh or frozen nervous tissue into Xenopus oocytes: A potent tool for expanding functional information. Prog. Neurobiol. 2009, 88, 32–40. [Google Scholar] [CrossRef] [PubMed]
- Miledi, R.; Palma, E.; Eusebi, F. Microtransplantation of neurotransmitter receptors from cells to Xenopus oocyte membranes: New procedure for ion channel studies. Methods Mol. Biol. 2006, 322, 347–355. [Google Scholar] [CrossRef] [PubMed]
- Palma, E.; Trettel, F.; Fucile, S.; Renzi, M.; Miledi, R.; Eusebi, F. Microtransplantation of membranes from cultured cells to Xenopus oocytes: A method to study neurotransmitter receptors embedded in native lipids. Proc. Natl. Acad. Sci. USA 2003, 100, 2896–2900. [Google Scholar] [CrossRef]
- Mazzo, F.; Zwart, R.; Serratto, G.M.; Gardinier, K.M.; Porter, W.; Reel, J.; Maraula, G.; Sher, E. Reconstitution of synaptic Ion channels from rodent and human brain in Xenopus oocytes: A biochemical and electrophysiological characterization. J. Neurochem. 2016, 138, 384–396. [Google Scholar] [CrossRef]
- Ruffolo, G.; Cifelli, P.; Roseti, C.; Thom, M.; van Vliet, E.A.; Limatola, C.; Aronica, E.; Palma, E. A novel GABAergic dysfunction in human Dravet syndrome. Epilepsia 2018, 59, 2106–2117. [Google Scholar] [CrossRef] [PubMed]
- Roseti, C.; Fucile, S.; Lauro, C.; Martinello, K.; Bertollini, C.; Esposito, V.; Mascia, A.; Catalano, M.; Aronica, E.; Limatola, C.; et al. Fractalkine/CX3CL1 modulates GABAA currents in human temporal lobe epilepsy. Epilepsia 2013, 54, 1834–1844. [Google Scholar] [CrossRef]
- Miledi, R.; Duenas, Z.; Martinez-Torres, A.; Kawas, C.H.; Eusebi, F. Microtransplantation of functional receptors and channels from the Alzheimer’s brain to frog oocytes. Proc. Natl. Acad. Sci. USA 2004, 101, 1760–1763. [Google Scholar] [CrossRef]
- Zwart, R.; Mazzo, F.; Sher, E. Microtransplantation of human brain receptors into oocytes to tackle key questions in drug discovery. Drug. Discov. Today 2019, 24, 533–543. [Google Scholar] [CrossRef]
- Ruffolo, G.; Di Bonaventura, C.; Cifelli, P.; Roseti, C.; Fattouch, J.; Morano, A.; Limatola, C.; Aronica, E.; Palma, E.; Giallonardo, A.T. A novel action of lacosamide on GABAA currents sets the ground for a synergic interaction with levetiracetam in treatment of epilepsy. NeuroBiol. Dis. 2018, 115, 59–68. [Google Scholar] [CrossRef]
- Roseti, C.; van Vliet, E.A.; Cifelli, P.; Ruffolo, G.; Baayen, J.C.; Di Castro, M.A.; Bertollini, C.; Limatola, C.; Aronica, E.; Vezzani, A.; et al. GABAA currents are decreased by IL-1beta in epileptogenic tissue of patients with temporal lobe epilepsy: Implications for ictogenesis. NeuroBiol. Dis. 2015, 82, 311–320. [Google Scholar] [CrossRef] [PubMed]
- Roseti, C.; Cifelli, P.; Ruffolo, G.; Barbieri, E.; Guescini, M.; Esposito, V.; Di Gennaro, G.; Limatola, C.; Giovannelli, A.; Aronica, E.; et al. Erythropoietin Increases GABA(A) Currents in Human Cortex from TLE Patients. Neuroscience 2020, 439, 153–162. [Google Scholar] [CrossRef] [PubMed]
- Ruffolo, G.; Cifelli, P.; Miranda-Lourenço, C.; De Felice, E.; Limatola, C.; Sebastião, A.M.; Diógenes, M.J.; Aronica, E.; Palma, E. Rare Diseases of Neurodevelopment: Maintain the Mystery or Use a Dazzling Tool for Investigation? The Case of Rett Syndrome. Neuroscience 2020, 439, 146–152. [Google Scholar] [CrossRef] [PubMed]
- Palma, E.; Reyes-Ruiz, J.M.; Lopergolo, D.; Roseti, C.; Bertollini, C.; Ruffolo, G.; Cifelli, P.; Onesti, E.; Limatola, C.; Miledi, R.; et al. Acetylcholine receptors from human muscle as pharmacological targets for ALS therapy. Proc. Natl. Acad. Sci. USA 2016, 113, 3060–3065. [Google Scholar] [CrossRef]
- Giovannardi, S.; Soragna, A.; Magagnin, S.; Faravelli, L. Functional expression of type 1 rat GABA transporter in microinjected Xenopus laevis oocytes. Methods Mol. Biol. 2007, 375, 235–255. [Google Scholar] [CrossRef]
- Dascal, N. Voltage clamp recordings from Xenopus oocytes. Curr. Protoc. Neurosci. 2001, 10, 6–12. [Google Scholar] [CrossRef]
- Guan, B.; Chen, X.; Zhang, H. Two-electrode voltage clamp. Methods Mol. Biol. 2013, 998, 79–89. [Google Scholar] [CrossRef] [PubMed]
- Hediger, M.A.; Coady, M.J.; Ikeda, T.S.; Wright, E.M. Expression cloning and cDNA sequencing of the Na+/glucose co-transporter. Nature 1987, 330, 379–381. [Google Scholar] [CrossRef]
- Blakely, R.D.; Robinson, M.B.; Amara, S.G. Expression of neurotransmitter transport from rat brain mRNA in Xenopus laevis oocytes. Proc. Natl. Acad. Sci. USA 1988, 85, 9846–9850. [Google Scholar] [CrossRef]
- Romero, M.F.; Kanai, Y.; Gunshin, H.; Hediger, M.A. Expression cloning using Xenopus laevis oocytes. Methods Enzym. 1998, 296, 17–52. [Google Scholar] [CrossRef]
- Markovich, D. Expression cloning and radiotracer uptakes in Xenopus laevis oocytes. Nature Protoc. 2008, 3, 1975–1980. [Google Scholar] [CrossRef]
- Cesar-Razquin, A.; Snijder, B.; Frappier-Brinton, T.; Isserlin, R.; Gyimesi, G.; Bai, X.; Reithmeier, R.A.; Hepworth, D.; Hediger, M.A.; Edwards, A.M.; et al. A Call for Systematic Research on Solute Carriers. Cell 2015, 162, 478–487. [Google Scholar] [CrossRef]
- Superti-Furga, G.; Lackner, D.; Wiedmer, T.; Ingles-Prieto, A.; Barbosa, B.; Girardi, E.; Goldmann, U.; Gurtl, B.; Klavins, K.; Klimek, C.; et al. The RESOLUTE consortium: Unlocking SLC transporters for drug discovery. Nature Rev. Drug Discov. 2020, 19, 429–430. [Google Scholar] [CrossRef]
- Wang, W.W.; Gallo, L.; Jadhav, A.; Hawkins, R.; Parker, C.G. The Druggability of Solute Carriers. J. Med. Chem. 2020, 63, 3834–3867. [Google Scholar] [CrossRef]
- Kanner, B.I. Transmembrane domain I of the gamma-aminobutyric acid transporter GAT-1 plays a crucial role in the transition between cation leak and transport modes. J. Biol. Chem. 2003, 278, 3705–3712. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.G.; Bennett, E.R.; Kanner, B.I. The aqueous accessibility in the external half of transmembrane domain I of the GABA transporter GAT-1 is modulated by its Ligands. J. Biol. Chem. 2004, 279, 13800–13808. [Google Scholar] [CrossRef] [PubMed]
- Yamashita, A.; Singh, S.K.; Kawate, T.; Jin, Y.; Gouaux, E. Crystal structure of a bacterial homologue of Na+/Cl-dependent neurotransmitter transporters. Nature 2005, 437, 215–223. [Google Scholar] [CrossRef]
- Barker, E.L.; Blakely, R.D. Structural Determinants of Neurotransmitter Transport using Cross-Species Chimeras: Studies on Serotonine Transporter. Methods Enzymol. 1998, 296, 475–498. [Google Scholar]
- Buck, K.J.; Amara, S.G. In Vivo Generation of chimeras. Methods Enzymol. 1998, 296, 466–475. [Google Scholar] [PubMed]
- Soragna, A.; Mari, S.A.; Pisani, R.; Peres, A.; Castagna, M.; Sacchi, V.F.; Bossi, E. Structural domains involved in substrate selectivity in two neutral amino acid transporters. Am. J. Physiol. Cell Physiol. 2004, 287, C754–C761. [Google Scholar] [CrossRef]
- Buck, K.J.; Amara, S.G. Chimeric dopamine-norepinephrine transporters delineate structural domains influencing selectivity for catecholamines and 1-methyl-4-phenylpyridinium. Proc. Natl. Acad. Sci. USA 1994, 91, 12584–12588. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buck, K.J.; Amara, S.G. Structural domains of catecholamine transporter chimeras involved in selective inhibition by antidepressants and psychomotor stimulants. Mol. Pharmacol. 1995, 48, 1030–1037. [Google Scholar] [PubMed]
- Buracco, S.; Peracino, B.; Cinquetti, R.; Signoretto, E.; Vollero, A.; Imperiali, F.; Castagna, M.; Bossi, E.; Bozzaro, S. Dictyostelium Nramp1, which is structurally and functionally similar to mammalian DMT1 transporter, mediates phagosomal iron efflux. J. Cell Sci. 2015, 128, 3304–3316. [Google Scholar] [CrossRef] [PubMed]
- Sacher, A.; Cohen, A.; Nelson, N. Properties of the mammalian and yeast metal-ion transporters DCT1 and Smf1p expressed in Xenopus laevis oocytes. J. Exp. Biol. 2001, 204, 1053–1061. [Google Scholar] [CrossRef] [PubMed]
- Bacconi, A.; Virkki, L.V.; Biber, J.; Murer, H.; Forster, I.C. Renouncing electroneutrality is not free of charge: Switching on electrogenicity in a Na+-coupled phosphate cotransporter. Proc. Natl. Acad. Sci. USA 2005, 102, 12606–12611. [Google Scholar] [CrossRef] [PubMed]
- Castagna, M.; Cinquetti, R.; Verri, T.; Vacca, F.; Giovanola, M.; Barca, A.; Romanazzi, T.; Roseti, C.; Galli, A.; Bossi, E. The Lepidopteran KAAT1 and CAATCH1: Orthologs to Understand Structure-Function Relationships in Mammalian SLC6 Transporters. Neurochem. Res. 2022, 47, 111–126. [Google Scholar] [CrossRef] [PubMed]
- Margheritis, E.; Terova, G.; Oyadeyi, A.S.; Renna, M.D.; Cinquetti, R.; Peres, A.; Bossi, E. Characterization of the transport of lysine-containing dipeptides by PepT1 orthologs expressed in Xenopus laevis oocytes. Comp. Biochem. Physiology. Part A Mol. Integr. Physiol. 2013, 164, 520–528. [Google Scholar] [CrossRef] [PubMed]
- Giovanola, M.; D’Antoni, F.; Santacroce, M.; Mari, S.A.; Cherubino, F.; Bossi, E.; Sacchi, V.F.; Castagna, M. Role of a conserved glycine triplet in the NSS amino acid transporter KAAT1. Biochim. Biophys. Acta 2012, 1818, 1737–1744. [Google Scholar] [CrossRef] [PubMed]
- Castagna, M.; Bossi, E.; Sacchi, V.F. Molecular physiology of the insect K-activated amino acid transporter 1 (KAAT1) and cation-anion activated amino acid transporter/channel 1 (CAATCH1) in the light of the structure of the homologous protein LeuT. Insect Mol. Biol. 2009, 18, 265–279. [Google Scholar] [CrossRef]
- Giovanola, M.; Vollero, A.; Cinquetti, R.; Bossi, E.; Forrest, L.R.; Di Cairano, E.S.; Castagna, M. Threonine 67 is a key component in the coupling of the NSS amino acid transporter KAAT1. Biochim. Biophys. Acta Biomembr. 2018, 1860, 1179–1186. [Google Scholar] [CrossRef]
- Mari, S.A.; Soragna, A.; Castagna, M.; Bossi, E.; Peres, A.; Sacchi, V.F. Aspartate 338 contributes to the cationic specificity and to driver-amino acid coupling in the insect cotransporter KAAT1. Cell Mol. Life Sci. 2004, 61, 243–256. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Li, M.; Mager, S.; Lester, H.A. Amino acid residues that control pH modulation of transport-associated current in mammalian serotonin transporters. J. Neurosci. 1998, 18, 7739–7749. [Google Scholar] [CrossRef] [PubMed]
- Kohler, K.; Forster, I.C.; Stange, G.; Biber, J.; Murer, H. Essential cysteine residues of the type IIa Na+/Pi cotransporter. Pflug. Arch. 2003, 446, 203–210. [Google Scholar] [CrossRef] [PubMed]
- Ehnes, C.; Forster, I.C.; Bacconi, A.; Kohler, K.; Biber, J.; Murer, H. Structure-function relations of the first and fourth extracellular linkers of the type IIa Na+/Pi cotransporter: II. Substrate interaction and voltage dependency of two functionally important sites. J. Gen. Physiol. 2004, 124, 489–503. [Google Scholar] [CrossRef] [PubMed]
- Ehnes, C.; Forster, I.C.; Kohler, K.; Bacconi, A.; Stange, G.; Biber, J.; Murer, H. Structure-function relations of the first and fourth predicted extracellular linkers of the type IIa Na+/Pi cotransporter: I. Cysteine scanning mutagenesis. J. Gen. Physiol. 2004, 124, 475–488. [Google Scholar] [CrossRef]
- Miszner, A.; Peres, A.; Castagna, M.; Bette, S.; Giovannardi, S.; Cherubino, F.; Bossi, E. Structural and functional basis of amino acid specificity in the invertebrate cotransporter KAAT1. J. Physiol. 2007, 581, 899–913. [Google Scholar] [CrossRef]
- Vandenberg, R.J.; Shaddick, K.; Ju, P. Molecular basis for substrate discrimination by glycine transporters. J. Biol. Chem. 2007, 282, 14447–14453. [Google Scholar] [CrossRef]
- Ben Yona, A.; Bendahan, A.; Kanner, B.I. A glutamine residue conserved in the neurotransmitter:Sodium:Symporters is essential for the interaction of chloride with the GABA transporter GAT-1. J. Biol. Chem. 2010, 286, 2826–2833. [Google Scholar] [CrossRef]
- Forrest, L.R.; Tavoulari, S.; Zhang, Y.W.; Rudnick, G.; Honig, B. Identification of a chloride ion binding site in Na+/Cl-dependent transporters. Proc. Natl. Acad. Sci. USA 2007, 104, 12761–12766. [Google Scholar] [CrossRef]
- Mari, S.A.; Soragna, A.; Castagna, M.; Santacroce, M.; Perego, C.; Bossi, E.; Peres, A.; Sacchi, V.F. Role of the conserved glutamine 291 in the rat gamma-aminobutyric acid transporter rGAT-1. Cell Mol. Life Sci. 2006, 63, 100–111. [Google Scholar] [CrossRef]
- Perez-Siles, G.; Nunez, E.; Morreale, A.; Jimenez, E.; Leo-Macias, A.; Pita, G.; Cherubino, F.; Sangaletti, R.; Bossi, E.; Ortiz, A.R.; et al. An aspartate residue in the external vestibule of glycine transporter 2 (GLYT2) controls cation access and transport coupling. Biochem. J. 2011, 442, 323–334. [Google Scholar] [CrossRef] [Green Version]
- Edelheit, O.; Hanukoglu, A.; Hanukoglu, I. Simple and efficient site-directed mutagenesis using two single-primer reactions in parallel to generate mutants for protein structure-function studies. BMC Biotechnol. 2009, 9, 61. [Google Scholar] [CrossRef] [PubMed]
- Ishii, T.M.; Zerr, P.; Xia, X.-M.; Bond, C.T.; Maylie, J.; Adelman, J.P. [4] Site-directed mutagenesis. In Methods in Enzymology; Academic Press: Cambridge, MA, USA, 1998; Volume 293, pp. 53–71. [Google Scholar]
- Forlani, G.; Bossi, E.; Ghirardelli, R.; Giovannardi, S.; Binda, F.; Bonadiman, L.; Ielmini, L.; Peres, A. Mutation K448E in the external loop 5 of rat GABA transporter rGAT1 induces pH sensitivity and alters substrate interactions. J. Physiol. 2001, 536, 479–494. [Google Scholar] [CrossRef]
- Mager, S.; Naeve, J.; Quick, M.; Labarca, C.; Davidson, N.; Lester, H.A. Steady states, charge movements, and rates for a cloned GABA transporter expressed in Xenopus oocytes. Neuron 1993, 10, 177–188. [Google Scholar] [CrossRef]
- Lester, H.A.; Cao, Y.; Mager, S. Listening to neurotransmitter transporters. Neuron 1996, 17, 807–810. [Google Scholar] [CrossRef]
- Mager, S.; KleinbergerDoron, N.; Keshet, G.I.; Davidson, N.; Kanner, B.I.; Lester, H.A. Ion binding and permeation at the GABA transporter GAT1. J. Neurosci. 1996, 16, 5405–5414. [Google Scholar] [CrossRef]
- Parent, L.; Supplisson, S.; Loo, D.D.; Wright, E.M. Electrogenic properties of the cloned Na+/glucose cotransporter: II—A transport model under nonrapid equilibrium conditions. J. Membr. Biol. 1992, 125, 63–79. [Google Scholar] [CrossRef]
- Sacher, A.; Nelson, N.; Ogi, J.T.; Wright, E.M.; Loo, D.D.; Eskandari, S. Pre-Steady state and Steady-State Kinetics and Turnover Rate of Mouse g-Aminobutyric Acid Transporter (mGAT3). J. Membr. Biol. 2002, 190, 57–73. [Google Scholar] [CrossRef]
- Sala-Rabanal, M.; Loo, D.D.; Hirayama, B.A.; Wright, E.M. Molecular mechanism of dipeptide and drug transport by the human renal H+/oligopeptide cotransporter hPEPT2. Am. J. Physiol. Ren. Physiol. 2008, 294, F1422–F1432. [Google Scholar] [CrossRef]
- Binda, F.; Bossi, E.; Giovannardi, S.; Forlani, G.; Peres, A. Temperature effects on the presteady-state and transport-associated currents of GABA cotransporter rGAT1. FEBS Lett. 2002, 512, 303–307. [Google Scholar] [CrossRef]
- Peres, A.; Giovannardi, S.; Bossi, E.; Fesce, R. Electrophysiological insights into the mechanism of ion-coupled cotransporters. N. Physiol. Sci. 2004, 19, 80–84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Renna, M.D.; Sangaletti, R.; Bossi, E.; Cherubino, F.; Kottra, G.; Peres, A. Unified modeling of the mammalian and fish proton-dependent oligopeptide transporter PepT1. Channels 2011, 5, 89–99. [Google Scholar] [CrossRef] [PubMed]
- Bossi, E.; Cherubino, F.; Margheritis, E.; Oyadeyi, A.S.; Vollero, A.; Peres, A. Temperature effects on the kinetic properties of the rabbit intestinal oligopeptide cotransporter PepT1. Pflug. Arch. 2012, 464, 183–191. [Google Scholar] [CrossRef] [PubMed]
- Vacca, F.; Gomes, A.S.; Murashita, K.; Cinquetti, R.; Roseti, C.; Barca, A.; Rønnestad, I.; Verri, T.; Bossi, E. Functional characterization of Atlantic salmon (Salmo salar L.) PepT2 transporters. J. Physiol. 2022, 600, 2377–2400. [Google Scholar] [CrossRef] [PubMed]
- Sangaletti, R.; Terova, G.; Peres, A.; Bossi, E.; Cora, S.; Saroglia, M. Functional expression of the oligopeptide transporter PepT1 from the sea bass (Dicentrarchus labrax). Pflug. Arch. Eur. J. Physiol. 2009, 459, 47–54. [Google Scholar] [CrossRef]
- Fesce, R.; Giovannardi, S.; Binda, F.; Bossi, E.; Peres, A. The relation between charge movement and transport-associated currents in the rat GABA cotransporter rGAT1. J. Physiol. 2002, 545, 739–750. [Google Scholar] [CrossRef] [PubMed]
- Loo, D.D.; Hazama, A.; Supplisson, S.; Turk, E.; Wright, E.M. Relaxation kinetics of the Na+/glucose cotransporter. Proc. Natl. Acad. Sci. USA 1993, 90, 5767–5771. [Google Scholar] [CrossRef]
- Sala-Rabanal, M.; Loo, D.D.; Hirayama, B.A.; Turk, E.; Wright, E.M. Molecular interactions between dipeptides, drugs and the human intestinal H+ -oligopeptide cotransporter hPEPT1. J. Physiol. 2006, 574, 149–166. [Google Scholar] [CrossRef]
- Forster, I.C. The molecular mechanism of SLC34 proteins: Insights from two decades of transport assays and structure-function studies. Pflug. Arch. Eur. J. Physiol. 2019, 471, 15–42. [Google Scholar] [CrossRef] [PubMed]
- Peres, A.; Vollero, A.; Margheritis, E.; D’Antoni, F.; Bossi, E. An Inverse Relationship Links Temperature and Substrate Apparent Affinity in the Ion-Coupled Cotransporters rGAT1 and KAAT1. Int. J. Mol. Sci. 2012, 13, 15565–15574. [Google Scholar] [CrossRef]
- Rizzello, A.; Romano, A.; Kottra, G.; Acierno, R.; Storelli, C.; Verri, T.; Daniel, H.; Maffia, M. Protein cold adaptation strategy via a unique seven-amino acid domain in the icefish (Chionodraco hamatus) PEPT1 transporter. Proc. Natl. Acad. Sci. USA 2013, 110, 7068–7073. [Google Scholar] [CrossRef] [Green Version]
- Bacconi, A.; Ravera, S.; Virkki, L.V.; Murer, H.; Forster, I.C. Temperature dependence of steady-state and presteady-state kinetics of a type IIb Na+/P i cotransporter. J. Membr. Biol. 2007, 215, 81–92. [Google Scholar] [CrossRef]
- Margheritis, E.; Imperiali, F.G.; Cinquetti, R.; Vollero, A.; Terova, G.; Rimoldi, S.; Girardello, R.; Bossi, E. Amino acid transporter B(0)AT1 (slc6a19) and ancillary protein: Impact on function. Pflug. Arch. Eur. J. Physiol. 2016, 468, 1363–1374. [Google Scholar] [CrossRef]
- Fairweather, S.J.; Broer, A.; Subramanian, N.; Tumer, E.; Cheng, Q.; Schmoll, D.; O’Mara, M.L.; Broer, S. Molecular basis for the interaction of the mammalian amino acid transporters B0AT1 and B0AT3 with their ancillary protein collectrin. J. Biol. Chem. 2015, 290, 24308–24325. [Google Scholar] [CrossRef] [PubMed]
- Cherubino, F.; Bertram, S.; Bossi, E.; Peres, A. Pre-steady-state and reverse transport currents in the GABA transporter GAT1. Am J. Physiol. Cell Physiol. 2012, 302, C1096–C1108. [Google Scholar] [CrossRef]
- Bertram, S.; Cherubino, F.; Bossi, E.; Castagna, M.; Peres, A. Gaba reverse transport by the neuronal cotransporter gat1: Influence of internal chloride depletion. Am. J. Physiol. Cell Physiol. 2011, 301, C1064–C1073. [Google Scholar] [CrossRef] [PubMed]
- Kottra, G.; Stamfort, A.; Daniel, H. PEPT1 as a paradigm for membrane carriers that mediate electrogenic bidirectional transport of anionic, cationic, and neutral substrates. J. Biol. Chem. 2002, 277, 32683–32691. [Google Scholar] [CrossRef] [PubMed]
- Song, L.; Mercado, A.; Vazquez, N.; Xie, Q.; Desai, R.; George, A.L., Jr.; Gamba, G.; Mount, D.B. Molecular, functional, and genomic characterization of human KCC2, the neuronal K-Cl cotransporter. Brain Res. Mol. Brain Res. 2002, 103, 91–105. [Google Scholar] [CrossRef]
- Sugiyama, H.; Ito, I.; Hirono, C. A new type of glutamate receptor linked to inositol phospholipid metabolism. Nature 1987, 325, 531–533. [Google Scholar] [CrossRef]
- Pin, J.P.; Waeber, C.; Prezeau, L.; Bockaert, J.; Heinemann, S.F. Alternative splicing generates metabotropic glutamate receptors inducing different patterns of calcium release in Xenopus oocytes. Proc. Natl. Acad. Sci. USA 1992, 89, 10331–10335. [Google Scholar] [CrossRef] [PubMed]
- Lechleiter, J.; Hellmiss, R.; Duerson, K.; Ennulat, D.; David, N.; Clapham, D.; Peralta, E. Distinct sequence elements control the specificity of G protein activation by muscarinic acetylcholine receptor subtypes. Embo J. 1990, 9, 4381–4390. [Google Scholar] [CrossRef]
- Courjaret, R.; Machaca, K. Xenopus Oocyte As a Model System to Study Store-Operated Ca2+ Entry (SOCE). Front. Cell Dev. Biol. 2016, 4, 66. [Google Scholar] [CrossRef]
- Priest, M.; Bezanilla, F. Functional Site-Directed Fluorometry. Adv. Exp. Med. Biol. 2015, 869, 55–76. [Google Scholar] [CrossRef] [PubMed]
- Virkki, L.V.; Murer, H.; Forster, I.C. Voltage clamp fluorometric measurements on a type II Na+-coupled Pi cotransporter: Shedding light on substrate binding order. J. Gen. Physiol. 2006, 127, 539–555. [Google Scholar] [CrossRef]
- Treger, J.S.; Priest, M.F.; Bezanilla, F. Single-molecule fluorimetry and gating currents inspire an improved optical voltage indicator. eLife 2015, 4, e10482. [Google Scholar] [CrossRef] [PubMed]
- Horne, A.J.; Fedida, D. Use of voltage clamp fluorimetry in understanding potassium channel gating: A review of Shaker fluorescence data. Can. J. Physiol. Pharmacol. 2009, 87, 411–418. [Google Scholar] [CrossRef] [PubMed]
- Kalstrup, T.; Blunck, R. Voltage-clamp Fluorometry in Xenopus Oocytes Using Fluorescent Unnatural Amino Acids. J. Vis. Exp. JoVE 2017, 27, e55598. [Google Scholar] [CrossRef] [PubMed]
- Toft-Bertelsen, T.L.; Krizaj, D.; MacAulay, N. When size matters: Transient receptor potential vanilloid 4 channel as a volume-sensor rather than an osmo-sensor. J. Physiol. 2017, 595, 3287–3302. [Google Scholar] [CrossRef]
- Iovino, L.; Giusti, V.; Pischedda, F.; Giusto, E.; Plotegher, N.; Marte, A.; Battisti, I.; Di Iacovo, A.; Marku, A.; Piccoli, G.; et al. Trafficking of the glutamate transporter is impaired in LRRK2-related Parkinson’s disease. Acta Neuropathol. 2022, 144, 81–106. [Google Scholar] [CrossRef]
- Bossi, E.; Renna, M.D.; Sangaletti, R.; D’Antoni, F.; Cherubino, F.; Kottra, G.; Peres, A. Residues R282 and D341 act as electrostatic gates in the proton-dependent oligopeptide transporter PepT1. J. Physiol. 2011, 589, 495–510. [Google Scholar] [CrossRef]
- Lin-Moshier, Y.; Marchant, J.S. A rapid Western blotting protocol for the Xenopus oocyte. Cold Spring Harb. Protoc. 2013, 2013, pdb-prot072793. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blumenstein, Y.; Ivanina, T.; Shistik, E.; Bossi, E.; Peres, A.; Dascal, N. Regulation of cardiac L-type Ca2+ channel by coexpression of G(alpha s) in Xenopus oocytes. FEBS Lett. 1999, 444, 78–84. [Google Scholar] [CrossRef]
- Ivanina, T.; Perets, T.; Thornhill, W.B.; Levin, G.; Dascal, N.; Lotan, I. Phosphorylation by protein kinase A of RCK1 K+ channels expressed in Xenopus oocytes. Biochemistry 1994, 33, 8786–8792. [Google Scholar] [CrossRef] [PubMed]
- Jørgensen, M.E.; Nour-Eldin, H.H.; Halkier, B.A. A Western Blot Protocol for Detection of Proteins Heterologously Expressed in Xenopus laevis Oocytes. Methods Mol. Biol. 2016, 1405, 99–107. [Google Scholar] [CrossRef] [PubMed]
- Schwappach, B.; Zerangue, N.; Jan, Y.N.; Jan, L.Y. Molecular Basis for KATP Assembly: Transmembrane Interactions Mediate Association of a K+ Channel with an ABC Transporter. Neuron 2000, 26, 155–167. [Google Scholar] [CrossRef]
- Mertl, M.; Daniel, H.; Kottra, G. Substrate-induced changes in the density of peptide transporter PEPT1 expressed in Xenopus oocytes. Am. J. Physiol. Cell Physiol. 2008, 295, C1332–C1343. [Google Scholar] [CrossRef]
- Kottra, G.; Spanier, B.; Verri, T.; Daniel, H. Peptide transporter isoforms are discriminated by the fluorophore-conjugated dipeptides beta-Ala- and d-Ala-Lys-N-7-amino-4-methylcoumarin-3-acetic acid. Physiol. Rep. 2013, 1, e00165. [Google Scholar] [CrossRef]
- Illing, A.C.; Shawki, A.; Cunningham, C.L.; Mackenzie, B. Substrate profile and metal-ion selectivity of human divalent metal-ion transporter-1. J. Biol. Chem. 2012, 287, 30485–30496. [Google Scholar] [CrossRef] [PubMed]
- Musa-Aziz, R.; Boron, W.F.; Parker, M.D. Using fluorometry and ion-sensitive microelectrodes to study the functional expression of heterologously-expressed ion channels and transporters in Xenopus oocytes. Methods 2010, 51, 134–145. [Google Scholar] [CrossRef]
- Bossi, E.; Zanella, D.; Gornati, R.; Bernardini, G. Cobalt oxide nanoparticles can enter inside the cells by crossing plasma membranes. Sci. Rep. 2016, 6, 22254. [Google Scholar] [CrossRef] [PubMed]
- Zanella, D.; Bossi, E.; Gornati, R.; Faria, N.; Powell, J.; Bernardini, G. The direct permeation of nanoparticles through the plasma membrane transiently modifies its properties. Biochim. Biophys. Acta Biomembr. 2019, 1861, 182997. [Google Scholar] [CrossRef] [PubMed]
- Zanella, D.; Bossi, E.; Gornati, R.; Bastos, C.; Faria, N.; Bernardini, G. Iron oxide nanoparticles can cross plasma membranes. Sci. Rep. 2017, 7, 11413. [Google Scholar] [CrossRef] [PubMed]
- Cinquetti, R.; Imperiali, F.G.; Bozzaro, S.; Zanella, D.; Vacca, F.; Roseti, C.; Peracino, B.; Castagna, M.; Bossi, E. Characterization of Transport Activity of SLC11 Transporters in Xenopus laevis Oocytes by Fluorophore Quenching. SLAS Discov. 2021, 26, 798–810. [Google Scholar] [CrossRef]
- Lee, E.E.L.; Bezanilla, F. Methodological improvements for fluorescence recordings in Xenopus laevis oocytes. J. Gen. Physiol. 2019, 151, 264–272. [Google Scholar] [CrossRef]
- Meredith, D.; Boyd, C.A. Dipeptide transport characteristics of the apical membrane of rat lung type II pneumocytes. Am. J. Physiol. 1995, 269, L137–L143. [Google Scholar] [CrossRef]
- Fairweather, S.J.; Okada, S.; Gauthier-Coles, G.; Javed, K.; Bröer, A.; Bröer, S. A GC-MS/Single-Cell Method to Evaluate Membrane Transporter Substrate Specificity and Signaling. Front. Mol. Biosci. 2021, 8, 646574. [Google Scholar] [CrossRef] [PubMed]
- Vollero, A.; Imperiali, F.G.; Cinquetti, R.; Margheritis, E.; Peres, A.; Bossi, E. The D-amino acid transport by the invertebrate SLC6 transporters KAAT1 and CAATCH1 from Manduca sexta. Physiol. Rep. 2016, 4, e12691. [Google Scholar] [CrossRef]
- Trumbull, J.D.; Maslana, E.S.; McKenna, D.G.; Nemcek, T.A.; Niforatos, W.; Pan, J.Y.; Parihar, A.S.; Shieh, C.C.; Wilkins, J.A.; Briggs, C.A.; et al. High throughput electrophysiology using a fully automated, multiplexed recording system. Recept. Channels 2003, 9, 19–28. [Google Scholar] [CrossRef]
- Malysz, J.; Grønlien, J.H.; Timmermann, D.B.; Håkerud, M.; Thorin-Hagene, K.; Ween, H.; Trumbull, J.D.; Xiong, Y.; Briggs, C.A.; Ahring, P.K.; et al. Evaluation of α7 Nicotinic Acetylcholine Receptor Agonists and Positive Allosteric Modulators Using the Parallel Oocyte Electrophysiology Test Station. ASSAY Drug Dev. Technol. 2009, 7, 374–390. [Google Scholar] [CrossRef]
- Shieh, C.C.; Trumbull, J.D.; Sarthy, J.F.; McKenna, D.G.; Parihar, A.S.; Zhang, X.F.; Faltynek, C.R.; Gopalakrishnan, M. Automated Parallel Oocyte Electrophysiology Test station (POETs): A screening platform for identification of ligand-gated ion channel modulators. Assay Drug Dev. Technol. 2003, 1, 655–663. [Google Scholar] [CrossRef]
- Barth, D.; Fronius, M. Shear force modulates the activity of acid-sensing ion channels at low pH or in the presence of non-proton ligands. Sci. Rep. 2019, 9, 6781. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liman, E.R.; Tytgat, J.; Hess, P. Subunit stoichiometry of a mammalian K+ channel determined by construction of multimeric cDNAs. Neuron 1992, 9, 861–871. [Google Scholar] [CrossRef]
- Trudeau, M.C.; Warmke, J.W.; Ganetzky, B.; Robertson, G.A. HERG, a human inward rectifier in the voltage-gated potassium channel family. Science 1995, 269, 92–95. [Google Scholar] [CrossRef] [PubMed]
- Nowak, M.W.; Gallivan, J.P.; Silverman, S.K.; Labarca, C.; Daugherty, D.A.; Lester, H.A. In Vivo Incorporation of Unnatural Amino Acids into Ion Channels in Xenopus Oocytes Expression System. Methods Enzymol. 1998, 293, 504–529. [Google Scholar] [PubMed]
- Ludewig, U.; von Wiren, N.; Frommer, W.B. Uniport of NH4+ by the root hair plasma membrane ammonium transporter LeAMT1; 1. J. Biol. Chem. 2002, 277, 13548–13555. [Google Scholar] [CrossRef] [PubMed]
- Fairweather, S.J.; Broer, A.; O’Mara, M.L.; Broer, S. Intestinal Peptidases Form Functional Complexes with Neutral Amino Acid Transporter B0AT1. Biochem. J. 2012, 446, 135–148. [Google Scholar] [CrossRef] [PubMed] [Green Version]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bhatt, M.; Di Iacovo, A.; Romanazzi, T.; Roseti, C.; Cinquetti, R.; Bossi, E. The “www” of Xenopus laevis Oocytes: The Why, When, What of Xenopus laevis Oocytes in Membrane Transporters Research. Membranes 2022, 12, 927. https://doi.org/10.3390/membranes12100927
Bhatt M, Di Iacovo A, Romanazzi T, Roseti C, Cinquetti R, Bossi E. The “www” of Xenopus laevis Oocytes: The Why, When, What of Xenopus laevis Oocytes in Membrane Transporters Research. Membranes. 2022; 12(10):927. https://doi.org/10.3390/membranes12100927
Chicago/Turabian StyleBhatt, Manan, Angela Di Iacovo, Tiziana Romanazzi, Cristina Roseti, Raffaella Cinquetti, and Elena Bossi. 2022. "The “www” of Xenopus laevis Oocytes: The Why, When, What of Xenopus laevis Oocytes in Membrane Transporters Research" Membranes 12, no. 10: 927. https://doi.org/10.3390/membranes12100927
APA StyleBhatt, M., Di Iacovo, A., Romanazzi, T., Roseti, C., Cinquetti, R., & Bossi, E. (2022). The “www” of Xenopus laevis Oocytes: The Why, When, What of Xenopus laevis Oocytes in Membrane Transporters Research. Membranes, 12(10), 927. https://doi.org/10.3390/membranes12100927