

Xenopus Oocytes as a Powerful Cellular Model to Study Foreign Fully-Processed Membrane Proteins

 , ,
, , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Background
2. Advantages and Drawbacks of Using Xenopus Oocytes to the Study of Membrane Proteins
- (a)
- Xenopus oocytes have a continuous and asynchronous development, allowing a year-round use of these cells [1].
- (b)
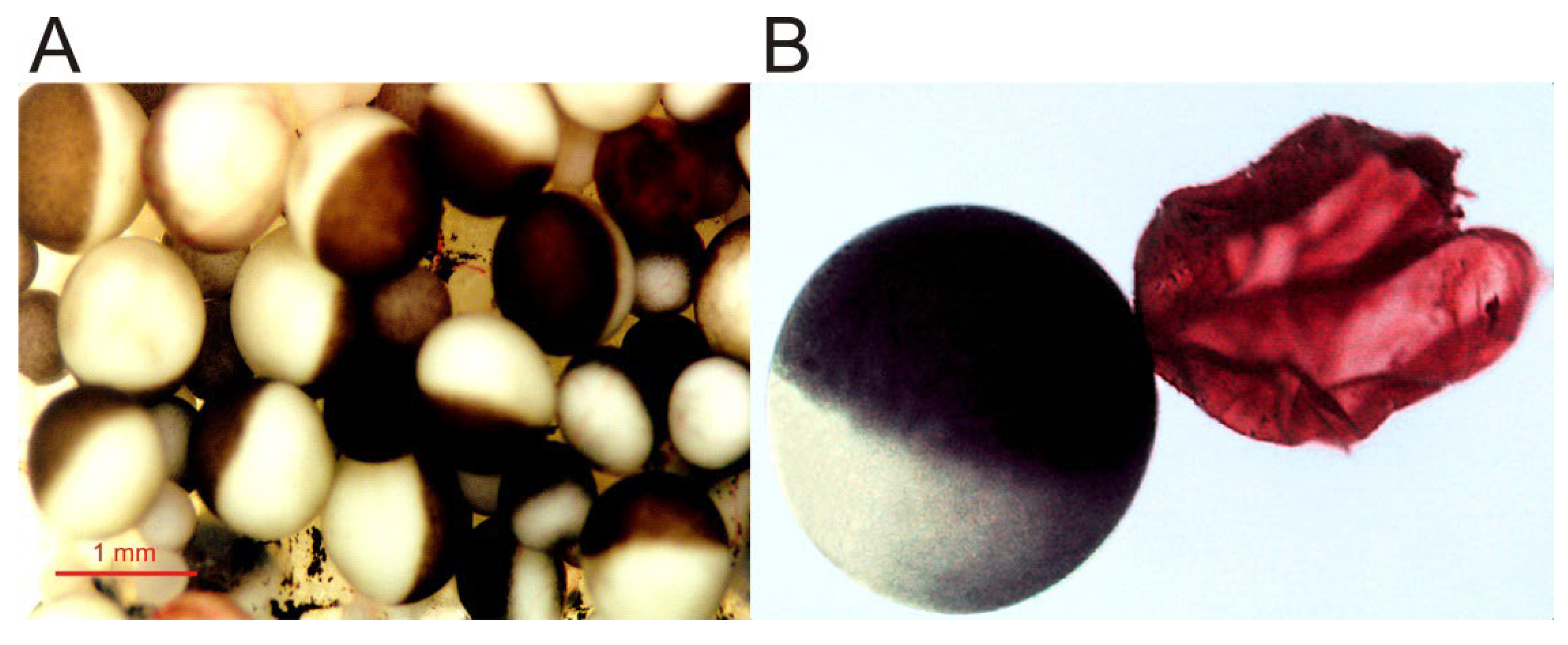
- Large size, up to 1.3 mm, and roughly spherical shape [1], which facilitates electrophysiological recordings, even allowing the introduction of additional micropipettes for microinjecting compounds [2]. Furthermore, its size enables simultaneous biochemical and optical techniques with quite high spatio-temporal discrimination [19].
- (c)
- (d)
- These cells are easy to manage and cheap to maintain. Moreover, these cells can be kept alive in an inorganic buffer for long periods (up to several weeks) after their isolation from the ovary, without the need of a specific sterile serum or medium [2,14,16]. Nevertheless, oocytes progressively uncouple from the surrounding follicular cells after their separation from the ovary, losing responsiveness to certain hormones and neurotransmitters.
- (e)
- (f)
- The oocyte plasma membrane can be quite easily isolated either manually ([30,31]; see Figure 1) or by simple biochemical procedures [32]. The isolation of plasma membranes enables one to determine the presence of the microtransplanted protein at the oocyte plasma membrane [31,33] and allows studies concerning both the targeting of membrane proteins and the quantitation of the ratios between functional and the total number of ion channels/receptors incorporated into the cell membrane [12,33,34].
- (g)
- Although the membrane of Xenopus oocytes might present certain ion channels and receptors, the expression of endogenous proteins from their own mRNA is low (about 5% of their stored mRNA, [13]). Thus the oocyte plasma membrane usually lacks significant levels of neuronal voltage-dependent Na+ and K+ channels and many neuroreceptors and transporters [10].
- (h)
- The oocyte has powerful intracellular signaling cascades, mostly involving InsP3 synthesis and Ca2+ release from intracellular stores after phospholipase-C activation. Thus, heterologous expression of certain metabotropic receptors can be simply monitored by recording Ca2+-dependent Cl− currents, since the Ca2+-dependent Cl− channel, anoctamin 1 (TMEM16A), is highly expressed in the oocyte membrane [2,27,35,36].
- (i)
- (a)
- In the ovary, oocytes are found as follicles, constituted by several cellular and acellular layers surrounding the oocyte, from inner to outer: (i) the oocyte; (ii) a fibrous vitelline membrane; (iii) a monolayer of follicular cells, which is coupled to microvilli of the oocyte membrane by gap junctions [37]; (iv) a theca, containing mainly collagen, fibrocytes and small blood vessels; and (v) a layer of epithelial cells. The layers surrounding the oocyte might constitute a handicap to study certain transplanted proteins since follicular cells express ion channels and receptors of their own and are electrically coupled to the oocyte. However, the layers surrounding the oocyte can be removed either manually or by enzymatic means.
- (b)
- There is a certain variability in the expression efficiency of oocytes, which has been related to seasonal differences [27]. Actually, some laboratories stop working with oocytes during summer because of their low quality. Furthermore, noise and other vibrations should be restricted around aquarium facilities since Xenopus are quite sensitive to them.
- (c)
- As previously mentioned, oocytes randomly express certain ion channels and receptors, which might be confused with those heterologously expressed [13]. Thus, it becomes necessary to keep in mind the channels and receptors that can be endogenously expressed by oocytes (excellent reviews regarding this issue are provided in [2,8,9,10,24,25]), though some of them can be pharmacologically blocked. Nevertheless, other cell models commonly used for the heterologous expression of ion channels and transporters such as human embryonic kidney cells (HEK) also have endogenous ion channels [38].
- (d)
- The large size of the oocyte together with the presence of several surrounding layers (cellular and acellular) constitute a limitation to obtaining fast ligand-applications, particularly when using large molecules [39], and consequently, to record fast-kinetics currents (it hinders the resolution of ligand-elicited currents at resolutions below hundred milliseconds; [40]). Moreover, when follicular cells remain attached to the oocyte, certain measured pharmacological values (i.e., half maximal effective concentration, EC50, or half maximal inhibitory concentration, IC50) might be inaccurate [39]. In addition, several factors such as the oocyte large size, the presence of numerous microvilli at its plasma membrane, and its electrical coupling to follicular cells contribute to eliciting extremely large capacitive artifacts, which prevent resolving fast voltage-dependent currents below a few milliseconds [14].
3. Transplant of Fully Processed Membrane Proteins to the Xenopus Oocyte
3.1. Characteristics of Protein Microtransplantation to the Oocyte Plasma Membrane
3.1.1. Proteoliposome-Plasma Membrane Fusion Does Not Require an Intracellular Ca2+ Increase
3.1.2. Microtransplanted Proteins Are Incorporated in Patches
3.1.3. Proteoliposomes of Different Lipid Composition Allow Functional nAChR Microtransplantation
3.2. Advantages and Limitations of Transplanting Purified Proteins Reconstituted in Artificial Lipid Bilayers
- (a)
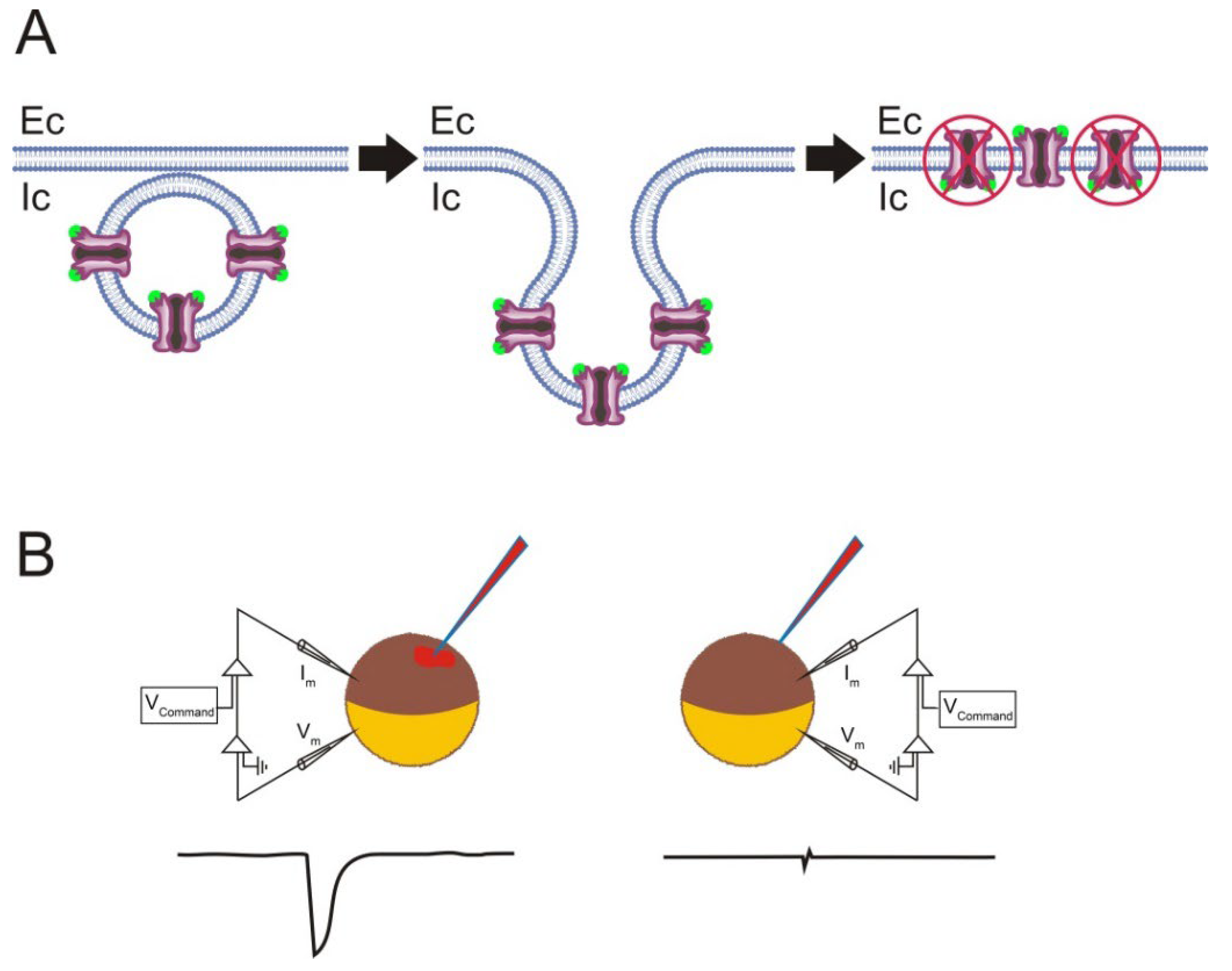
- Incorrect orientation of the incorporated proteins in the oocyte plasma membrane. Although the microtransplantation of foreign membrane proteins allows several million nAChRs to be functionally incorporated, the overall efficiency of this approach is, at least for certain proteins, rather low, considering the amount of injected protein [33,67]. Noticeably, membrane proteins of different sizes and shapes might generate proteoliposomes with different protein orientations during the process of detergent dialysis [97]. Therefore, it could be that some of the microtransplanted nAChRs were incorporated into the oocyte membrane in the “wrong” orientation, hence being functionally silent (see Figure 5). It is possible to determine the orientation of the microtransplanted nAChRs in oocytes by combining electrophysiological and binding techniques. These experiments showed the following. (i) Roughly 90% of the microtransplanted nAChRs were incorporated into the oocyte membrane with the binding domain facing up to the cytoplasm [33]. This outside-in orientation of nAChRs in the oocyte membrane could account for the low efficiency of functional nAChR microtransplantation, since nAChRs incorporated with the “wrong” orientation lost their functional activity (Figure 5; [67]). Interestingly, most nAChRs in proteoliposomes are oriented outside-out [91], but fusion of proteoliposomes with the cell membrane commonly results in an inverted protein orientation (see Figure 5; [98]). (ii) The proportion of the incorporated versus injected nAChRs, determined at a fixed time, reached up to 3%, indicating that the fusion of proteoliposomes with the cell membrane is quite an efficient process [33]. Moreover, other membrane proteins such as Torpedo Cl− channels (ClC0) were functionally incorporated in the oocyte plasma membrane much more efficiently than nAChRs [33]. Remarkably, a protein change in orientation within the lipid bilayer (protein-flipping) can be triggered, at least in certain proteins, by modifying the lipid composition of the bilayer, both in vitro and in vivo [99].
- (b)
- Presence of multilamellar proteoliposomes. Proteoliposomes bearing purified membrane proteins are commonly obtained by detergent removal dialysis [67,69]. Many of the proteoliposomes harvested under these experimental conditions are multilamellar, particularly when large proteins such as the nAChR are incorporated into liposomes. This constitutes a handicap since the presence of multilamellar proteoliposomes might affect both the protein function and its incorporation to the oocyte plasma membrane. To diminish this handicap, the harvested multilamellar proteoliposomes should be broken into smaller unilamellar vesicles, prior to injecting the sample into the oocytes. This can be attained by sonication of the sample in a water bath until the suspension becomes homogenized and less turbid [67,100,101]. Alternatively, small unilamellar proteoliposomes of homogenous sizes can be obtained from multilamellar vesicles by using a mini-extruder [102].
- (c)
- Microtransplanted proteins do not always remain surrounded by the original proteoliposome lipids. As above-mentioned, some lipid exchange might take place around the protein, as evidenced by the functional recovery of nAChR reconstituted in PC when microtransplanted to the oocyte membrane.
4. Microtransplanting nAChRs to the Xenopus Oocyte Membrane as a Useful Approach to Study Lipid–Protein Interactions
5. Use of Microtransplanted Proteins to Study Functional and Pharmacological Properties of Ion Channels and Receptors
6. Conclusions and Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| Ach | Acetylcholine; |
| AMPA | α-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid; |
| BAPTA-AM | 1,2-bis (2-aminophenoxy)ethane N,N,N′,N′-tetraacetic acid acetoxymethyl ester |
| ClC0 | Chloride channel type 0 (Torpedo) |
| Chol | Cholesterol |
| Chol/PL | Cholesterol to phospholipid molar ratio |
| δ | Fraction of the voltage field sensed by a drug in the membrane |
| DDT | Dichlorodiphenyltrichloroethane |
| EGTA | Ethyleneglycol-bis (β-aminoethylether)-N,N,N′,N′-tetraacetic acid |
| Ec | Extracellular side |
| EC50 | Half maximal effective concentration |
| GABAAR | Gamma-aminobutyric acid receptor type A |
| HEK (HEK 293) | Human embryonic kidney 293 cells |
| IACh | ACh-elicited current |
| IC50 | Half maximal inhibitory concentration |
| InsP3 | Inositol 1,4,5-trisphosphate |
| Ic | Intracellular side |
| [Ca2+]i | Intracellular Ca2+ concentration |
| KCC1 | K+–Cl− cotransporter type 1 |
| nAChR | Nicotinic acetylcholine receptor |
| NMDA | N-methyl-D-aspartate |
| PA | Phosphatidic acid |
| PC | Phosphatidylcholine |
| KcsA | A potassium channel from the Streptomyces lividans |
| R-Aso | Proteoliposome of asolectin bearing nAChRs |
| R-PA+PC+Chol | Proteoliposome of PA, PC and Chol bearing nAChRs |
| R-PC+Chol | Proteoliposome of PC and Chol bearing nAChRs |
| NKCC2 | Na+–K+–Cl− cotransporter type 2 |
| TMEM16A | Ca2+ activated Cl− channel, anoctamin1 |
References
- Dumont, J.N. Oogenesis in Xenopus laevis (Daudin). I. Stages of oocyte development in laboratory maintained animals. J. Morphol. 1972, 136, 153–179. [Google Scholar] [CrossRef] [PubMed]
- Miledi, R.; Parker, I.; Sumikawa, K. Transplanting receptors from brains into oocytes. In FIDIA Research Foundation Neuroscience Award Lectures; Raven Press, Ltd.: New York, NY, USA, 1989; Volume 3, pp. 57–90. [Google Scholar]
- Gurdon, J.B. Introductory comments: Xenopus as a laboratory animal. In The Biology of Xenopus; Tinsley, R.C., Kobel, H.R., Eds.; Clarendon Press: Oxford, UK, 1996; pp. 3–6. [Google Scholar]
- Mowry, K.L. Using the Xenopus Oocyte Toolbox. Cold Spring Harb. Protoc. 2020, 4, 095844. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Robertis, E.M.; Gurdon, J.B. A Brief History of Xenopus in Biology. Cold Spring Harb. Protoc. 2021, 12, 107615. [Google Scholar] [CrossRef] [PubMed]
- Kusano, K.; Miledi, R.; Stinnakre, J. Acetylcholine receptors in the oocyte membrane. Nature 1977, 270, 739–741. [Google Scholar] [CrossRef]
- Kusano, K.; Miledi, R.; Stinnakre, J. Cholinergic and catecholaminergic receptors in the Xenopus oocyte membrane. J. Physiol. 1982, 328, 143–170. [Google Scholar] [CrossRef]
- Dascal, N. The use of Xenopus oocytes for the study of ion channels. Crit. Rev. Biochem. Mol. Biol. 1987, 22, 317–387. [Google Scholar] [CrossRef]
- Sobczak, K.; Bangel-Ruland, N.; Leier, G.; Weber, W.M. Endogenous transport systems in the Xenopus laevis oocyte plasma membrane. Methods 2010, 51, 183–189. [Google Scholar] [CrossRef]
- Martinez-Torres, A.; Pereida-Jaramillo, E. The use of Xenopus oocytes to study the biophysics and pharmacological properties of receptors and channels. In Xenopus: From Basic Biology to Disease Models in the Genomic Era; Fainsod, A., Moody, S.A., Eds.; CRC Press: Boca Raton, FL, USA, 2022; pp. 143–151. [Google Scholar] [CrossRef]
- Weber, W.M. Ion currents of Xenopus laevis oocytes: State of the art. Biochim. Biophys. Acta 1999, 1421, 213–233. [Google Scholar] [CrossRef] [Green Version]
- Eusebi, F.; Palma, E.; Amici, M.; Miledi, R. Microtransplantation of ligand-gated receptor-channels from fresh or frozen nervous tissue into Xenopus oocytes: A potent tool for expanding functional information. Prog. Neurobiol. 2009, 88, 32–40. [Google Scholar] [CrossRef]
- Zeng, S.L.; Sudlow, L.C.; Berezin, M.Y. Using Xenopus oocytes in neurological disease drug discovery. Expert Opin. Drug Discov. 2020, 15, 39–52. [Google Scholar] [CrossRef]
- Stühmer, W.; Parekh, A.B. Electrophysiological recordings from Xenopus oocytes. In Single-Channel Recording; Sakmann, B., Neher, E., Eds.; Springer: Boston, MA, USA, 1995; pp. 341–356. [Google Scholar] [CrossRef]
- Palma, E.; Trettel, F.; Fucile, S.; Renzi, M.; Miledi, R.; Eusebi, F. Microtransplantation of membranes from cultured cells to Xenopus oocytes: A method to study neurotransmitter receptors embedded in native lipids. Proc. Natl. Acad. Sci. USA 2003, 100, 2896–2900. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kvist, T.; Hansen, K.B.; Bräuner-Osborne, H. The use of Xenopus oocytes in drug screening. Expert Opin. Drug Discov. 2011, 6, 141–153. [Google Scholar] [CrossRef] [PubMed]
- Gurdon, J.B.; Wickens, M.P. The use of Xenopus oocytes for the expression of cloned genes. Methods Enzym. 1983, 101, 370–386. [Google Scholar] [CrossRef]
- Krogh, A. The progress of Physiology. Science 1929, 70, 200–204. [Google Scholar] [CrossRef]
- Demuro, A.; Parker, I. Optical single-channel recording: Imaging Ca2+ flux through individual N-type voltage-gated channels expressed in Xenopus oocytes. Cell Calcium 2003, 34, 499–509. [Google Scholar] [CrossRef]
- Gurdon, J.B.; Lane, C.D.; Woodland, H.R.; Marbaix, G. Use of frog eggs and oocytes for the study of messenger RNA and its translation in living cells. Nature 1971, 233, 177–182. [Google Scholar] [CrossRef]
- Barnard, E.A.; Miledi, R.; Sumikawa, K. Translation of exogenous messenger RNA coding for nicotinic acetylcholine receptors produces functional receptors in Xenopus oocytes. Proc. R. Soc. Lond. Ser. B. Biol. Sci. 1982, 215, 241–246. [Google Scholar] [CrossRef]
- Miledi, R.; Parker, I.; Sumikawa, K. Properties of acetylcholine receptors translated by cat muscle mRNA in Xenopus oocytes. EMBO J. 1982, 1, 1307–1312. [Google Scholar] [CrossRef]
- Noda, M.; Ikeda, T.; Suzuki, H.; Takeshima, H.; Takahashi, T.; Kuno, M.; Numa, S. Expression of functional sodium channels from cloned cDNA. Nature 1986, 322, 826–828. [Google Scholar] [CrossRef]
- Soreq, H.; Seidman, S. Xenopus oocyte microinjection: From gene to protein. Methods Enzym. 1992, 207, 225–265. [Google Scholar] [CrossRef]
- Miller, A.J.; Zhou, J.J. Xenopus oocytes as an expression system for plant transporters. Biochim. Biophys. Acta 2000, 1465, 343–358. [Google Scholar] [CrossRef] [Green Version]
- Miledi, R.; Palma, E.; Eusebi, F. Microtransplantation of neurotransmitter receptors from cells to Xenopus oocyte membranes: New procedure for ion channel studies. Methods Mol. Biol. 2006, 322, 347–355. [Google Scholar] [CrossRef]
- Terhag, J.; Cavara, N.A.; Hollmann, M. Cave Canalem: How endogenous ion channels may interfere with heterologous expression in Xenopus oocytes. Methods 2010, 51, 66–74. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.B.; Logee, K.A.; Verkman, A.S. Expression of mRNA coding for kidney and red cell water channels in Xenopus oocytes. J. Biol. Chem. 1990, 265, 15375–15378. [Google Scholar] [CrossRef]
- Preston, G.M.; Carroll, T.P.; Guggino, W.B.; Agre, P. Appearance of water channels in Xenopus oocytes expressing red cell CHIP28 protein. Science 1992, 256, 385–387. [Google Scholar] [CrossRef] [Green Version]
- Sadler, S.E.; Maller, J.L. Progesterone inhibits adenylate cyclase in Xenopus oocytes. Action on the guanine nucleotide regulatory protein. J. Biol. Chem. 1981, 256, 6368–6373. [Google Scholar] [CrossRef]
- Aleu, J.; Ivorra, I.; Lejarreta, M.; González-Ros, J.M.; Morales, A.; Ferragut, J.A. Functional incorporation of P-glycoprotein into Xenopus oocyte plasma membrane fails to elicit a swelling-evoked conductance. Biochem. Biophys. Res. Commun. 1997, 237, 407–412. [Google Scholar] [CrossRef] [Green Version]
- Leduc-Nadeau, A.; Lahjouji, K.; Bissonnette, P.; Lapointe, J.Y.; Bichet, D.G. Elaboration of a novel technique for purification of plasma membranes from Xenopus laevis oocytes. Am. J. Physiol. Cell Physiol. 2007, 292, C1132–C1136. [Google Scholar] [CrossRef]
- Ivorra, I.; Fernández, A.; Gal, B.; Aleu, J.; González-Ros, J.M.; Ferragut, J.A.; Morales, A. Protein orientation affects the efficiency of functional protein transplantation into the Xenopus oocyte membrane. J. Membr. Biol. 2002, 185, 117–127. [Google Scholar] [CrossRef]
- Murenzi, E.; Toltin, A.C.; Symington, S.B.; Morgan, M.M.; Clark, J.M. Evaluation of microtransplantation of rat brain neurolemma into Xenopus laevis oocytes as a technique to study the effect of neurotoxicants on endogenous voltage-sensitive ion channels. Neurotoxicology 2017, 60, 260–273. [Google Scholar] [CrossRef]
- Miledi, R. A calcium-dependent transient outward current in Xenopus laevis oocytes. Proc. R. Soc. Lond. B. Biol. Sci. 1982, 215, 491–497. [Google Scholar] [CrossRef] [PubMed]
- Barish, M.E. A transient calcium-dependent chloride current in the immature Xenopus oocyte. J. Physiol. 1983, 342, 309–325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Browne, C.L.; Wiley, H.S.; Dumont, J.N. Oocyte-follicle cell gap junctions in Xenopus laevis and the effects of gonadotropin on their permeability. Science 1979, 203, 182–183. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Yuan, H.; Yao, X.; Chen, S. Endogenous ion channels expressed in human embryonic kidney (HEK-293) cells. Pflugers Arch.-Eur. J. Physiol. 2022, 474, 665–680. [Google Scholar] [CrossRef]
- Madeja, M.; Musshoff, U.; Speckmann, E.J. Follicular tissues reduce drug effects on ion channels in oocytes of Xenopus laevis. Eur. J. Neurosci. 1997, 9, 599–604. [Google Scholar] [CrossRef]
- Gielen, M.; Corringer, P.-J. The dual-gate model for pentameric ligand-gated ion channels activation and desensitization. J. Physiol. 2018, 596, 1873–1902. [Google Scholar] [CrossRef] [Green Version]
- Buller, A.L.; White, M.M. Altered patterns of N-linked glycosylation of the Torpedo acetylcholine receptor expressed in Xenopus oocytes. J. Membr. Biol. 1990, 115, 179–189. [Google Scholar] [CrossRef]
- Sivilotti, L.G.; McNeil, D.K.; Lewis, T.M.; Nassar, M.A.; Schoepfer, R.; Colquhoun, D. Recombinant nicotinic receptors, expressed in Xenopus oocytes, do not resemble native rat sympathetic ganglion receptors in single-channel behaviour. J. Physiol. 1997, 500, 123–138. [Google Scholar] [CrossRef]
- Truong, A.; Xing, X.; Forsayeth, J.R.; Dwoskin, L.P.; Crooks, P.A.; Cohen, B.N. Pharmacological differences between immunoisolated native brain and heterologously expressed rat alpha4beta2 nicotinic receptors. Brain Res. Mol. Brain Res. 2001, 96, 68–76. [Google Scholar] [CrossRef]
- Jospin, M.; Bonneau, B.; Lainé, V.; Bessereau, J.L. An extracellular scaffolding complex confers unusual rectification upon an ionotropic acetylcholine receptor in C. elegans. Proc. Natl. Acad. Sci. USA 2022, 119, e2113545119. [Google Scholar] [CrossRef]
- Opekarová, M.; Tanner, W. Specific lipid requirements of membrane proteins-a putative bottleneck in heterologous expression. Biochim. Biophys. Acta 2003, 1610, 11–22. [Google Scholar] [CrossRef] [Green Version]
- Mesoy, S.M.; Bridgland-Taylor, M.; Lummis, S.C.R. Mutations of the nACh Receptor M4 Helix Reveal Different Phenotypes in Different Expression Systems: Could Lipids be Responsible? Front. Physiol. 2022, 13, 850782. [Google Scholar] [CrossRef] [PubMed]
- Marsal, J.; Tigyi, G.; Miledi, R. Incorporation of acetylcholine receptors and Cl- channels in Xenopus oocytes injected with Torpedo electroplaque membranes. Proc. Natl. Acad. Sci. USA 1995, 92, 5224–5228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bernareggi, A.; Reyes-Ruiz, J.M.; Lorenzon, P.; Ruzzier, F.; Miledi, R. Microtransplantation of acetylcholine receptors from normal or denervated rat skeletal muscles to frog oocytes. J. Physiol. 2011, 589, 1133–1142. [Google Scholar] [CrossRef]
- Palma, E.; Inghilleri, M.; Conti, L.; Deflorio, C.; Frasca, V.; Manteca, A.; Pichiorri, F.; Roseti, C.; Torchia, G.; Limatola, C.; et al. Physiological characterization of human muscle acetylcholine receptors from ALS patients. Proc. Natl. Acad. Sci. USA 2011, 108, 20184–20188. [Google Scholar] [CrossRef] [Green Version]
- Olivera-Bravo, S.; Ivorra, I.; Morales, A. Differential effects of quaternary ammonium anticholinesterases on microtransplanted neuroreceptors: Selective modulation of nicotinic receptor function. J. Mol. Neurosci. 2010, 40, 251–252. [Google Scholar] [CrossRef]
- Sanna, E.; Motzo, C.; Usala, M.; Pau, D.; Cagetti, E.; Biggio, G. Functional changes in rat nigral GABA(A) receptors induced by degeneration of the striatonigral GABAergic pathway: An electrophysiological study of receptors incorporated into Xenopus oocytes. J. Neurochem. 1998, 70, 2539–2544. [Google Scholar] [CrossRef]
- Miledi, R.; Eusebi, F.; Martínez-Torres, A.; Palma, E.; Trettel, F. Expression of functional neurotransmitter receptors in Xenopus oocytes after injection of human brain membranes. Proc. Natl. Acad. Sci. USA 2002, 99, 13238–13242. [Google Scholar] [CrossRef]
- Palma, E.; Torchia, G.; Limatola, C.; Trettel, F.; Arcella, A.; Cantore, G.; Di Gennaro, G.; Manfredi, M.; Esposito, V.; Quarato, P.P.; et al. BDNF modulates GABAA receptors microtransplanted from the human epileptic brain to Xenopus oocytes. Proc. Natl. Acad. Sci. USA 2005, 102, 1667–1672. [Google Scholar] [CrossRef] [Green Version]
- Palma, E.; Spinelli, G.; Torchia, G.; Martínez-Torres, A.; Ragozzino, D.; Miledi, R.; Eusebi, F. Abnormal GABAA receptors from the human epileptic hippocampal subiculum microtransplanted to Xenopus oocytes. Proc. Natl. Acad. Sci. USA. 2005, 102, 2514–2518. [Google Scholar] [CrossRef] [Green Version]
- Palma, E.; Amici, M.; Sobrero, F.; Spinelli, G.; Di Angelantonio, S.; Ragozzino, D.; Mascia, A.; Scoppetta, C.; Esposito, V.; Miledi, R.; et al. Anomalous levels of Cl- transporters in the hippocampal subiculum from temporal lobe epilepsy patients make GABA excitatory. Proc. Natl. Acad. Sci. USA 2006, 103, 8465–8468. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palma, E.; Roseti, C.; Maiolino, F.; Fucile, S.; Martinello, K.; Mazzuferi, M.; Aronica, E.; Manfredi, M.; Esposito, V.; Cantore, G.; et al. GABA(A)-current rundown of temporal lobe epilepsy is associated with repetitive activation of GABA(A) “phasic” receptors. Proc. Natl. Acad. Sci. USA 2007, 104, 20944–20948. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palma, E.; Ragozzino, D.; Di Angelantonio, S.; Mascia, A.; Maiolino, F.; Manfredi, M.; Cantore, G.; Esposito, V.; Di Gennaro, G.; Quarato, P.; et al. The antiepileptic drug levetiracetam stabilizes the human epileptic GABAA receptors upon repetitive activation. Epilepsia 2007, 48, 1842–1849. [Google Scholar] [CrossRef] [PubMed]
- Limon, A.; Reyes-Ruiz, J.M.; Miledi, R. Microtransplantation of neurotransmitter receptors from postmortem autistic brains to Xenopus oocytes. Proc. Natl. Acad. Sci. USA 2008, 105, 10973–10977. [Google Scholar] [CrossRef] [Green Version]
- Alberola-Die, A.; Fernández-Ballester, G.; González-Ros, J.M.; Ivorra, I.; Morales, A. Muscle-type nicotinic receptor blockade by diethylamine, the hydrophilic moiety of lidocaine. Front. Mol. Neurosci. 2016, 9, 12. [Google Scholar] [CrossRef] [Green Version]
- Alberola-Die, A.; Fernández-Ballester, G.; González-Ros, J.M.; Ivorra, I.; Morales, A. Muscle-type nicotinic receptor modulation by 2,6-dimethylaniline, a molecule resembling the hydrophobic moiety of lidocaine. Front. Mol. Neurosci. 2016, 9, 127. [Google Scholar] [CrossRef] [Green Version]
- Mazzo, F.; Zwart, R.; Serratto, G.M.; Gardinier, K.M.; Porter, W.; Reel, J.; Maraula, G.; Sher, E. Reconstitution of synaptic ion channels from rodent and human brain in Xenopus oocytes: A biochemical and electrophysiological characterization. J. Neurochem. 2016, 138, 384–396. [Google Scholar] [CrossRef] [Green Version]
- Burgos, J.S.; Aleu, J.; Barat, A.; Solsona, C.; Marsal, J.; Ramírez, G. Kainate-triggered currents in Xenopus oocytes injected with chick retinal membrane fragments: Effect of guanine nucleotides. Investig. Ophthalmol. Vis. Sci. 2003, 44, 3124–3129. [Google Scholar] [CrossRef]
- Bernareggi, A.; Dueñas, Z.; Reyes-Ruiz, J.M.; Ruzzier, F.; Miledi, R. Properties of glutamate receptors of Alzheimer’s disease brain transplanted to frog oocytes. Proc. Natl. Acad. Sci. USA 2007, 104, 2956–2960. [Google Scholar] [CrossRef] [Green Version]
- Sandoval, M.; Sandoval, R.; Thomas, U.; Spilker, C.; Smalla, K.H.; Falcon, R.; Marengo, J.J.; Calderón, R.; Saavedra, V.; Heumann, R.; et al. Antagonistic effects of TrkB and p75(NTR) on NMDA receptor currents in post-synaptic densities transplanted into Xenopus oocytes. J. Neurochem. 2007, 101, 1672–1684. [Google Scholar] [CrossRef] [PubMed]
- Miledi, R.; Dueñas, Z.; Martinez-Torres, A.; Kawas, C.H.; Eusebi, F. Microtransplantation of functional receptors and channels from the Alzheimer’s brain to frog oocytes. Proc. Natl. Acad. Sci. USA 2004, 101, 1760–1763. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ivorra, I.; Henriquez, M.; Lax, P.; Riquelme, G.; Morales, A. Functional transplantation of chloride channels from the human syncytiotrophoblast microvillous membrane to Xenopus oocytes. Pflug. Arch.-Eur. J. Physiol. 2002, 444, 685–691. [Google Scholar] [CrossRef]
- Morales, A.; Aleu, J.; Ivorra, I.; Ferragut, J.A.; González-Ros, J.M.; Miledi, R. Incorporation of reconstituted acetylcholine receptors from Torpedo into the Xenopus oocyte membrane. Proc. Natl. Acad. Sci. USA 1995, 92, 8468–8472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morales, A.; de Juan, E.; Fernández-Carvajal, A.M.; Martinez-Pinna, J.; Poveda, J.A.; Encinar, J.A.; Ivorra, I.; González-Ros, J.M. Nicotinic acetylcholine receptor properties are modulated by surrounding lipids: An in vivo study. J. Mol. Neurosci. 2006, 30, 5–6. [Google Scholar] [CrossRef] [Green Version]
- Le Cahérec, F.; Bron, P.; Verbavatz, J.M.; Garret, A.; Morel, G.; Cavalier, A.; Bonnec, G.; Thomas, D.; Gouranton, J.; Hubert, J.F. Incorporation of proteins into (Xenopus) oocytes by proteoliposome microinjection: Functional characterization of a novel aquaporin. J. Cell Sci. 1996, 109, 1285–1295. [Google Scholar] [CrossRef]
- Thompson, M.J.; Baenziger, J.E. Ion channels as lipid sensors: From structures to mechanisms. Nat. Chem. Biol. 2020, 16, 1331–1342. [Google Scholar] [CrossRef]
- Vallés, A.S.; Barrantes, F.J. Dysregulation of neuronal nicotinic acetylcholine receptor-cholesterol crosstalk in autism spectrum disorder. Front. Mol. Neurosci. 2021, 14, 744597. [Google Scholar] [CrossRef]
- Martinac, B.; Hamill, O.P. Gramicidin A channels switch between stretch activation and stretch inactivation depending on bilayer thickness. Proc. Natl. Acad. Sci. USA 2002, 99, 4308–4312. [Google Scholar] [CrossRef]
- Lundbaek, J.A.; Birn, P.; Hansen, A.J.; Søgaard, R.; Nielsen, C.; Girshman, J.; Bruno, M.J.; Tape, S.E.; Egebjerg, J.; Greathouse, D.V.; et al. Regulation of sodium channel function by bilayer elasticity: The importance of hydrophobic coupling. Effects of Micelle-forming amphiphiles and cholesterol. J. Gen. Physiol. 2004, 123, 599–621. [Google Scholar] [CrossRef] [Green Version]
- Gal, B.; Ivorra, I.; Morales, A. Functional incorporation of exogenous proteins into the Xenopus oocyte membrane does not depend on intracellular calcium increase. Pflug. Arch.-Eur. J. Physiol. 2000, 440, 852–857. [Google Scholar] [CrossRef]
- Tigyi, G.; Dyer, D.; Matute, C.; Miledi, R. A serum factor that activates the phosphatidylinositol phosphate signaling system in Xenopus oocytes. Proc. Natl. Acad. Sci. USA 1990, 87, 1521–1525. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Durieux, M.E.; Salafranca, M.N.; Lynch, K.R.; Moorman, J.R. Lysophosphatidic acid induces a pertussis toxin-sensitive Ca(2+)-activated Cl- current in Xenopus laevis oocytes. Am. J. Physiol. 1992, 263, C896–C900. [Google Scholar] [CrossRef] [PubMed]
- Joardar, A.; Pattnaik, G.P.; Chakraborty, H. Mechanism of membrane fusion: Interplay of lipid and peptide. J. Membr. Biol. 2022, 255, 211–224. [Google Scholar] [CrossRef] [PubMed]
- Unwin, N. Protein-lipid interplay at the neuromuscular junction. Microscopy 2022, 71, i66–i71. [Google Scholar] [CrossRef]
- Unwin, N. Structure of a cholinergic cell membrane. Proc. Natl. Acad. Sci. USA 2022, 119, e2207641119. [Google Scholar] [CrossRef]
- Fong, T.M.; McNamee, M.G. Correlation between acetylcholine receptor function and structural properties of membranes. Biochemistry 1986, 25, 830–840. [Google Scholar] [CrossRef]
- Fernández-Ballester, G.; Castresana, J.; Fernández, A.M.; Arrondo, J.L.; Ferragut, J.A.; González-Ros, J.M. A role for cholesterol as a structural effector of the nicotinic acetylcholine receptor. Biochemistry 1994, 33, 4065–4071. [Google Scholar] [CrossRef]
- Fernández, A.M.; Poveda, J.A.; Encinar, J.A.; Morales, A.; González-Ros, J.M. Structural and functional modulation of ion channels by specific lipids: From model systems to cell membranes. In Protein-Lipid Interactions, 1st ed.; Mateo, C.R., Gómez, J., Villalaín, J., González-Ros, J.M., Eds.; Springer Series in Biophysics; Springer: Berlin, Germany, 2006; Volume 9, pp. 121–231. [Google Scholar] [CrossRef]
- Poveda, J.A.; Giudici, M.A.; Renart, L.M.; Morales, A.; González-Ros, J.M. Towards understanding the molecular basis of ion channel modulation by lipids: Mechanistic models and current paradigms. Biochim. Biophys. Acta Biomembr. 2017, 1859, 1507–1516. [Google Scholar] [CrossRef]
- daCosta, C.J.; Ogrel, A.A.; McCardy, E.A.; Blanton, M.P.; Baenziger, J.E. Lipid-protein interactions at the nicotinic acetylcholine receptor. A functional coupling between nicotinic receptors and phosphatidic acid-containing lipid bilayers. J. Biol. Chem. 2002, 277, 201–208. [Google Scholar] [CrossRef] [Green Version]
- Poveda, J.A.; Encinar, J.A.; Fernández, A.M.; Mateo, C.R.; Ferragut, J.A.; González-Ros, J.M. Segregation of phosphatidic acid-rich domains in reconstituted acetylcholine receptor membranes. Biochemistry 2002, 41, 12253–12262. [Google Scholar] [CrossRef]
- Caldironi, H.A.; Alonso, T.S. Lipidic characterization of full-grown amphibian oocytes and their plasma membrane-enriched fractions. Lipids 1996, 31, 651–656. [Google Scholar] [CrossRef] [PubMed]
- Stith, B.J.; Hall, J.; Ayres, P.; Waggoner, L.; Moore, J.D.; Shaw, W.A. Quantification of major classes of Xenopus phospholipids by high performance liquid chromatography with evaporative light scattering detection. J. Lipid. Res. 2000, 41, 1448–1454. [Google Scholar] [CrossRef]
- Unwin, N. Segregation of lipids near acetylcholine-receptor channels imaged by cryo-EM. IUCrJ 2017, 4, 393–399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Unwin, N. Protein-lipid architecture of a cholinergic postsynaptic membrane. IUCrJ 2020, 7, 852–859. [Google Scholar] [CrossRef]
- González-Ros, J.M.; Llanillo, M.; Paraschos, A.; Martinez-Carrión, M. Lipid environment of acetylcholine receptor from Torpedo Californica. Biochemistry 1982, 21, 3467–3474. [Google Scholar] [CrossRef]
- Criado, M.; Eibl, H.; Barrantes, F.J. Effects of lipids on acetylcholine receptor. Essential need of cholesterol for maintenance of agonist-induced state transitions in lipid vesicles. Biochemistry 1982, 21, 3622–3629. [Google Scholar] [CrossRef]
- Hamouda, A.K.; Sanghvi, M.; Sauls, D.; Machu, T.K.; Blanton, M.P. Assessing the lipid requirements of the Torpedo californica nicotinic acetylcholine receptor. Biochemistry 2006, 45, 4327–4337. [Google Scholar] [CrossRef]
- Brannigan, G.; Hénin, J.; Law, R.; Eckenhoff, R.; Klein, M.L. Embedded cholesterol in the nicotinic acetylcholine receptor. Proc. Natl. Acad. Sci. USA 2008, 105, 14418–14423. [Google Scholar] [CrossRef] [Green Version]
- Santiago, J.; Guzmán, G.R.; Rojas, L.V.; Marti, R.; Asmar-Rovira, G.A.; Santana, L.F.; McNamee, M.; Lasal-de-Dominicci, J.A. Probing the effects of membrane cholesterol in the Torpedo californica acetylcholine receptor and the novel lipid-exposed mutation alpha C418W in Xenopus oocytes. J. Biol. Chem. 2001, 276, 46523–46532. [Google Scholar] [CrossRef] [Green Version]
- Latorre, R.; Labarca, P.; Naranjo, D. Surface charge effects on ion conduction in ion channels. Methods Enzym. 1992, 207, 471–501. [Google Scholar] [CrossRef]
- Anzai, K.; Takano, C.; Tanaka, K.; Kirino, Y. Asymmetrical lipid charge changes the subconducting state of the potassium channel from sarcoplasmic reticulum. Biochem. Biophys. Res. Commun. 1994, 199, 1081–1087. [Google Scholar] [CrossRef] [PubMed]
- Wrigglesworth, J.M.; Wooster, M.S.; Elsden, J.; Danneel, H.J. Dynamics of proteoliposome formation. Intermediate states during detergent dialysis. Biochem. J. 1987, 246, 737–744. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Richards, M.J.; Hsia, C.Y.; Singh, R.R.; Haider, H.; Kumpf, J.; Kawate, T.; Daniel, S. Membrane protein mobility and orientation preserved in supported bilayers created directly from cell plasma membrane blebs. Langmuir 2016, 32, 2963–2974. [Google Scholar] [CrossRef] [PubMed]
- Vitrac, H.; MacLean, D.M.; Jayaraman, V.; Bogdanov, M.; Dowhan, W. Dynamic membrane protein topological switching upon changes in phospholipid environment. Proc. Natl. Acad. Sci. USA 2015, 112, 13874–13879. [Google Scholar] [CrossRef] [Green Version]
- Pace, H.; Simonsson Nyström, L.; Gunnarsson, A.; Eck, E.; Monson, C.; Geschwindner, S.; Snijder, A.; Höök, F. Preserved transmembrane protein mobility in polymer-supported lipid bilayers derived from cell membranes. Anal. Chem. 2015, 87, 9194–9203. [Google Scholar] [CrossRef]
- Stockbridge, R.B.; Tsai, M.F. Lipid reconstitution and recording of recombinant ion channels. Methods Enzym. 2015, 556, 385–404. [Google Scholar] [CrossRef]
- Sejwal, K.; Chami, M.; Baumgartner, P.; Kowal, J.; Müller, S.; Stahlberg, H. Proteoliposomes-a system to study membrane proteins under buffer gradients by cryo-EM. Nanotechnol. Rev. 2017, 6, 57–74. [Google Scholar] [CrossRef]
- Poveda, J.A.; Giudici, A.M.; Renart, M.L.; Millet, O.; Morales, A.; González-Ros, J.M.; Oakes, V.; Furini, S.; Domene, C. Modulation of the potassium channel KcsA by anionic phospholipids: Role of arginines at the non-annular lipid binding sites. Biochim. Biophys. Acta Biomembr. 2019, 1861, 183029. [Google Scholar] [CrossRef]
- Renart, M.L.; Giudici, A.M.; Díaz-García, C.; Molina, M.L.; Morales, A.; González-Ros, J.M.; Poveda, J.A. Modulation of function, structure and clustering of K+ channels by lipids: Lessons learnt from KcsA. Int. J. Mol. Sci. 2020, 21, 2554. [Google Scholar] [CrossRef] [Green Version]
- Olivera-Bravo, S.; Ivorra, I.; Morales, A. The acetylcholinesterase inhibitor BW284c51 is a potent blocker of Torpedo nicotinic AchRs incorporated into the Xenopus oocyte membrane. Br. J. Pharmacol. 2005, 144, 88–97. [Google Scholar] [CrossRef] [Green Version]
- Olivera-Bravo, S.; Ivorra, I.; Morales, A. Diverse inhibitory actions of quaternary ammonium cholinesterase inhibitors on Torpedo nicotinic ACh receptors transplanted to Xenopus oocytes. Br. J. Pharmacol. 2007, 151, 1280–1292. [Google Scholar] [CrossRef] [PubMed]
- Alberola-Die, A.; Martinez-Pinna, J.; González-Ros, J.M.; Ivorra, I.; Morales, A. Multiple inhibitory actions of lidocaine on Torpedo nicotinic acetylcholine receptors transplanted to Xenopus oocytes. J. Neurochem. 2011, 117, 1009–1019. [Google Scholar] [CrossRef]
- Cobo, R.; Nikolaeva, M.; Alberola-Die, A.; Fernández-Ballester, G.; González-Ros, J.M.; Ivorra, I.; Morales, A. Mechanisms underlying the strong inhibition of muscle-type nicotinic receptors by tetracaine. Front. Mol. Neurosci. 2018, 11, 193. [Google Scholar] [CrossRef] [PubMed]
- Cobo, R.; Nikolaeva-Koleva, M.; Alberola-Die, A.; Fernández-Ballester, G.; González-Ros, J.M.; Ivorra, I.; Morales, A. Mechanisms of blockade of the muscle-type nicotinic receptor by benzocaine, a permanently uncharged local anesthetic. Neuroscience 2020, 439, 62–79. [Google Scholar] [CrossRef] [PubMed]
- Alberola-Die, A.; Encinar, J.A.; Cobo, R.; Fernández-Ballester, G.; González-Ros, J.M.; Ivorra, I.; Morales, A. Peimine, an anti-inflammatory compound from Chinese herbal extracts, modulates muscle-type nicotinic receptors. Int. J. Mol. Sci. 2021, 22, 11287. [Google Scholar] [CrossRef] [PubMed]
- Woodhull, A.M. Ionic blockage of sodium channels in nerve. J. Gen. Physiol. 1973, 61, 687–708. [Google Scholar] [CrossRef] [Green Version]
- Spitzer, N.C. A Rosetta stone for analysis of human membrane protein function. Proc. Natl. Acad. Sci. USA 2008, 105, 10641–10642. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ivorra, I.; Alberola-Die, A.; Cobo, R.; González-Ros, J.M.; Morales, A. Xenopus Oocytes as a Powerful Cellular Model to Study Foreign Fully-Processed Membrane Proteins. Membranes 2022, 12, 986. https://doi.org/10.3390/membranes12100986
Ivorra I, Alberola-Die A, Cobo R, González-Ros JM, Morales A. Xenopus Oocytes as a Powerful Cellular Model to Study Foreign Fully-Processed Membrane Proteins. Membranes. 2022; 12(10):986. https://doi.org/10.3390/membranes12100986
Chicago/Turabian StyleIvorra, Isabel, Armando Alberola-Die, Raúl Cobo, José Manuel González-Ros, and Andrés Morales. 2022. "Xenopus Oocytes as a Powerful Cellular Model to Study Foreign Fully-Processed Membrane Proteins" Membranes 12, no. 10: 986. https://doi.org/10.3390/membranes12100986
APA StyleIvorra, I., Alberola-Die, A., Cobo, R., González-Ros, J. M., & Morales, A. (2022). Xenopus Oocytes as a Powerful Cellular Model to Study Foreign Fully-Processed Membrane Proteins. Membranes, 12(10), 986. https://doi.org/10.3390/membranes12100986