
Transmembrane Membrane Readers form a Novel Class of Proteins That Include Peripheral Phosphoinositide Recognition Domains and Viral Spikes

Abstract
:1. Introduction
2. Materials and Methods
2.1. Sequences and Modifications
2.2. Protein Structures
2.3. Transmembrane Helix Prediction
2.4. Coiled Coil Prediction
2.5. Membrane Docking Site Prediction
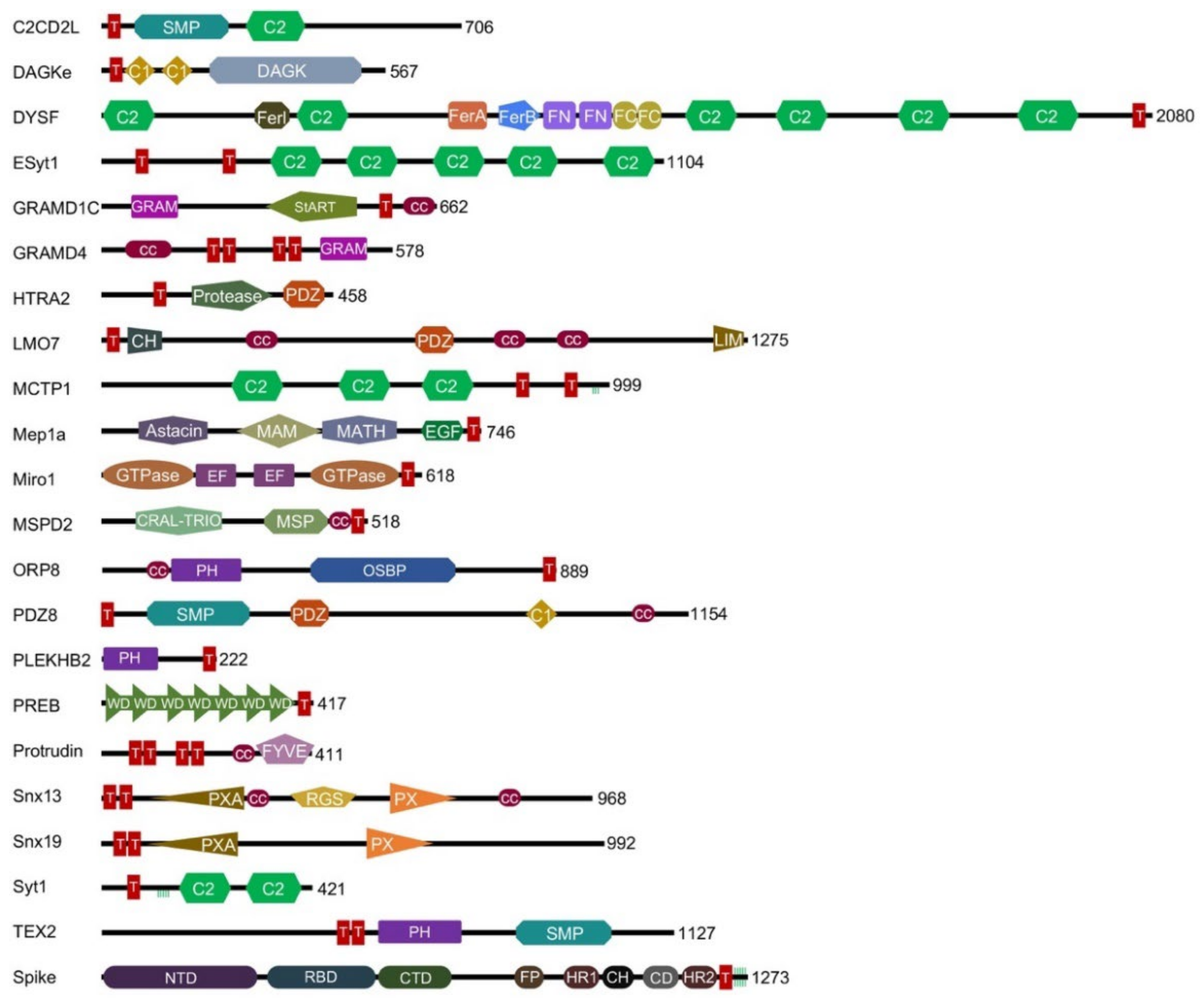
3. Results
3.1. The C2 Domains Are the Most Common TMMR Module
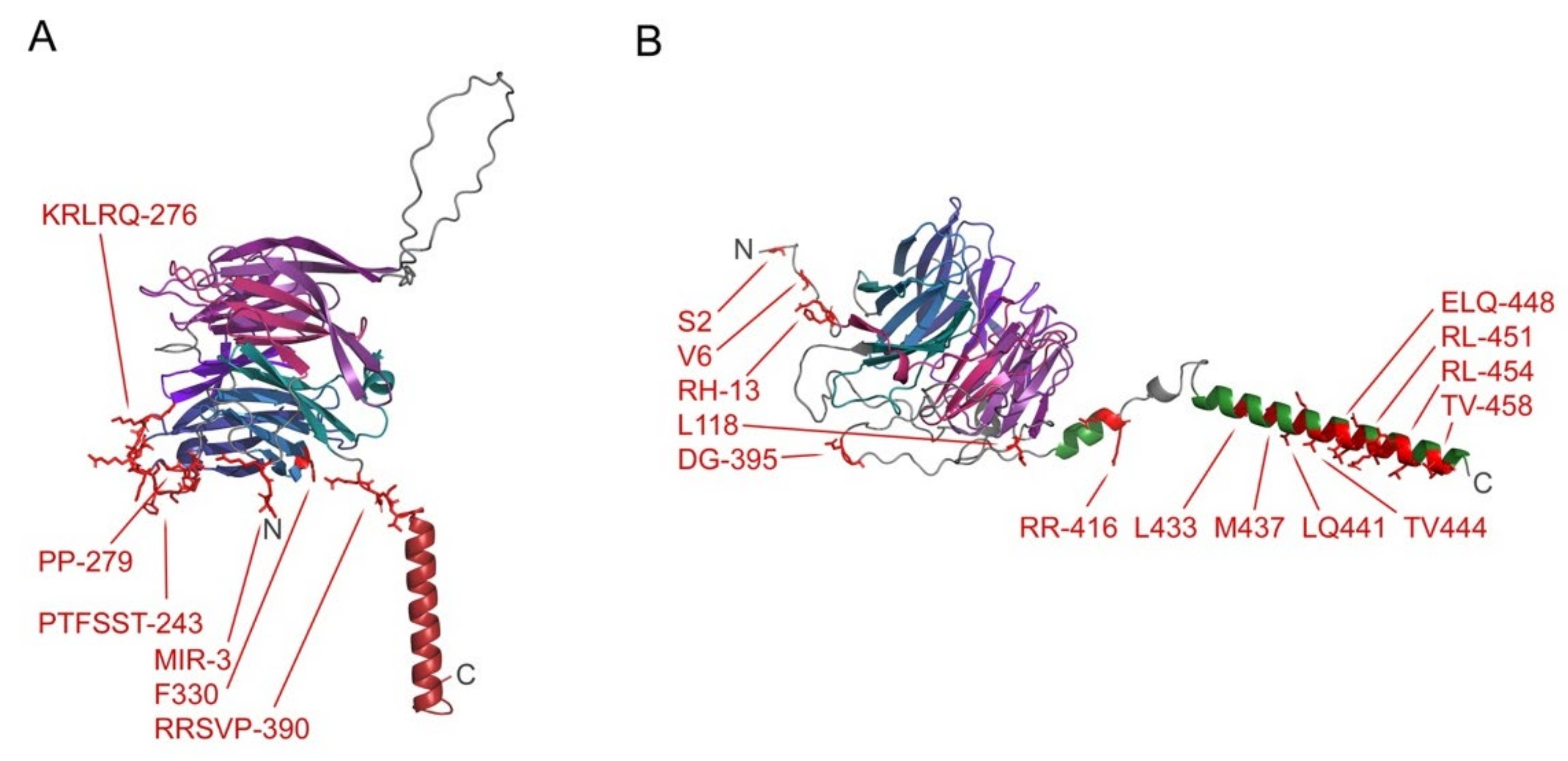
3.2. C1 Domain-Containing Lipid Kinase
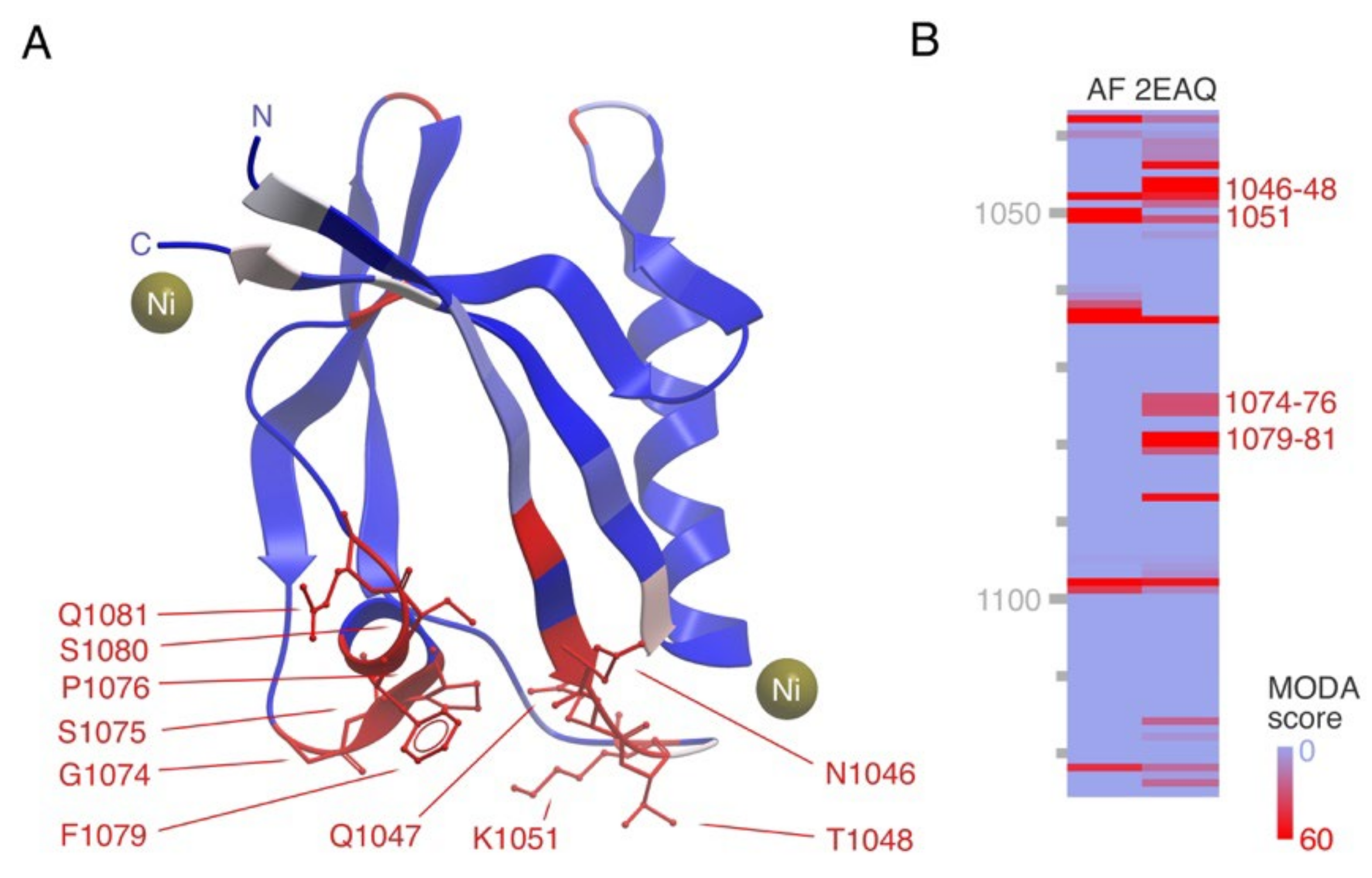
3.3. PDZ Domain-Containing Integral Membrane Proteins
3.4. PH and GRAM Domains Are Common in TMMRs
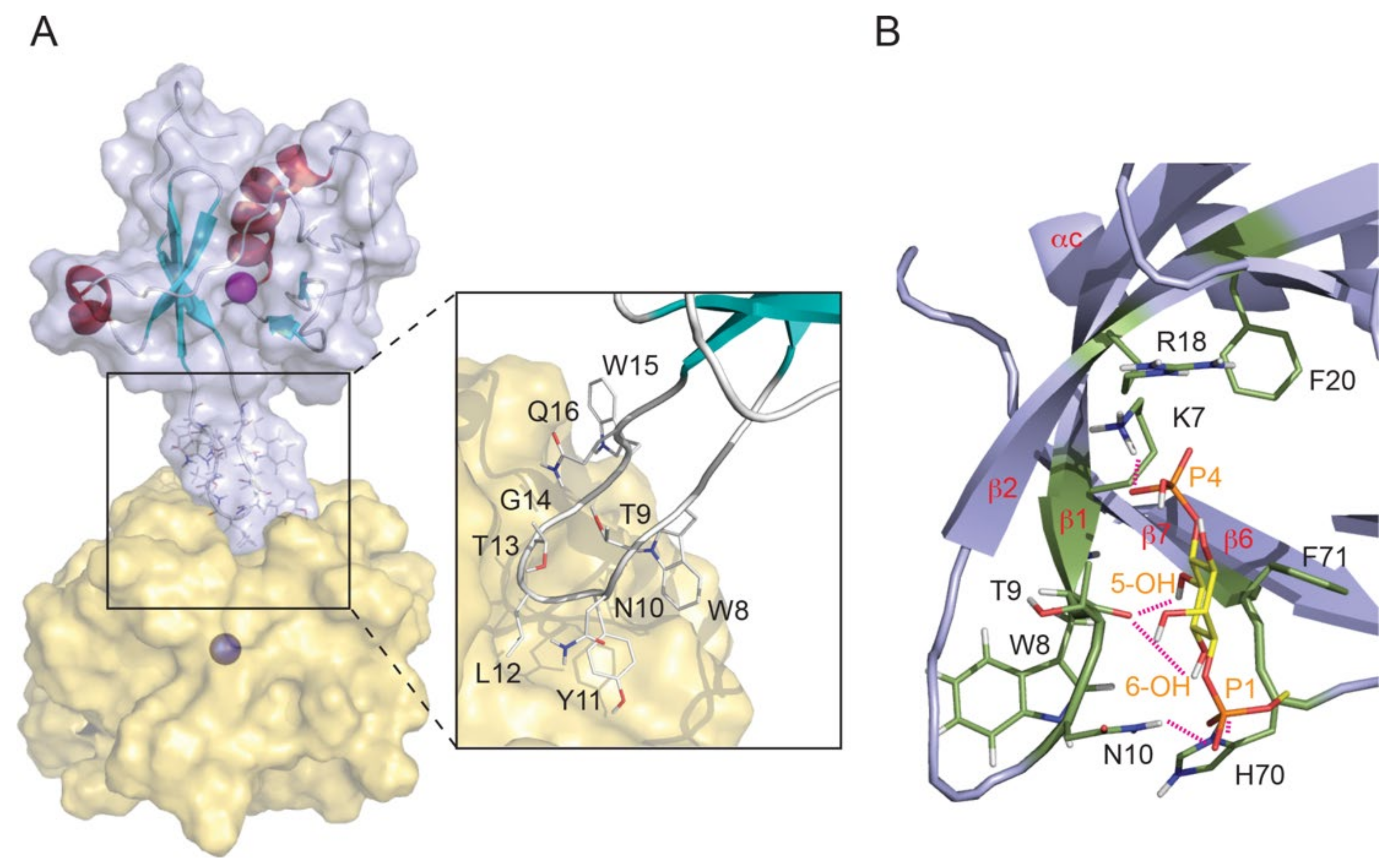
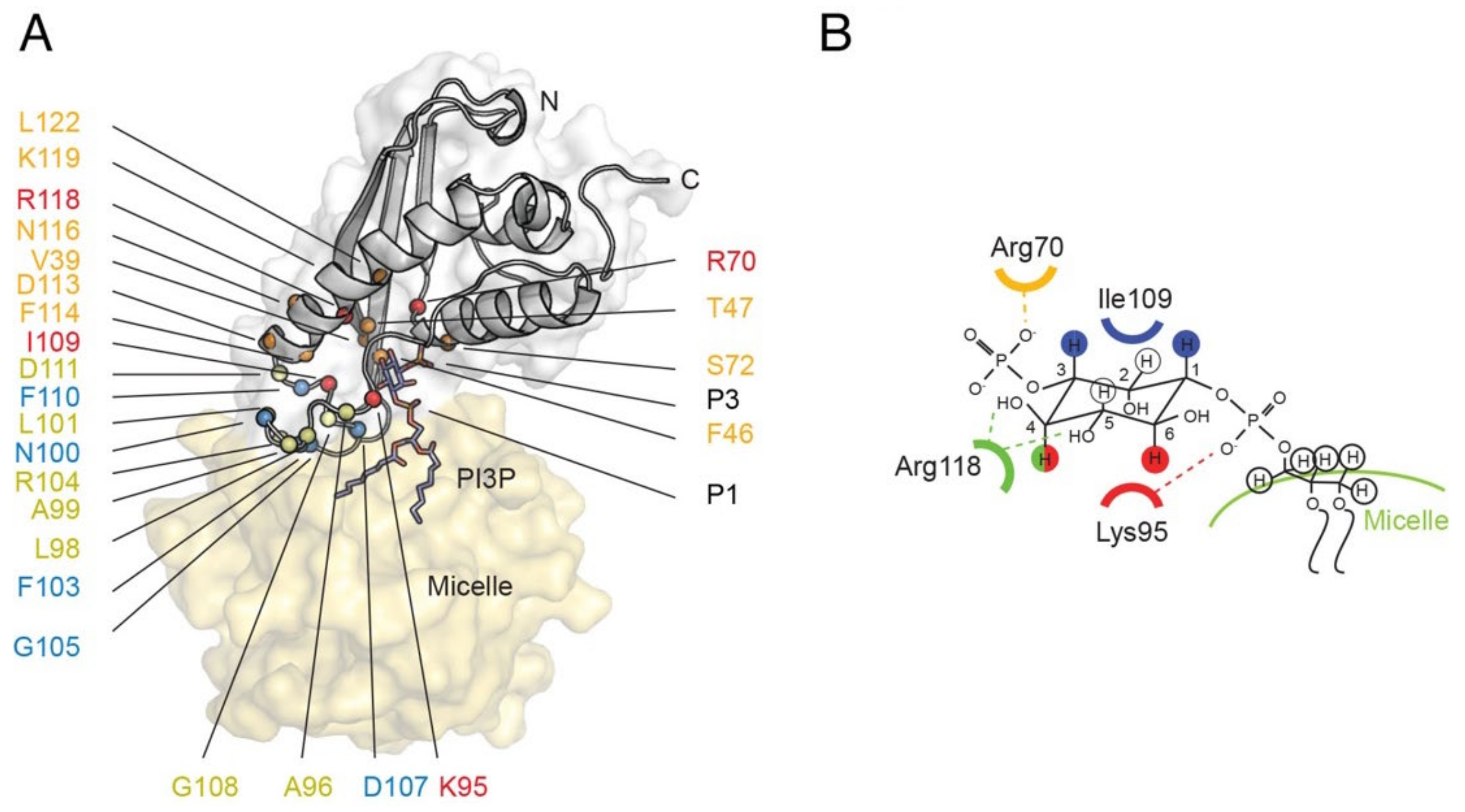
3.5. Sorting Nexins and PX Proteins
3.6. FYVE Protein-Containing Protrudin
3.7. Sec14 Domain-Containing MSPD2 Protein
3.8. WD Repeat-Containing PREB Protein
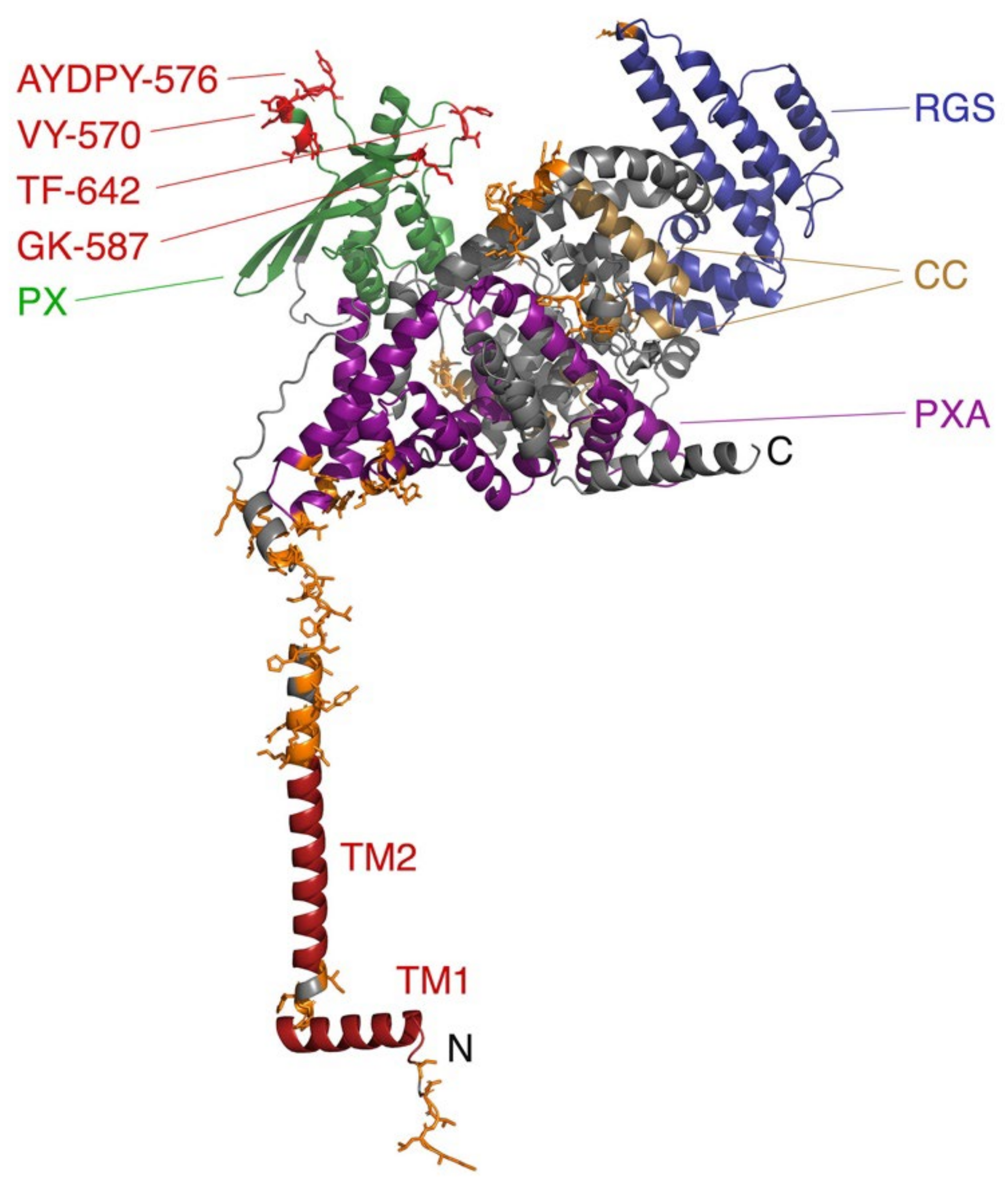
3.9. GTPase Domain-Containing Miro Proteins
3.10. MATH Domain-Containing Proteins
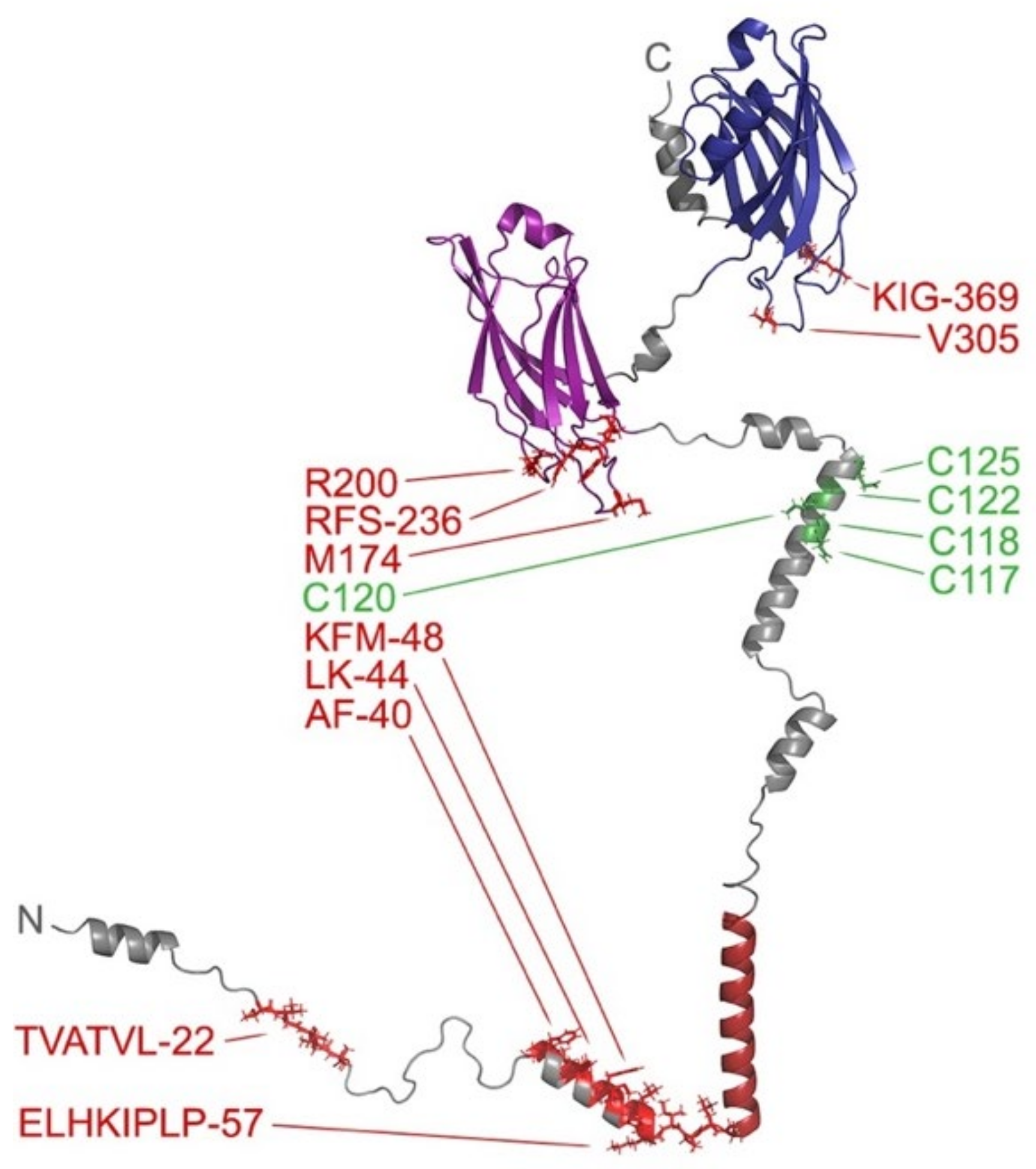
3.11. Spike Proteins
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Reis, R.; Moraes, I. Structural biology and structure-function relationships of membrane proteins. Biochem. Soc. Trans. 2019, 47, 47–61. [Google Scholar] [CrossRef] [PubMed]
- Overduin, M.; Kervin, T.A. The phosphoinositide code is read by a plethora of protein domains. Expert Rev. Proteom. 2021, 18, 483–502. [Google Scholar] [CrossRef] [PubMed]
- Larsen, A.H.; John, L.H.; Sansom, M.S.P.; Corey, R.A. Specific interactions of peripheral membrane proteins with lipids: What can molecular simulations show us? Biosci. Rep. 2022, 42, BSR20211406. [Google Scholar] [CrossRef] [PubMed]
- Tran, A.; Kervin, T.A.; Overduin, M. Multifaceted membrane binding head of the SARS-CoV-2 spike protein. Curr. Res. Struct. Biol. 2022, 4, 146–157. [Google Scholar] [CrossRef] [PubMed]
- Overduin, M.; Kervin, T.A.; Tran, A. Progressive membrane-binding mechanism of SARS-CoV-2 variant spike proteins. iScience 2022, 25, 104722. [Google Scholar] [CrossRef]
- Overduin, M.; Esmaili, M. Memtein: The fundamental unit of membrane-protein structure and function. Chem. Phys. Lipids 2019, 218, 73–84. [Google Scholar] [CrossRef] [PubMed]
- Overduin, M.; Trieber, C.; Prosser, R.S.; Picard, L.P.; Sheff, J.G. Structures and Dynamics of Native-State Transmembrane Protein Targets and Bound Lipids. Membranes 2021, 11, 451. [Google Scholar] [CrossRef]
- Varadi, M.; Anyango, S.; Deshpande, M.; Nair, S.; Natassia, C.; Yordanova, G.; Yuan, D.; Stroe, O.; Wood, G.; Laydon, A.; et al. AlphaFold Protein Structure Database: Massively expanding the structural coverage of protein-sequence space with high-accuracy models. Nucleic Acids Res. 2022, 50, D439–D444. [Google Scholar] [CrossRef]
- Baek, M.; DiMaio, F.; Anishchenko, I.; Dauparas, J.; Ovchinnikov, S.; Lee, G.R.; Wang, J.; Cong, Q.; Kinch, L.N.; Schaeffer, R.D.; et al. Accurate prediction of protein structures and interactions using a three-track neural network. Science 2021, 373, 871–876. [Google Scholar] [CrossRef]
- Tunyasuvunakool, K.; Adler, J.; Wu, Z.; Green, T.; Zielinski, M.; Žídek, A.; Bridgland, A.; Cowie, A.; Meyer, C.; Laydon, A.; et al. Highly accurate protein structure prediction for the human proteome. Nature 2021, 596, 590–596. [Google Scholar] [CrossRef]
- Lomize, M.A.; Pogozheva, I.D.; Joo, H.; Mosberg, H.I.; Lomize, A.L. OPM database and PPM web server: Resources for positioning of proteins in membranes. Nucleic Acids Res. 2012, 40, D370–D376. [Google Scholar] [CrossRef] [PubMed]
- Lomize, A.L.; Todd, S.C.; Pogozheva, I.D. Spatial arrangement of proteins in planar and curved membranes by PPM 3.0. Protein Sci. 2022, 31, 209–220. [Google Scholar] [CrossRef] [PubMed]
- Schramm, C.A.; Hannigan, B.T.; Donald, J.E.; Keasar, C.; Saven, J.G.; Degrado, W.F.; Samish, I. Knowledge-based potential for positioning membrane-associated structures and assessing residue-specific energetic contributions. Structure 2012, 20, 924–935. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kufareva, I.; Lenoir, M.; Dancea, F.; Sridhar, P.; Raush, E.; Bissig, C.; Gruenberg, J.; Abagyan, R.; Overduin, M. Discovery of novel membrane binding structures and functions. Biochem. Cell Biol. 2014, 92, 555–563. [Google Scholar] [CrossRef] [Green Version]
- Bissig, C.; Lenoir, M.; Velluz, M.C.; Kufareva, I.; Abagyan, R.; Overduin, M.; Gruenberg, J. Viral infection controlled by a calcium-dependent lipid-binding module in ALIX. Dev. Cell 2013, 25, 364–373. [Google Scholar] [CrossRef] [Green Version]
- Bryant, J.A.; Morris, F.C.; Knowles, T.J.; Maderbocus, R.; Heinz, E.; Boelter, G.; Alodaini, D.; Colyer, A.; Wotherspoon, P.J.; Staunton, K.A.; et al. Structure of dual-BON domain protein DolP identifies phospholipid binding as a new mechanism for protein localization. eLife 2020, 9, e62614. [Google Scholar] [CrossRef]
- Overduin, M.; Wille, H.; Westaway, D. Multisite interactions of prions with membranes and native nanodiscs. Chem. Phys. Lipids 2021, 236, 105063. [Google Scholar] [CrossRef]
- Lenoir, M.; Kufareva, I.; Abagyan, R.; Overduin, M. Membrane and Protein Interactions of the Pleckstrin Homology Domain Superfamily. Membranes 2015, 5, 646–663. [Google Scholar] [CrossRef]
- De Castro, E.; Sigrist, C.J.; Gattiker, A.; Bulliard, V.; Langendijk-Genevaux, P.S.; Gasteiger, E.; Bairoch, A.; Hulo, N. ScanProsite: Detection of PROSITE signature matches and ProRule-associated functional and structural residues in proteins. Nucleic Acids Res. 2006, 34, W362–W365. [Google Scholar] [CrossRef]
- Letunic, I.; Khedkar, S.; Bork, P. SMART: Recent updates, new developments and status in 2020. Nucleic Acids Res. 2021, 49, D458–D460. [Google Scholar] [CrossRef]
- Hiller, K.; Grote, A.; Scheer, M.; Münch, R.; Jahn, D. PrediSi: Prediction of signal peptides and their cleavage positions. Nucleic Acids Res. 2004, 32, W375–W379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teufel, F.; Almagro Armenteros, J.J.; Johansen, A.R.; Gíslason, M.H.; Pihl, S.I.; Tsirigos, K.D.; Winther, O.; Brunak, S.; von Heijne, G.; Nielsen, H. SignalP 6.0 predicts all five types of signal peptides using protein language models. Nat. Biotechnol. 2022, 40, 1023–1025. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.; Zheng, Y.; Li, H.; Luo, X.; He, Z.; Cao, S.; Shi, Y.; Zhao, Q.; Xue, Y.; Zuo, Z.; et al. GPS-Lipid: A robust tool for the prediction of multiple lipid modification sites. Sci. Rep. 2016, 6, 28249. [Google Scholar] [CrossRef] [PubMed]
- Berman, H.M.; Westbrook, J.; Feng, Z.; Gilliland, G.; Bhat, T.N.; Weissig, H.; Shindyalov, I.N.; Bourne, P.E. The Protein Data Bank. Nucleic Acids Res. 2000, 28, 235–242. [Google Scholar] [CrossRef] [Green Version]
- Lomize, A.L.; Lomize, M.A.; Krolicki, S.R.; Pogozheva, I.D. Membranome: A database for proteome-wide analysis of single-pass membrane proteins. Nucleic Acids Res. 2017, 45, D250–D255. [Google Scholar] [CrossRef] [Green Version]
- Kozma, D.; Simon, I.; Tusnády, G.E. PDBTM: Protein Data Bank of transmembrane proteins after 8 years. Nucleic Acids Res. 2013, 41, D524–D529. [Google Scholar] [CrossRef] [Green Version]
- Newport, T.D.; Sansom, M.S.P.; Stansfeld, P.J. The MemProtMD database: A resource for membrane-embedded protein structures and their lipid interactions. Nucleic Acids Res. 2019, 47, D390–D397. [Google Scholar] [CrossRef] [Green Version]
- White, S.H. Biophysical dissection of membrane proteins. Nature 2009, 459, 344–346. [Google Scholar] [CrossRef]
- DeLano, W.L. The PyMOL Molecular Graphics System, Version 1.8; Schrödinger LLC: New York, NY, USA, 2014.
- Käll, L.; Krogh, A.; Sonnhammer, E.L. An HMM posterior decoder for sequence feature prediction that includes homology information. Bioinformatics 2005, 21, i251–i257. [Google Scholar] [CrossRef]
- Tsaousis, G.N.; Bagos, P.G.; Hamodrakas, S.J. HMMpTM: Improving transmembrane protein topology prediction using phosphorylation and glycosylation site prediction. Biochim. Biophys. Acta 2014, 1844, 316–322. [Google Scholar] [CrossRef]
- Zimmermann, L.; Stephens, A.; Nam, S.Z.; Rau, D.; Kübler, J.; Lozajic, M.; Gabler, F.; Söding, J.; Lupas, A.N.; Alva, V. A Completely Reimplemented MPI Bioinformatics Toolkit with a New HHpred Server at its Core. J. Mol. Biol. 2018, 430, 2237–2243. [Google Scholar] [CrossRef] [PubMed]
- Lees, J.A.; Messa, M.; Sun, E.W.; Wheeler, H.; Torta, F.; Wenk, M.R.; de Camilli, P.; Reinisch, K.M. Lipid transport by TMEM24 at ER-plasma membrane contacts regulates pulsatile insulin secretion. Science 2017, 355, eaah6171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, E.W.; Guillén-Samander, A.; Bian, X.; Wu, Y.; Cai, Y.; Messa, M.; De Camilli, P. Lipid transporter TMEM24/C2CD2L is a Ca2+-regulated component of ER-plasma membrane contacts in mammalian neurons. Proc. Natl. Acad. Sci. USA 2019, 116, 5775–5784. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bozelli, J.C.; Yune, J.; Aulakh, S.S.; Cao, Z.; Fernandes, A.; Seitova, A.; Tong, Y.; Schreier, S.; Epand, R.M. Human Diacylglycerol Kinase ε N-Terminal Segment Regulates the Phosphatidylinositol Cycle, Controlling the Rate but Not the Acyl Chain Composition of Its Lipid Intermediates. ACS Chem. Biol. 2022, 17, 2495–2506. [Google Scholar] [CrossRef] [PubMed]
- Bozelli, J.C.; Yune, J.; Hou, Y.H.; Chatha, P.; Fernandes, A.; Cao, Z.; Tong, Y.; Epand, R.M. Regulation of DGKε Activity and Substrate Acyl Chain Specificity by Negatively Charged Phospholipids. Biophys. J. 2020, 118, 957–966. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, N.; Hozumi, Y.; Ito, T.; Hosoya, T.; Kondo, H.; Goto, K. Differential subcellular targeting and activity-dependent subcellular localization of diacylglycerol kinase isozymes in transfected cells. Eur. J. Cell Biol. 2007, 86, 433–444. [Google Scholar] [CrossRef]
- Therrien, C.; Di Fulvio, S.; Pickles, S.; Sinnreich, M. Characterization of lipid binding specificities of dysferlin C2 domains reveals novel interactions with phosphoinositides. Biochemistry 2009, 48, 2377–2384. [Google Scholar] [CrossRef]
- Redpath, G.M.; Sophocleous, R.A.; Turnbull, L.; Whitchurch, C.B.; Cooper, S.T. Ferlins Show Tissue-Specific Expression and Segregate as Plasma Membrane/Late Endosomal or Trans-Golgi/Recycling Ferlins. Traffic 2016, 17, 245–266. [Google Scholar] [CrossRef] [Green Version]
- Xu, L.; Pallikkuth, S.; Hou, Z.; Mignery, G.A.; Robia, S.L.; Han, R. Dysferlin forms a dimer mediated by the C2 domains and the transmembrane domain in vitro and in living cells. PLoS ONE 2011, 6, e27884. [Google Scholar] [CrossRef] [Green Version]
- Giordano, F.; Saheki, Y.; Idevall-Hagren, O.; Colombo, S.F.; Pirruccello, M.; Milosevic, I.; Gracheva, E.O.; Bagriantsev, S.N.; Borgese, N.; De Camilli, P. PI(4,5)P(2)-dependent and Ca(2+)-regulated ER-PM interactions mediated by the extended synaptotagmins. Cell 2013, 153, 1494–1509. [Google Scholar] [CrossRef]
- Chang, C.L.; Hsieh, T.S.; Yang, T.T.; Rothberg, K.G.; Azizoglu, D.B.; Volk, E.; Liao, J.C.; Liou, J. Feedback regulation of receptor-induced Ca2+ signaling mediated by E-Syt1 and Nir2 at endoplasmic reticulum-plasma membrane junctions. Cell Rep. 2013, 5, 813–825. [Google Scholar] [CrossRef] [Green Version]
- Schauder, C.M.; Wu, X.; Saheki, Y.; Narayanaswamy, P.; Torta, F.; Wenk, M.R.; De Camilli, P.; Reinisch, K.M. Structure of a lipid-bound extended synaptotagmin indicates a role in lipid transfer. Nature 2014, 510, 552–555. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naito, T.; Ercan, B.; Krshnan, L.; Triebl, A.; Koh, D.H.Z.; Wei, F.Y.; Tomizawa, K.; Torta, F.T.; Wenk, M.R.; Saheki, Y. Movement of accessible plasma membrane cholesterol by the GRAMD1 lipid transfer protein complex. eLife 2019, 8, e51401. [Google Scholar] [CrossRef] [PubMed]
- Laraia, L.; Friese, A.; Corkery, D.P.; Konstantinidis, G.; Erwin, N.; Hofer, W.; Karatas, H.; Klewer, L.; Brockmeyer, A.; Metz, M.; et al. The cholesterol transfer protein GRAMD1A regulates autophagosome biogenesis. Nat. Chem. Biol. 2019, 15, 710–720. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horenkamp, F.A.; Valverde, D.P.; Nunnari, J.; Reinisch, K.M. Molecular basis for sterol transport by StART-like lipid transfer domains. EMBO J. 2018, 37, e98002. [Google Scholar] [CrossRef]
- Ercan, B.; Naito, T.; Koh, D.H.Z.; Dharmawan, D.; Saheki, Y. Molecular basis of accessible plasma membrane cholesterol recognition by the GRAM domain of GRAMD1b. EMBO J. 2021, 40, e106524. [Google Scholar] [CrossRef]
- Besprozvannaya, M.; Dickson, E.; Li, H.; Ginburg, K.S.; Bers, D.M.; Auwerx, J.; Nunnari, J. GRAM domain proteins specialize functionally distinct ER-PM contact sites in human cells. eLife 2018, 7, e31019. [Google Scholar] [CrossRef]
- John, K.; Alla, V.; Meier, C.; Pützer, B.M. GRAMD4 mimics p53 and mediates the apoptotic function of p73 at mitochondria. Cell Death Differ. 2011, 18, 874–886. [Google Scholar] [CrossRef]
- Parui, A.L.; Mishra, V.; Dutta, S.; Bhaumik, P.; Bose, K. Inter-subunit crosstalk via PDZ synergistically governs allosteric activation of proapoptotic HtrA2. Structure 2022, 30, 1307–1320.e5. [Google Scholar] [CrossRef]
- Gomes, G.; do Amaral, M.J.; Bagri, K.M.; Vasconcellos, L.M.; Almeida, M.D.S.; Alvares, L.E.; Mermelstein, C. New Findings on LMO7 Transcripts, Proteins and Regulatory Regions in Human and Vertebrate Model Organisms and the Intracellular Distribution in Skeletal Muscle Cells. Int. J. Mol. Sci. 2021, 22, 12885. [Google Scholar] [CrossRef]
- Téllez-Arreola, J.L.; Martínez-Torres, A.; Flores-Moran, A.E.; Lazaro-Guevara, J.M.; Estrada-Mondragón, A. Analysis of the MCTP Amino Acid Sequence Reveals the Conservation of Putative Calcium- and Lipid-Binding Pockets Within the C2 Domains In Silico. J. Mol. Evol. 2022, 90, 271–282. [Google Scholar] [CrossRef] [PubMed]
- Shin, O.H.; Han, W.; Wang, Y.; Südhof, T.C. Evolutionarily conserved multiple C2 domain proteins with two transmembrane regions (MCTPs) and unusual Ca2+ binding properties. J. Biol. Chem. 2005, 280, 1641–1651. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Genç, Ö.; Dickman, D.K.; Ma, W.; Tong, A.; Fetter, R.D.; Davis, G.W. MCTP is an ER-resident calcium sensor that stabilizes synaptic transmission and homeostatic plasticity. eLife 2017, 6, e22904. [Google Scholar] [CrossRef] [PubMed]
- Bertenshaw, G.P.; Norcum, M.T.; Bond, J.S. Structure of homo- and hetero-oligomeric meprin metalloproteases. Dimers, tetramers, and high molecular mass multimers. J. Biol. Chem. 2003, 278, 2522–2532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Covill-Cooke, C.; Toncheva, V.S.; Drew, J.; Birsa, N.; López-Doménech, G.; Kittler, J.T. Peroxisomal fission is modulated by the mitochondrial Rho-GTPases, Miro1 and Miro2. EMBO Rep. 2020, 21, e49865. [Google Scholar] [CrossRef] [PubMed]
- Fransson, A.; Ruusala, A.; Aspenström, P. Atypical Rho GTPases have roles in mitochondrial homeostasis and apoptosis. J. Biol. Chem. 2003, 278, 6495–6502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, K.P.; Focia, P.J.; Chakravarthy, S.; Landahl, E.C.; Klosowiak, J.L.; Rice, S.E.; Freymann, D.M. Insight into human Miro1/2 domain organization based on the structure of its N-terminal GTPase. J. Struct. Biol. 2020, 212, 107656. [Google Scholar] [CrossRef]
- Costello, J.L.; Castro, I.G.; Camões, F.; Schrader, T.A.; McNeall, D.; Yang, J.; Giannopoulou, E.A.; Gomes, S.; Pogenberg, V.; Bonekamp, N.A.; et al. Predicting the targeting of tail-anchored proteins to subcellular compartments in mammalian cells. J. Cell Sci. 2017, 130, 1675–1687. [Google Scholar] [CrossRef] [Green Version]
- Zouiouich, M.; Di Mattia, T.; Martinet, A.; Eichler, J.; Wendling, C.; Tomishige, N.; Grandgirard, E.; Fuggetta, N.; Fromental-Ramain, C.; Mizzon, G.; et al. MOSPD2 is an endoplasmic reticulum-lipid droplet tether functioning in LD homeostasis. J. Cell Biol. 2022, 221, e202110044. [Google Scholar] [CrossRef]
- Di Mattia, T.; Wilhelm, L.P.; Ikhlef, S.; Wendling, C.; Spehner, D.; Nominé, Y.; Giordano, F.; Mathelin, C.; Drin, G.; Tomasetto, C.; et al. Identification of MOSPD2, a novel scaffold for endoplasmic reticulum membrane contact sites. EMBO Rep. 2018, 19, e45453. [Google Scholar] [CrossRef]
- Chung, J.; Torta, F.; Masai, K.; Lucast, L.; Czapla, H.; Tanner, L.B.; Narayanaswamy, P.; Wenk, M.R.; Nakatsu, F.; De Camilli, P. Intracellular Transport. PI4P/phosphatidylserine countertransport at ORP5- and ORP8-mediated ER-plasma membrane contacts. Science 2015, 349, 428–432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghai, R.; Du, X.; Wang, H.; Dong, J.; Ferguson, C.; Brown, A.J.; Parton, R.G.; Wu, J.W.; Yang, H. ORP5 and ORP8 bind phosphatidylinositol-4, 5-biphosphate (PtdIns(4,5)P. Nat. Commun. 2017, 8, 757. [Google Scholar] [CrossRef] [Green Version]
- Du, X.; Zhou, L.; Aw, Y.C.; Mak, H.Y.; Xu, Y.; Rae, J.; Wang, W.; Zadoorian, A.; Hancock, S.E.; Osborne, B.; et al. ORP5 localizes to ER-lipid droplet contacts and regulates the level of PI(4)P on lipid droplets. J. Cell Biol. 2020, 219, e201905162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guyard, V.; Monteiro-Cardoso, V.F.; Omrane, M.; Sauvanet, C.; Houcine, A.; Boulogne, C.; Ben Mbarek, K.; Vitale, N.; Faklaris, O.; El Khallouki, N.; et al. ORP5 and ORP8 orchestrate lipid droplet biogenesis and maintenance at ER-mitochondria contact sites. J. Cell Biol. 2022, 221, e202112107. [Google Scholar] [CrossRef] [PubMed]
- Zhou, T.; Li, S.; Zhong, W.; Vihervaara, T.; Béaslas, O.; Perttilä, J.; Luo, W.; Jiang, Y.; Lehto, M.; Olkkonen, V.M.; et al. OSBP-related protein 8 (ORP8) regulates plasma and liver tissue lipid levels and interacts with the nucleoporin Nup62. PLoS ONE 2011, 6, e21078. [Google Scholar] [CrossRef] [Green Version]
- Padmanarayana, M.; Hams, N.; Speight, L.C.; Petersson, E.J.; Mehl, R.A.; Johnson, C.P. Characterization of the lipid binding properties of Otoferlin reveals specific interactions between PI(4,5)P2 and the C2C and C2F domains. Biochemistry 2014, 53, 5023–5033. [Google Scholar] [CrossRef] [Green Version]
- Jeyasimman, D.; Ercan, B.; Dharmawan, D.; Naito, T.; Sun, J.; Saheki, Y. PDZD-8 and TEX-2 regulate endosomal PI(4,5)P. Nat. Commun. 2021, 12, 6065. [Google Scholar] [CrossRef]
- Shirane, M.; Wada, M.; Morita, K.; Hayashi, N.; Kunimatsu, R.; Matsumoto, Y.; Matsuzaki, F.; Nakatsumi, H.; Ohta, K.; Tamura, Y.; et al. Protrudin and PDZD8 contribute to neuronal integrity by promoting lipid extraction required for endosome maturation. Nat. Commun. 2020, 11, 4576. [Google Scholar] [CrossRef]
- Krappa, R.; Nguyen, A.; Burrola, P.; Deretic, D.; Lemke, G. Evectins: Vesicular proteins that carry a pleckstrin homology domain and localize to post-Golgi membranes. Proc. Natl. Acad. Sci. USA 1999, 96, 4633–4638. [Google Scholar] [CrossRef] [Green Version]
- Okazaki, S.; Kato, R.; Uchida, Y.; Taguchi, T.; Arai, H.; Wakatsuki, S. Structural basis of the strict phospholipid binding specificity of the pleckstrin homology domain of human evectin-2. Acta Crystallogr. D Biol. Crystallogr. 2012, 68, 117–123. [Google Scholar] [CrossRef]
- Uchida, Y.; Hasegawa, J.; Chinnapen, D.; Inoue, T.; Okazaki, S.; Kato, R.; Wakatsuki, S.; Misaki, R.; Koike, M.; Uchiyama, Y.; et al. Intracellular phosphatidylserine is essential for retrograde membrane traffic through endosomes. Proc. Natl. Acad. Sci. USA 2011, 108, 15846–15851. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, S.; Ladak, R.; Swanson, D.A.; Soltyk, A.; Sun, H.; Ploder, L.; Vidgen, D.; Duncan, A.M.; Garami, E.; Valle, D.; et al. PHR1 encodes an abundant, pleckstrin homology domain-containing integral membrane protein in the photoreceptor outer segments. J. Biol. Chem. 1999, 274, 35676–35685. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Centonze, F.G.; Reiterer, V.; Nalbach, K.; Saito, K.; Pawlowski, K.; Behrends, C.; Farhan, H. LTK is an ER-resident receptor tyrosine kinase that regulates secretion. J. Cell Biol. 2019, 218, 2470–2480. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Zhang, M.; Ge, L. A new type of membrane contact in the ER-Golgi system regulates autophagosome biogenesis. Autophagy 2021, 17, 4499–4501. [Google Scholar] [CrossRef]
- Raiborg, C.; Wenzel, E.M.; Pedersen, N.M.; Olsvik, H.; Schink, K.O.; Schultz, S.W.; Vietri, M.; Nisi, V.; Bucci, C.; Brech, A.; et al. Repeated ER-endosome contacts promote endosome translocation and neurite outgrowth. Nature 2015, 520, 234–238. [Google Scholar] [CrossRef]
- Gil, J.E.; Kim, E.; Kim, I.S.; Ku, B.; Park, W.S.; Oh, B.H.; Ryu, S.H.; Cho, W.; Heo, W.D. Phosphoinositides differentially regulate protrudin localization through the FYVE domain. J. Biol. Chem. 2012, 287, 41268–41276. [Google Scholar] [CrossRef] [Green Version]
- Chandra, M.; Chin, Y.K.; Mas, C.; Feathers, J.R.; Paul, B.; Datta, S.; Chen, K.E.; Jia, X.; Yang, Z.; Norwood, S.J.; et al. Classification of the human phox homology (PX) domains based on their phosphoinositide binding specificities. Nat. Commun. 2019, 10, 1528. [Google Scholar] [CrossRef] [Green Version]
- Saric, A.; Freeman, S.A.; Williamson, C.D.; Jarnik, M.; Guardia, C.M.; Fernandopulle, M.S.; Gershlick, D.C.; Bonifacino, J.S. SNX19 restricts endolysosome motility through contacts with the endoplasmic reticulum. Nat. Commun. 2021, 12, 4552. [Google Scholar] [CrossRef]
- Henne, W.M.; Zhu, L.; Balogi, Z.; Stefan, C.; Pleiss, J.A.; Emr, S.D. Mdm1/Snx13 is a novel ER-endolysosomal interorganelle tethering protein. J. Cell Biol. 2015, 210, 541–551. [Google Scholar] [CrossRef] [Green Version]
- Lu, A.; Hsieh, F.; Sharma, B.R.; Vaughn, S.R.; Enrich, C.; Pfeffer, S.R. CRISPR screens for lipid regulators reveal a role for ER-bound SNX13 in lysosomal cholesterol export. J. Cell Biol. 2022, 221, e202105060. [Google Scholar] [CrossRef]
- Mas, C.; Norwood, S.J.; Bugarcic, A.; Kinna, G.; Leneva, N.; Kovtun, O.; Ghai, R.; Ona Yanez, L.E.; Davis, J.L.; Teasdale, R.D.; et al. Structural basis for different phosphoinositide specificities of the PX domains of sorting nexins regulating G-protein signaling. J. Biol. Chem. 2014, 289, 28554–28568. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Datta, S.; Bowerman, J.; Hariri, H.; Ugrankar, R.; Eckert, K.M.; Corley, C.; Vale, G.; McDonald, J.G.; Henne, W.M. Snx14 proximity labeling reveals a role in saturated fatty acid metabolism and ER homeostasis defective in SCAR20 disease. Proc. Natl. Acad. Sci. USA 2020, 117, 33282–33294. [Google Scholar] [CrossRef] [PubMed]
- Datta, S.; Liu, Y.; Hariri, H.; Bowerman, J.; Henne, W.M. Cerebellar ataxia disease-associated Snx14 promotes lipid droplet growth at ER-droplet contacts. J. Cell Biol. 2019, 218, 1335–1351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bard, M.P.; Hegmans, J.P.; Hemmes, A.; Luider, T.M.; Willemsen, R.; Severijnen, L.A.; van Meerbeeck, J.P.; Burgers, S.A.; Hoogsteden, H.C.; Lambrecht, B.N. Proteomic analysis of exosomes isolated from human malignant pleural effusions. Am. J. Respir. Cell Mol. Biol. 2004, 31, 114–121. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Chen, R.; Dong, Y.; Zhu, J.; Su, K.; Liu, J.; Xu, J. Structural Studies Reveal Unique Non-canonical Regulators of G Protein Signaling Homology (RH) Domains in Sorting Nexins. J. Mol. Biol. 2022, 434, 167823. [Google Scholar] [CrossRef] [PubMed]
- Ugrankar, R.; Bowerman, J.; Hariri, H.; Chandra, M.; Chen, K.; Bossanyi, M.F.; Datta, S.; Rogers, S.; Eckert, K.M.; Vale, G.; et al. Drosophila Snazarus Regulates a Lipid Droplet Population at Plasma Membrane-Droplet Contacts in Adipocytes. Dev. Cell 2019, 50, 557–572.e5. [Google Scholar] [CrossRef]
- Wolfes, A.C.; Dean, C. The diversity of synaptotagmin isoforms. Curr. Opin. Neurobiol. 2020, 63, 198–209. [Google Scholar] [CrossRef] [PubMed]
- Tran, H.T.; Anderson, L.H.; Knight, J.D. Membrane-Binding Cooperativity and Coinsertion by C2AB Tandem Domains of Synaptotagmins 1 and 7. Biophys. J. 2019, 116, 1025–1036. [Google Scholar] [CrossRef] [Green Version]
- Honigmann, A.; van den Bogaart, G.; Iraheta, E.; Risselada, H.J.; Milovanovic, D.; Mueller, V.; Müllar, S.; Diederichsen, U.; Fasshauer, D.; Grubmüller, H.; et al. Phosphatidylinositol 4,5-bisphosphate clusters act as molecular beacons for vesicle recruitment. Nat. Struct. Mol. Biol. 2013, 20, 679–686. [Google Scholar] [CrossRef] [Green Version]
- Bai, J.; Tucker, W.C.; Chapman, E.R. PIP2 increases the speed of response of synaptotagmin and steers its membrane-penetration activity toward the plasma membrane. Nat. Struct. Mol. Biol. 2004, 11, 36–44. [Google Scholar] [CrossRef]
- Zanetti, M.N.; Bello, O.D.; Wang, J.; Coleman, J.; Cai, Y.; Sindelar, C.V.; Rothman, J.E.; Krishnakumar, S.S. Ring-like oligomers of Synaptotagmins and related C2 domain proteins. eLife 2016, 5, e17262. [Google Scholar] [CrossRef] [PubMed]
- Fukuda, M.; Kojima, T.; Mikoshiba, K. Phospholipid composition dependence of Ca2+-dependent phospholipid binding to the C2A domain of synaptotagmin IV. J. Biol. Chem. 1996, 271, 8430–8434. [Google Scholar] [CrossRef] [Green Version]
- Voleti, R.; Tomchick, D.R.; Südhof, T.C.; Rizo, J. Exceptionally tight membrane-binding may explain the key role of the synaptotagmin-7 C. Proc. Natl. Acad. Sci. USA 2017, 114, E8518–E8527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, L.K.; Choudhary, V.; Toulmay, A.; Prinz, W.A. An inducible ER-Golgi tether facilitates ceramide transport to alleviate lipotoxicity. J. Cell Biol. 2017, 216, 131–147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toulmay, A.; Prinz, W.A. A conserved membrane-binding domain targets proteins to organelle contact sites. J. Cell Sci. 2012, 125, 49–58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grushin, K.; Wang, J.; Coleman, J.; Rothman, J.E.; Sindelar, C.V.; Krishnakumar, S.S. Structural basis for the clamping and Ca. Nat. Commun. 2019, 10, 2413. [Google Scholar] [CrossRef] [Green Version]
- Ruchala, P.; Waring, A.J.; Cilluffo, M.; Whitelegge, J.P.; Gundersen, C.B. Insights into the structure and molecular topography of the fatty acylated domain of synaptotagmin-1. Biochim. Biophys. Acta Biomembr. 2019, 1861, 677–684. [Google Scholar] [CrossRef]
- Harsini, F.M.; Chebrolu, S.; Fuson, K.L.; White, M.A.; Rice, A.M.; Sutton, R.B. FerA is a Membrane-Associating Four-Helix Bundle Domain in the Ferlin Family of Membrane-Fusion Proteins. Sci. Rep. 2018, 8, 10949. [Google Scholar] [CrossRef] [Green Version]
- Muriel, J.; Lukyanenko, V.; Kwiatkowski, T.; Bhattacharya, S.; Garman, D.; Weisleder, N.; Bloch, R.J. The C2 domains of dysferlin: Roles in membrane localization, Ca. J. Physiol. 2022, 600, 1953–1968. [Google Scholar] [CrossRef]
- Ware, T.B.; Franks, C.E.; Granade, M.E.; Zhang, M.; Kim, K.B.; Park, K.S.; Gahlmann, A.; Harris, T.E.; Hsu, K.L. Reprogramming fatty acyl specificity of lipid kinases via C1 domain engineering. Nat. Chem. Biol. 2020, 16, 170–178. [Google Scholar] [CrossRef]
- Chen, Y.; Sheng, R.; Kallberg, M.; Silkov, A.; Tun, M.P.; Bhardwaj, N.; Kurilova, S.; Hall, R.A.; Honig, B.; Lu, H.; et al. Genome-wide functional annotation of dual-specificity protein- and lipid-binding modules that regulate protein interactions. Mol. Cell 2012, 46, 226–237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, H.; Chen, L.; Tan, L.; Im, Y.J. Structural basis of human PDZD8-Rab7 interaction for the ER-late endosome tethering. Sci. Rep. 2021, 11, 18859. [Google Scholar] [CrossRef] [PubMed]
- Isakoff, S.J.; Cardozo, T.; Andreev, J.; Li, Z.; Ferguson, K.M.; Abagyan, R.; Lemmon, M.A.; Aronheim, A.; Skolnik, E.Y. Identification and analysis of PH domain-containing targets of phosphatidylinositol 3-kinase using a novel in vivo assay in yeast. EMBO J. 1998, 17, 5374–5387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lenoir, M.; Coskun, U.; Grzybek, M.; Cao, X.; Buschhorn, S.B.; James, J.; Simon, K.; Overduin, M. Structural basis of wedging the Golgi membrane by FAPP pleckstrin homology domains. EMBO Reports 2010, 11, 279–284. [Google Scholar] [CrossRef]
- Jian, X.; Tang, W.K.; Zhai, P.; Roy, N.S.; Luo, R.; Gruschus, J.M.; Yohe, M.E.; Chen, P.W.; Li, Y.; Byrd, R.A.; et al. Molecular Basis for Cooperative Binding of Anionic Phospholipids to the PH Domain of the Arf GAP ASAP1. Structure 2015, 23, 1977–1988. [Google Scholar] [CrossRef] [Green Version]
- Tong, J.; Manik, M.K.; Im, Y.J. Structural basis of sterol recognition and nonvesicular transport by lipid transfer proteins anchored at membrane contact sites. Proc. Natl. Acad. Sci. USA 2018, 115, E856–E865. [Google Scholar] [CrossRef] [Green Version]
- Zheng, B.; Ma, Y.C.; Ostrom, R.S.; Lavoie, C.; Gill, G.N.; Insel, P.A.; Huang, X.Y.; Farquhar, M.G. RGS-PX1, a GAP for GalphaS and sorting nexin in vesicular trafficking. Science 2001, 294, 1939–1942. [Google Scholar] [CrossRef]
- Ha, C.M.; Park, D.; Kim, Y.; Na, M.; Panda, S.; Won, S.; Kim, H.; Ryu, H.; Park, Z.Y.; Rasenick, M.M.; et al. SNX14 is a bifunctional negative regulator for neuronal 5-HT6 receptor signaling. J. Cell Sci. 2015, 128, 1848–1861. [Google Scholar] [CrossRef] [Green Version]
- Tiu, A.C.; Yang, J.; Asico, L.D.; Konkalmatt, P.; Zheng, X.; Cuevas, S.; Wang, X.; Lee, H.; Mazhar, M.; Felder, R.A.; et al. Lipid rafts are required for effective renal D. FASEB J. 2020, 34, 6999–7017. [Google Scholar] [CrossRef] [Green Version]
- Ishii, M.; Inanobe, A.; Kurachi, Y. PIP3 inhibition of RGS protein and its reversal by Ca2+/calmodulin mediate voltage-dependent control of the G protein cycle in a cardiac K+ channel. Proc. Natl. Acad. Sci. USA 2002, 99, 4325–4330. [Google Scholar] [CrossRef]
- Zhou, C.Z.; De La Sierra-Gallay, I.L.; Quevillon-Cheruel, S.; Collinet, B.; Minard, P.; Blondeau, K.; Henckes, G.; Aufrère, R.; Leulliot, N.; Graille, M.; et al. Crystal Structure of the Yeast Phox Homology (PX) Domain Protein Grd19p Complexed to Phosphatidylinositol-3-phosphate. J. Biol. Chem. 2003, 278, 50371–50376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bravo, J.; Karathanassis, D.; Pacold, C.M.; Pacold, M.E.; Ellson, C.D.; Anderson, K.E.; Butler, P.J.; Lavenir, I.; Perisic, O.; Hawkins, P.T.; et al. The crystal structure of the PX domain from p40(phox) bound to phosphatidylinositol 3-phosphate. Mol. Cell 2001, 8, 829–839. [Google Scholar] [CrossRef]
- Karathanassis, D.; Stahelin, R.V.; Bravo, J.; Perisic, O.; Pacold, C.M.; Cho, W.; Williams, R.L. Binding of the PX domain of p47phoxto phosphatidylinositol 3, 4-bisphosphate and phosphatidic acid is masked by an intramolecular interaction. EMBO J. 2002, 21, 5057–5068. [Google Scholar] [CrossRef] [PubMed]
- Stampoulis, P.; Ueda, T.; Matsumoto, M.; Terasawa, H.; Miyano, K.; Sumimoto, H.; Shimada, I. Atypical membrane-embedded phosphatidylinositol 3,4-bisphosphate (PI(3,4)P2)-binding site on p47(phox) Phox homology (PX) domain revealed by NMR. J. Biol. Chem. 2012, 287, 17848–17859. [Google Scholar] [CrossRef] [Green Version]
- Lenoir, M.; Ustunel, C.; Rajesh, S.; Kaur, J.; Moreau, D.; Gruenberg, J.; Overduin, M. Phosphorylation of conserved phosphoinositide binding pocket regulates sorting nexin membrane targeting. Nat. Commun. 2018, 9, 993. [Google Scholar] [CrossRef]
- Pylypenko, O.; Lundmark, R.; Rasmuson, E.; Carlsson, S.R.; Rak, A. The PX-BAR membrane-remodeling unit of sorting nexin 9. EMBO J. 2007, 26, 4788–4800. [Google Scholar] [CrossRef] [Green Version]
- Cheever, M.L.; Sato, T.K.; de Beer, T.; Kutateladze, T.G.; Emr, S.D.; Overduin, M. Phox domain interaction with PtdIns(3)P targets the Vam7 t-SNARE to vacuole membranes. Nat. Cell Biol. 2001, 3, 613–618. [Google Scholar] [CrossRef]
- Dancea, F.; Kami, K.; Overduin, M. Lipid interaction networks of peripheral membrane proteins revealed by data-driven micelle docking. Biophys. J. 2008, 94, 515–524. [Google Scholar] [CrossRef] [Green Version]
- Xu, T.; Gan, Q.; Wu, B.; Yin, M.; Xu, J.; Shu, X.; Liu, J. Molecular Basis for PI(3,5)P2 Recognition by SNX11, a Protein Involved in Lysosomal Degradation and Endosome Homeostasis Regulation. J. Mol. Biol. 2020, 432, 4750–4761. [Google Scholar] [CrossRef]
- Leneva, N.; Kovtun, O.; Morado, D.R.; Briggs, J.A.G.; Owen, D.J. Architecture and mechanism of metazoan retromer:SNX3 tubular coat assembly. Sci. Adv. 2021, 7, eabf859. [Google Scholar] [CrossRef]
- Saita, S.; Shirane, M.; Natume, T.; Iemura, S.I.; Nakayama, K.I. Promotion of neurite extension by protrudin requires its interaction with vesicle-associated membrane protein-associated protein. J. Biol. Chem. 2009, 284, 13766–13777. [Google Scholar] [CrossRef] [Green Version]
- Matsuzaki, F.; Shirane, M.; Matsumoto, M.; Nakayama, K.I. Protrudin serves as an adaptor molecule that connects KIF5 and its cargoes in vesicular transport during process formation. Mol. Biol. Cell 2011, 22, 4602–4620. [Google Scholar] [CrossRef]
- Panagabko, C.; Morley, S.; Hernandez, M.; Cassolato, P.; Gordon, H.; Parsons, R.; Manor, D.; Atkinson, J. Ligand specificity in the CRAL-TRIO protein family. Biochemistry 2003, 42, 6467–6474. [Google Scholar] [CrossRef]
- Miller, M.B.; Vishwanatha, K.S.; Mains, R.E.; Eipper, B.A. An N-terminal Amphipathic Helix Binds Phosphoinositides and Enhances Kalirin Sec14 Domain-mediated Membrane Interactions. J. Biol. Chem. 2015, 290, 13541–13555. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seto, S.; Tsujimura, K.; Koide, Y. Coronin-1a inhibits autophagosome formation around Mycobacterium tuberculosis-containing phagosomes and assists mycobacterial survival in macrophages. Cell. Microbiol. 2012, 14, 710–727. [Google Scholar] [CrossRef] [PubMed]
- Tsujita, K.; Itoh, T.; Kondo, A.; Oyama, M.; Kozuka-Hata, H.; Irino, Y.; Hasegawa, J.; Takenawa, T. Proteome of acidic phospholipid-binding proteins: Spatial and temporal regulation of Coronin 1A by phosphoinositides. J. Biol. Chem. 2010, 285, 6781–6789. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joiner, A.M.N.; Fromme, J.C. Structural basis for the initiation of COPII vesicle biogenesis. Structure 2021, 29, 859–872.e6. [Google Scholar] [CrossRef]
- Klosowiak, J.L.; Park, S.; Smith, K.P.; French, M.E.; Focia, P.J.; Freymann, D.M.; Rice, S.E. Structural insights into Parkin substrate lysine targeting from minimal Miro substrates. Sci. Rep. 2016, 6, 33019. [Google Scholar] [CrossRef] [Green Version]
- Cao, S.; Chung, S.; Kim, S.; Li, Z.; Manor, D.; Buck, M. K-Ras G-domain binding with signaling lipid phosphatidylinositol (4,5)-phosphate (PIP2): Membrane association, protein orientation, and function. J. Biol. Chem. 2019, 294, 7068–7084. [Google Scholar] [CrossRef] [Green Version]
- Lee, K.Y.; Fang, Z.; Enomoto, M.; Gasmi-Seabrook, G.; Zheng, L.; Koide, S.; Ikura, M.; Marshall, C.B. Two Distinct Structures of Membrane-Associated Homodimers of GTP- and GDP-Bound KRAS4B Revealed by Paramagnetic Relaxation Enhancement. Angew. Chem. Int. Ed. Engl. 2020, 59, 11037–11045. [Google Scholar] [CrossRef]
- Modi, S.; López-Doménech, G.; Halff, E.F.; Covill-Cooke, C.; Ivankovic, D.; Melandri, D.; Arancibia-Cárcamo, I.L.; Burden, J.J.; Lowe, A.R.; Kittler, J.T. Miro clusters regulate ER-mitochondria contact sites and link cristae organization to the mitochondrial transport machinery. Nat. Commun. 2019, 10, 4399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rousseau, A.; McEwen, A.G.; Poussin-Courmontagne, P.; Rognan, D.; Nomine, Y.; Rio, M.C.; Tomasetto, C.; Alpy, F. TRAF4 is a novel phosphoinositide-binding protein modulating tight junctions and favoring cell migration. PLoS Biol. 2013, 11, e1001726. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lottaz, D.; Hahn, D.; Müller, S.; Müller, C.; Sterchi, E.E. Secretion of human meprin from intestinal epithelial cells depends on differential expression of the alpha and beta subunits. Eur. J. Biochem. 1999, 259, 496–504. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, S.; Oneda, B.; Yap, L.M.; Jewell, D.P.; Matters, G.L.; Fitzpatrick, L.R.; Seibold, F.; Sterchi, E.E.; Ahmad, T.; Lottaz, D.; et al. MEP1A allele for meprin A metalloprotease is a susceptibility gene for inflammatory bowel disease. Mucosal. Immunol. 2009, 2, 220–231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rochman, N.D.; Wolf, Y.I.; Koonin, E.V. Molecular adaptations during viral epidemics. EMBO Rep. 2022, 23, e55393. [Google Scholar] [CrossRef]
- Martinez-Molledo, M.; Nji, E.; Reyes, N. Structural insights into the lysophospholipid brain uptake mechanism and its inhibition by syncytin-2. Nat. Struct. Mol. Biol. 2022, 29, 604–612. [Google Scholar] [CrossRef]
- Wong, L.H.; Gatta, A.T.; Levine, T.P. Lipid transfer proteins: The lipid commute via shuttles, bridges and tubes. Nat. Rev. Mol. Cell Biol. 2019, 20, 85–101. [Google Scholar] [CrossRef]
- Scheidt, H.A.; Kolocaj, K.; Veje Kristensen, J.; Huster, D.; Langosch, D. Transmembrane Helix Induces Membrane Fusion through Lipid Binding and Splay. J. Phys. Chem. Lett. 2018, 9, 3181–3186. [Google Scholar] [CrossRef]
- Churchward, M.A.; Rogasevskaia, T.; Brandman, D.M.; Khosravani, H.; Nava, P.; Atkinson, J.K.; Coorssen, J.R. Specific lipids supply critical negative spontaneous curvature—An essential component of native Ca2+-triggered membrane fusion. Biophys. J. 2008, 94, 3976–3986. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.A.; Eyeson, R.; Cheever, M.L.; Geng, J.; Verkhusha, V.V.; Burd, C.; Overduin, M.; Kutateladze, T.G. Targeting of the FYVE domain to endosomal membranes is regulated by a histidine switch. Proc. Natl. Acad. Sci. USA 2005, 102, 13052–13057. [Google Scholar] [CrossRef]
- Milovanovic, D.; Platen, M.; Junius, M.; Diederichsen, U.; Schaap, I.A.; Honigmann, A.; Jahn, R.; van den Bogaart, G. Calcium Promotes the Formation of Syntaxin 1 Mesoscale Domains through Phosphatidylinositol 4,5-Bisphosphate. J. Biol. Chem. 2016, 291, 7868–7876. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vardar, G.; Salazar-Lázaro, A.; Zobel, S.; Trimbuch, T.; Rosenmund, C. Syntaxin-1A modulates vesicle fusion in mammalian neurons via juxtamembrane domain dependent palmitoylation of its transmembrane domain. eLife 2022, 11, e78182. [Google Scholar] [CrossRef] [PubMed]
- Ma, H.; Lou, Y.; Lin, W.H.; Xue, H.W. MORN motifs in plant PIPKs are involved in the regulation of subcellular localization and phospholipid binding. Cell Res. 2006, 16, 466–478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sajko, S.; Grishkovskaya, I.; Kostan, J.; Graewert, M.; Setiawan, K.; Trübestein, L.; Niedermüller, K.; Gehin, C.; Sponga, A.; Puchinger, M.; et al. Structures of three MORN repeat proteins and a re-evaluation of the proposed lipid-binding properties of MORN repeats. PLoS ONE 2020, 15, e0242677. [Google Scholar] [CrossRef] [PubMed]
- Maass, K.; Fischer, M.A.; Seiler, M.; Temmerman, K.; Nickel, W.; Seedorf, M. A signal comprising a basic cluster and an amphipathic alpha-helix interacts with lipids and is required for the transport of Ist2 to the yeast cortical ER. J. Cell Sci. 2009, 122, 625–635. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fischer, M.A.; Temmerman, K.; Ercan, E.; Nickel, W.; Seedorf, M. Binding of plasma membrane lipids recruits the yeast integral membrane protein Ist2 to the cortical ER. Traffic 2009, 10, 1084–1097. [Google Scholar] [CrossRef] [PubMed]
- De Jesús-Pérez, J.J.; Cruz-Rangel, S.; Espino-Saldaña, Á.; Martínez-Torres, A.; Qu, Z.; Hartzell, H.C.; Corral-Fernandez, N.E.; Pérez-Cornejo, P.; Arreola, J. Phosphatidylinositol 4,5-bisphosphate, cholesterol, and fatty acids modulate the calcium-activated chloride channel TMEM16A (ANO1). Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2018, 1863, 299–312. [Google Scholar] [CrossRef]
- Le, S.C.; Jia, Z.; Chen, J.; Yang, H. Molecular basis of PIP. Nat. Commun. 2019, 10, 3769. [Google Scholar] [CrossRef] [Green Version]
- Bushell, S.R.; Pike, A.C.W.; Falzone, M.E.; Rorsman, N.J.G.; Ta, C.M.; Corey, R.A.; Newport, T.D.; Christianson, J.C.; Scofano, L.F.; Shintre, C.A.; et al. The structural basis of lipid scrambling and inactivation in the endoplasmic reticulum scramblase TMEM16K. Nat. Commun. 2019, 10, 3956. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein | Uniprot | TM | MR | Lipid Ligands | Oligomer | Function | Location | References |
|---|---|---|---|---|---|---|---|---|
| C2CD2 | Q9Y426 | 1α-N | SMP, C2 | PI | lipid transfer | ER | [33] | |
| C2CD2L | O14523 | 1α-N | SMP, C2 | PI | lipid transfer | ER, PM | [34] | |
| DGKε | P52429 | 1α-N | C1 (2) | DAG | lipid kinase | ER, PM | [35,36,37] | |
| DYSF | O75923 | 1α-C | C2 (7) | PtdSer, PI4P, PI(4,5)P2 (Ca2+) | dimer | PM, LE, LY | [38,39,40] | |
| ESyt1 | Q9BSJ8 | 2α-N | SMP, C2 (5) | PI(4,5)P2 (Ca2+) | dimer | lipid transfer | ER, PM | [41,42] |
| ESyt2 | A0FGR8 | 2α-N | SMP, C2 (3) | PI(4,5)P2 (Ca2+) | dimer | lipid transfer | ER, PM | [41,43] |
| ESyt3 | A0FGR9 | 2α-N | SMP, C2 (3) | PI(4,5)P2 (Ca2+) | dimer | lipid transfer | ER, PM | [41] |
| FR1L4 | A9Z1Z3 | 1α-C | C2 (6) | cc | ||||
| FR1L5 | A0AVI2 | 1α-C | C2 (7) | |||||
| FR1L6 | Q2WGJ9 | 1α-C | C2 (6) | cc | PM, TGN | [39,44] | ||
| GRAMD1A | Q96CP6 | 1α-C | GRAM, StART | cholesterol | oligomer | lipid transfer | AP, ER, PM | [44,45,46] |
| GRAMD1B | Q3KR37 | 1α-C | GRAM, StART | PtdSer, cholesterol | oligomer | lipid transfer | ER, PM | [46,47] |
| GRAMD1C | Q8IYS0 | 1α-C | GRAM, StART | cholesterol | cc | lipid transfer | ER, PM | [46] |
| GRAMD2A | Q8IUY3 | 1α-C | GRAM | PI(4,5)P2 | calcium entry | ER, PM | [48] | |
| GRAMD2B | Q96HH9 | 1α-C | GRAM | cc | ||||
| GRAMD4 | Q6IC98 | 4α-N | GRAM | cc | membrane permeabilization | ER, MOM | [49] | |
| HTRA2 | O43464 | 1α-N | PDZ | protease | [50] | |||
| LMO7 | E9PMS6 | 1α-N | PDZ | cc | PM, NE | [51] | ||
| MCTP1 | Q6DN14 | 2α-C | C2 (3) | ER, SV | [52,53,54] | |||
| MCTP2 | Q6DN12 | 2α-C | C2 (3) | cc | ER, SV | [52,53,54] | ||
| Mep1A | Q16819 | 1α-C | MATH | oligomer | metalloprotease | EC, PM | [55] | |
| Miro1 | Q8IXI2 | 1α-C | GTPase (2) | dimer | membrane transport | MOM, PO | [56,57,58] | |
| Miro2 | Q8IXI1 | 1α-C | GTPase (2) | membrane transport | ER, MOM, PO | [56,57,59] | ||
| MSPD2 | Q8NHP6 | 1α-C | CRAL-Trio | phospholipid defects | cc | ER, E, G, MOM, LD | [60,61] | |
| MYOF | Q9NZM1 | 1α-C | C2 (7) | cc | PM, LE, LY | [39] | ||
| ORP5 | Q9H0X9 | 1α-C | PH | PtdSer, PI4P, PIP2, PIP3 | oligomer | lipid transfer | ER, LD, PM | [62,63,64,65] |
| ORP8 | Q9BZF1 | 1α-C | PH | PtdSer, PI4P, PIP2, PIP3 | oligomer | lipid transfer | ER, PM, NE | [62,63,66] |
| OTOF | Q9HC10 | 1α-C | C2 (7) | PtdSer, PI(4,5)P2 (Ca2+) | cc | PM, SV | [67] | |
| PDZD8 | Q8NEN9 | 1α-N | SMP, PDZ, C1 | PtdSer, PI4P, PI(4,5)P2 | cc | lipid transfer | ER, LE, LY, PM | [68,69] |
| PLEKHB1 | Q9UF11 | 1α-C | PH | [70] | ||||
| PLEKHB2 | Q96CS7 | 1α-C | PH | PtdSer | RE | [71,72,73] | ||
| PREB | Q9HCU5 | 1α-C | WD | ER export | ER, G | [74,75] | ||
| Protrudin | Q5T4F4 | 4α-N | FYVE | PIP2, PIP3 | oligomer | lipid transfer | ER, LE | [69,76,77] |
| Snx13 | Q9Y5W8 | 2α-N | RGS, PX | cholesterol, PI3P, PI(3,4)P2 | dimer | lipid transport | EL, ER, LY | [78,79,80,81,82] |
| Snx14 | Q9Y5W7 | 2α-N | RGS, PX | lipid homeostasis | ER, LD | [78,79,82,83,84] | ||
| Snx19 | Q92543 | 2α-N | PX | PI3P | EL, ER, LD | [78,79,82] | ||
| Snx25 | Q9H3E2 | 1α-C | RGS, PX | PIP2, PIP3 | dimer | lipid homeostasis | ER, LD, PM | [78,82,85,86,87] |
| Spike | P0DTC2 | 1α-C | NTD, RBD | trimer | membrane fusion | E, EV, PM, VM | [4,5] | |
| SYJ2B | P57105 | 1α-C | PDZ | MOM | ||||
| Syt1 | P21579 | 1α-N | C2 (2) | PtdSer, PI(4,5)P2 (Ca2+) | oligomer | membrane fusion | PRCO, SV | [88,89,90,91] |
| Syt2 | Q8N9I0 | 1α-N | C2 (2) | PtdSer, PI(4,5)P2 (Ca2+) | oligomer | membrane fusion | SV | [88,92,93] |
| Syt3 | Q9BQG1 | 1α-N | C2 (2) | PtdSer/PI | oligomer | membrane fusion | PM, PRCO | [88,93] |
| Syt4 | Q9H2B2 | 1α-N | C2 (2) | PtdSer, PI (Ca2+) | oligomer | SDSV, TV | [88,93] | |
| Syt5 | O00445 | 1α-N | C2 (2) | PtdSer, PI (Ca2+) | oligomer | membrane fusion | SDSV, SV | [88,93] |
| Syt6 | Q5T7P8 | 1α-N | C2 (2) | PtdSer, PI (Ca2+) | oligomer | membrane fusion | PM, SDSV | [88,93] |
| Syt7 | O43581 | 1α-N | C2 (2) | PI(4,5)P2 (Ca2+) | oligomer | membrane fusion | PM | [88,89,92,94] |
| Syt8 | Q8NBV8 | 1α-N | C2 (2) | oligomer | [88] | |||
| Syt9 | Q86SS6 | 1α-N | C2 (2) | PI(4,5)P2 (Ca2+) | oligomer | membrane fusion | SDSV, SV | [88,92] |
| Syt10 | Q6XYQ8 | 1α-N | C2 (2) | oligomer | membrane fusion | TV | [88] | |
| Syt11 | Q9BT88 | 1α-N | C2 (2) | oligomer | [88] | |||
| Syt12 | Q8IV01 | 1α-N | C2 (2) | oligomer | SV | [88] | ||
| Syt13 | Q7L8C5 | 1α-N | C2 (2) | [88] | ||||
| Syt14 | Q8NB59 | 1α-N | C2 (2) | oligomer | [88] | |||
| Syt15 | Q9BQS2 | 1α-N | C2 (2) | oligomer | [88] | |||
| TEX2 | Q8IWB9 | 2α-N | PH, SMP | PI(4,5)P2 | lipid transfer | ER | [68,95,96] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Overduin, M.; Tran, A.; Eekels, D.M.; Overduin, F.; Kervin, T.A. Transmembrane Membrane Readers form a Novel Class of Proteins That Include Peripheral Phosphoinositide Recognition Domains and Viral Spikes. Membranes 2022, 12, 1161. https://doi.org/10.3390/membranes12111161
Overduin M, Tran A, Eekels DM, Overduin F, Kervin TA. Transmembrane Membrane Readers form a Novel Class of Proteins That Include Peripheral Phosphoinositide Recognition Domains and Viral Spikes. Membranes. 2022; 12(11):1161. https://doi.org/10.3390/membranes12111161
Chicago/Turabian StyleOverduin, Michael, Anh Tran, Dominic M. Eekels, Finn Overduin, and Troy A. Kervin. 2022. "Transmembrane Membrane Readers form a Novel Class of Proteins That Include Peripheral Phosphoinositide Recognition Domains and Viral Spikes" Membranes 12, no. 11: 1161. https://doi.org/10.3390/membranes12111161
APA StyleOverduin, M., Tran, A., Eekels, D. M., Overduin, F., & Kervin, T. A. (2022). Transmembrane Membrane Readers form a Novel Class of Proteins That Include Peripheral Phosphoinositide Recognition Domains and Viral Spikes. Membranes, 12(11), 1161. https://doi.org/10.3390/membranes12111161