
Insight into the Structure, Functions, and Dynamics of the Leptospira Outer Membrane Proteins with the Pathogenicity


{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Leptospira Membrane Structure and the Virulence Factors
2.1. LPS
2.2. LipL32
2.3. LipL21
2.4. Loa22
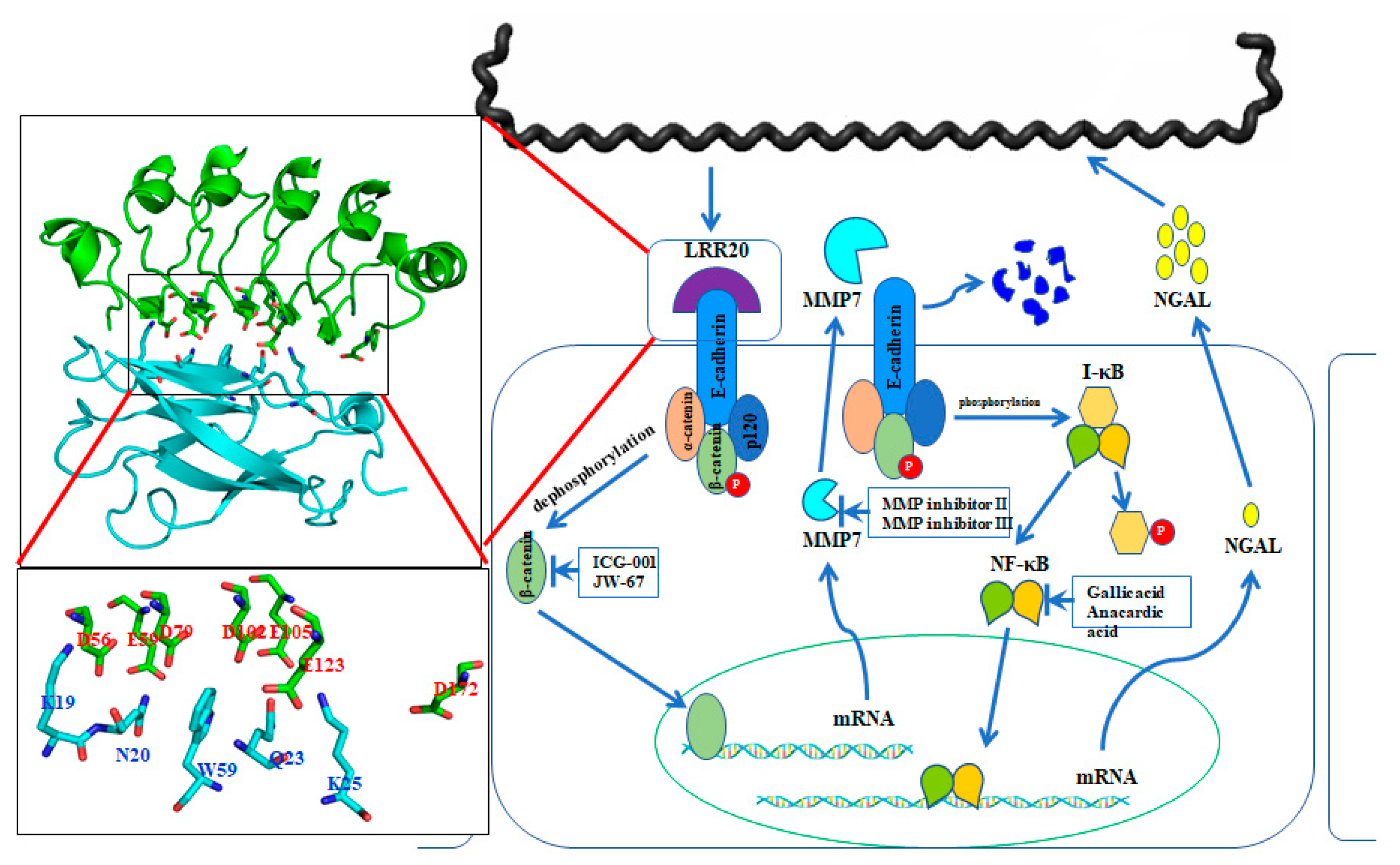
2.5. LRR20
2.6. LipL71
2.7. LigA and LigB
2.8. LipL41
3. The Diagnosis of Leptospira
3.1. Serological Diagnosis of Leptospirosis
3.2. Bacteria and Molecules Diagnosis of Leptospirosis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bharti, A.R.; Nally, J.E.; Ricaldi, J.N.; Matthias, M.A.; Diaz, M.M.; Lovett, M.A.; Levett, P.N.; Gilman, R.H.; Willig, M.R.; Gotuzzo, E.; et al. Leptospirosis: A zoonotic disease of global importance. Lancet Infect. Dis. 2003, 3, 757–771. [Google Scholar] [CrossRef]
- Adler, B.; de la Pena Moctezuma, A. Leptospira and leptospirosis. Vet. Microbiol. 2010, 140, 287–296. [Google Scholar] [CrossRef]
- Guerra, M.A. Leptospirosis: Public health perspectives. Biol. J. Int. Assoc. Biol. Stand. 2013, 41, 295–297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hattori, T.; Chagan-Yasutan, H.; Shiratori, B.; Egawa, S.; Izumi, T.; Kubo, T.; Nakajima, C.; Suzuki, Y.; Niki, T.; Alisjahbana, B.; et al. Development of point-of-care testing for disaster-related infectious diseases. Tohoku J. Exp. Med. 2016, 238, 287–293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, C.W. Leptospirosis renal disease: Emerging culprit of chronic kidney disease unknown etiology. Nephron 2017, 138. [Google Scholar] [CrossRef] [PubMed]
- Johnston, V.; Stockley, J.M.; Dockrell, D.; Warrell, D.; Bailey, R.; Pasvol, G.; Klein, J.; Ustianowski, A.; Jones, M.; Beeching, N.J.; et al. Fever in returned travellers presenting in the United Kingdom: Recommendations for investigation and initial management. J. Infect. 2009, 59, 1–18. [Google Scholar] [CrossRef]
- Yang, C.W. Leptospirosis renal disease: Understanding the initiation by Toll-like receptors. Kidney Int. 2007, 72, 918–925. [Google Scholar] [CrossRef] [Green Version]
- Costa, F.; Hagan, J.E.; Calcagno, J.; Kane, M.; Torgerson, P.; Martinez-Silveira, M.S.; Stein, C.; Abela-Ridder, B.; Ko, A.I. Global morbidity and mortality of leptospirosis: A systematic review. PLoS Negl. Trop. Dis. 2015, 9, e0003898. [Google Scholar] [CrossRef] [Green Version]
- Yang, C.W. Leptospirosis in Taiwan—An underestimated infectious disease. Chang. Gung Med. J. 2007, 30, 109–115. [Google Scholar]
- Sharp, T.M.; Rivera Garcia, B.; Perez-Padilla, J.; Galloway, R.L.; Guerra, M.; Ryff, K.R.; Haberling, D.; Ramakrishnan, S.; Shadomy, S.; Blau, D.; et al. Early Indicators of Fatal Leptospirosis during the 2010 Epidemic in Puerto Rico. PLoS Negl. Trop. Dis. 2016, 10, e0004482. [Google Scholar] [CrossRef]
- Yang, H.Y.; Hung, C.C.; Liu, S.H.; Guo, Y.G.; Chen, Y.C.; Ko, Y.C.; Huang, C.T.; Chou, L.F.; Tian, Y.C.; Chang, M.Y.; et al. Overlooked risk for chronic kidney disease after leptospiral infection: A population-based survey and epidemiological cohort evidence. PLoS Negl. Trop. Dis. 2015, 9, e0004105. [Google Scholar] [CrossRef] [PubMed]
- Cagliero, J.; Villanueva, S.Y.A.M.; Matsui, M. Leptospirosis pathophysiology: Into the storm of cytokines. Front. Cell Infect. Microbiol. 2018, 8, 204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Senavirathna, I.; Rathish, D.; Agampodi, S. Cytokine response in human leptospirosis with different clinical outcomes: A systematic review. BMC Infect. Dis. 2020, 20, 268. [Google Scholar] [CrossRef] [PubMed]
- Dickson, K.; Lehmann, C. Inflammatory response to different toxins in experimental sepsis models. Int. J. Mol. Sci. 2019, 20, 4341. [Google Scholar] [CrossRef] [Green Version]
- Fernandes, L.G.; Siqueira, G.H.; Teixeira, A.R.; Silva, L.P.; Figueredo, J.M.; Cosate, M.R.; Vieira, M.L.; Nascimento, A.L. Leptospira spp.: Novel insights into host-pathogen interactions. Vet. Immunol. Immunopathol. 2016, 176, 50–57. [Google Scholar] [CrossRef]
- Vieira, M.L.; Herwald, H.; Nascimento, A. The interplay between host haemostatic systems and Leptospira spp. infections. Crit. Rev. Microbiol. 2020, 46, 121–135. [Google Scholar] [CrossRef]
- Daroz, B.B.; Fernandes, L.G.V.; Cavenague, M.F.; Kochi, L.T.; Passalia, F.J.; Takahashi, M.B.; Nascimento Filho, E.G.; Teixeira, A.F.; Nascimento, A. A review on host-Leptospira Interactions: What we know and future expectations. Front. Cell Infect. Microbiol. 2021, 11, 777709. [Google Scholar] [CrossRef]
- Haake, D.A.; Levett, P.N. Leptospirosis in humans. Leptospira Leptospirosis 2015, 387, 65–97. [Google Scholar] [CrossRef] [Green Version]
- Yang, C.W.; Wu, M.S.; Pan, M.J. Leptospirosis renal disease. Nephrol. Dial. Transpl. 2001, 16 (Suppl. 5), 73–77. [Google Scholar] [CrossRef]
- Haake, D.A. Spirochaetal lipoproteins and pathogenesis. Microbiology 2000, 146 Pt 7, 1491–1504. [Google Scholar] [CrossRef] [Green Version]
- Shimizu, T.; Matsusaka, E.; Takayanagi, K.; Masuzawa, T.; Iwamoto, Y.; Morita, T.; Mifuchi, I.; Yanagihara, Y. Biological activities of lipopolysaccharide-like substance (LLS) extracted from Leptospira interrogans serovar canicola strain Moulton. Microbiol. Immunol. 1987, 31, 727–735. [Google Scholar] [CrossRef] [PubMed]
- Raetz, C.R.; Whitfield, C. Lipopolysaccharide endotoxins. Annu Rev. Biochem. 2002, 71, 635–700. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Werts, C.; Tapping, R.I.; Mathison, J.C.; Chuang, T.H.; Kravchenko, V.; Saint Girons, I.; Haake, D.A.; Godowski, P.J.; Hayashi, F.; Ozinsky, A.; et al. Leptospiral lipopolysaccharide activates cells through a TLR2-dependent mechanism. Nat. Immunol. 2001, 2, 346–352. [Google Scholar] [CrossRef]
- Weidel, W.; Pelzer, H. Bagshaped macromolecules-a new outlook on bacterial cell walls. Adv. Enzym. Relat. Subj. Biochem. 1964, 26, 193–232. [Google Scholar] [CrossRef]
- Dhar, S.; Kumari, H.; Balasubramanian, D.; Mathee, K. Cell-wall recycling and synthesis in Escherichia coli and Pseudomonas aeruginosa—Their role in the development of resistance. J. Med. Microbiol. 2018, 67, 1–21. [Google Scholar] [CrossRef]
- Hung, C.C.; Chang, C.T.; Tian, Y.C.; Wu, M.S.; Yu, C.C.; Pan, M.J.; Vandewalle, A.; Yang, C.W. Leptospiral membrane proteins stimulate pro-inflammatory chemokines secretion by renal tubule epithelial cells through toll-like receptor 2 and p38 mitogen activated protein kinase. Nephrol. Dial. Transpl. 2006, 21, 898–910. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cullen, P.A.; Haake, D.A.; Adler, B. Outer membrane proteins of pathogenic spirochetes. FEMS Microbiol. Rev. 2004, 28, 291–318. [Google Scholar] [CrossRef] [Green Version]
- Santecchia, I.; Ferrer, M.F.; Vieira, M.L.; Gomez, R.M.; Werts, C. Phagocyte escape of Leptospira: The role of TLRs and NLRs. Front. Immunol. 2020, 11, 571816. [Google Scholar] [CrossRef]
- Murray, G.L.; Srikram, A.; Henry, R.; Hartskeerl, R.A.; Sermswan, R.W.; Adler, B. Mutations affecting Leptospira interrogans lipopolysaccharide attenuate virulence. Mol. Microbiol. 2010, 78, 701–709. [Google Scholar] [CrossRef]
- Murray, G.L.; Srikram, A.; Hoke, D.E.; Wunder, E.A., Jr.; Henry, R.; Lo, M.; Zhang, K.; Sermswan, R.W.; Ko, A.I.; Adler, B. Major surface protein LipL32 is not required for either acute or chronic infection with Leptospira interrogans. Infect. Immun. 2009, 77, 952–958. [Google Scholar] [CrossRef] [Green Version]
- Ristow, P.; Bourhy, P.; da Cruz McBride, F.W.; Figueira, C.P.; Huerre, M.; Ave, P.; Girons, I.S.; Ko, A.I.; Picardeau, M. The OmpA-like protein Loa22 is essential for leptospiral virulence. PLoS Pathog. 2007, 3, e97. [Google Scholar] [CrossRef] [PubMed]
- King, A.M.; Bartpho, T.; Sermswan, R.W.; Bulach, D.M.; Eshghi, A.; Picardeau, M.; Adler, B.; Murray, G.L. Leptospiral outer membrane protein LipL41 is not essential for acute leptospirosis but requires a small chaperone protein, lep, for stable expression. Infect. Immun. 2013, 81, 2768–2776. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, K.; Murray, G.L.; Seemann, T.; Srikram, A.; Bartpho, T.; Sermswan, R.W.; Adler, B.; Hoke, D.E. Leptospiral LruA is required for virulence and modulates an interaction with mammalian apolipoprotein AI. Infect. Immun. 2013, 81, 3872–3879. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pappas, C.J.; Picardeau, M. Control of gene expression in Leptospira spp. by transcription activator-like effectors demonstrates a potential role for LigA and LigB in Leptospira interrogans Virulence. Appl. Environ. Microbiol. 2015, 81, 7888–7892. [Google Scholar] [CrossRef] [Green Version]
- Croda, J.; Figueira, C.P.; Wunder, E.A., Jr.; Santos, C.S.; Reis, M.G.; Ko, A.I.; Picardeau, M. Targeted mutagenesis in pathogenic Leptospira species: Disruption of the LigB gene does not affect virulence in animal models of leptospirosis. Infect. Immun. 2008, 76, 5826–5833. [Google Scholar] [CrossRef] [Green Version]
- Ratet, G.; Santecchia, I.; Fanton d’Andon, M.; Vernel-Pauillac, F.; Wheeler, R.; Lenormand, P.; Fischer, F.; Lechat, P.; Haake, D.A.; Picardeau, M.; et al. LipL21 lipoprotein binding to peptidoglycan enables Leptospira interrogans to escape NOD1 and NOD2 recognition. PLoS Pathog. 2017, 13, e1006725. [Google Scholar] [CrossRef] [Green Version]
- Ren, S.X.; Fu, G.; Jiang, X.G.; Zeng, R.; Miao, Y.G.; Xu, H.; Zhang, Y.X.; Xiong, H.; Lu, G.; Lu, L.F.; et al. Unique physiological and pathogenic features of Leptospira interrogans revealed by whole-genome sequencing. Nature 2003, 422, 888–893. [Google Scholar] [CrossRef]
- Chou, L.F.; Chen, Y.T.; Lu, C.W.; Ko, Y.C.; Tang, C.Y.; Pan, M.J.; Tian, Y.C.; Chiu, C.H.; Hung, C.C.; Yang, C.W. Sequence of Leptospira santarosai serovar Shermani genome and prediction of virulence-associated genes. Gene 2012, 511, 364–370. [Google Scholar] [CrossRef]
- Mitchison, M.; Bulach, D.M.; Vinh, T.; Rajakumar, K.; Faine, S.; Adler, B. Identification and characterization of the dTDP-rhamnose biosynthesis and transfer genes of the lipopolysaccharide-related rfb locus in Leptospira interrogans serovar Copenhageni. J. Bacteriol. 1997, 179, 1262–1267. [Google Scholar] [CrossRef] [Green Version]
- Vinh, T.; Adler, B.; Faine, S. Ultrastructure and chemical composition of lipopolysaccharide extracted from Leptospira interrogans serovar Copenhageni. J. Gen. Microbiol. 1986, 132, 103–109. [Google Scholar] [CrossRef] [Green Version]
- Nascimento, A.L.; Ko, A.I.; Martins, E.A.; Monteiro-Vitorello, C.B.; Ho, P.L.; Haake, D.A.; Verjovski-Almeida, S.; Hartskeerl, R.A.; Marques, M.V.; Oliveira, M.C.; et al. Comparative genomics of two Leptospira interrogans serovars reveals novel insights into physiology and pathogenesis. J. Bacteriol. 2004, 186, 2164–2172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lo, Y.Y.; Hsu, S.H.; Ko, Y.C.; Hung, C.C.; Chang, M.Y.; Hsu, H.H.; Pan, M.J.; Chen, Y.W.; Lee, C.H.; Tseng, F.G.; et al. Essential calcium-binding cluster of Leptospira LipL32 protein for inflammatory responses through the Toll-like receptor 2 pathway. J. Biol. Chem. 2013, 288, 12335–12344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hauk, P.; Guzzo, C.R.; Roman Ramos, H.; Ho, P.L.; Farah, C.S. Structure and calcium-binding activity of LipL32, the major surface antigen of pathogenic Leptospira sp. J. Mol. Biol. 2009, 390, 722–736. [Google Scholar] [CrossRef] [PubMed]
- Vivian, J.P.; Beddoe, T.; McAlister, A.D.; Wilce, M.C.; Zaker-Tabrizi, L.; Troy, S.; Byres, E.; Hoke, D.E.; Cullen, P.A.; Lo, M.; et al. Crystal structure of LipL32, the most abundant surface protein of pathogenic Leptospira spp. J. Mol. Biol. 2009, 387, 1229–1238. [Google Scholar] [CrossRef] [PubMed]
- Tung, J.Y.; Yang, C.W.; Chou, S.W.; Lin, C.C.; Sun, Y.J. Calcium binds to LipL32, a lipoprotein from pathogenic Leptospira, and modulates fibronectin binding. J. Biol. Chem. 2010, 285, 3245–3252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hauk, P.; Macedo, F.; Romero, E.C.; Vasconcellos, S.A.; de Morais, Z.M.; Barbosa, A.S.; Ho, P.L. In LipL32, the major leptospiral lipoprotein, the C terminus is the primary immunogenic domain and mediates interaction with collagen IV and plasma fibronectin. Infect. Immun. 2008, 76, 2642–2650. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoke, D.E.; Egan, S.; Cullen, P.A.; Adler, B. LipL32 is an extracellular matrix-interacting protein of Leptospira spp. and Pseudoalteromonas tunicata. Infect. Immun. 2008, 76, 2063–2069. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vieira, M.L.; Fernandes, L.G.; Domingos, R.F.; Oliveira, R.; Siqueira, G.H.; Souza, N.M.; Teixeira, A.R.; Atzingen, M.V.; Nascimento, A.L. Leptospiral extracellular matrix adhesins as mediators of pathogen-host interactions. FEMS Microbiol. Lett. 2014, 352, 129–139. [Google Scholar] [CrossRef] [Green Version]
- Sun, Z.; Bao, L.; Li, D.; Huang, B.; Wu, B. Effect of Leptospira interrogans outer membrane proteins LipL32 on HUVEC. Microb. Pathog. 2010, 49, 116–121. [Google Scholar] [CrossRef]
- Tian, Y.C.; Chen, Y.C.; Hung, C.C.; Chang, C.T.; Wu, M.S.; Phillips, A.O.; Yang, C.W. Leptospiral outer membrane protein induces extracellular matrix accumulation through a TGF-beta1/Smad-dependent pathway. J. Am. Soc. Nephrol. JASN 2006, 17, 2792–2798. [Google Scholar] [CrossRef] [Green Version]
- Hsu, S.H.; Hung, C.C.; Chang, M.Y.; Ko, Y.C.; Yang, H.Y.; Hsu, H.H.; Tian, Y.C.; Chou, L.F.; Pan, R.L.; Tseng, F.G.; et al. Active components of Leptospira outer membrane protein LipL32 to Toll-like receptor 2. Sci. Rep. 2017, 7, 8363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cullen, P.A.; Haake, D.A.; Bulach, D.M.; Zuerner, R.L.; Adler, B. LipL21 is a novel surface-exposed lipoprotein of pathogenic Leptospira species. Infect. Immun. 2003, 71, 2414–2421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cullen, P.A.; Xu, X.; Matsunaga, J.; Sanchez, Y.; Ko, A.I.; Haake, D.A.; Adler, B. Surfaceome of Leptospira spp. Infect. Immun. 2005, 73, 4853–4863. [Google Scholar] [CrossRef] [Green Version]
- Seenichamy, A.; Bahaman, A.R.; Mutalib, A.R.; Khairani-Bejo, S. Production and characterization of a polyclonal antibody of anti-rLipL21-IgG against leptospira for early detection of acute leptospirosis. BioMed Res. Int. 2014, 2014, 592858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mb, T.; Af, T.; Alto, N. The leptospiral LipL21 and LipL41 proteins exhibit a broad spectrum of interactions with host cell components. Virulence 2021, 12, 2798–2813. [Google Scholar] [CrossRef] [PubMed]
- Vieira, M.L.; Teixeira, A.F.; Pidde, G.; Ching, A.T.C.; Tambourgi, D.V.; Nascimento, A.; Herwald, H. Leptospira interrogans outer membrane protein LipL21 is a potent inhibitor of neutrophil myeloperoxidase. Virulence 2018, 9, 414–425. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arnhold, J.; Flemmig, J. Human myeloperoxidase in innate and acquired immunity. Arch. Biochem. Biophys. 2010, 500, 92–106. [Google Scholar] [CrossRef]
- Pinne, M.; Haake, D.A. A comprehensive approach to identification of surface-exposed, outer membrane-spanning proteins of Leptospira interrogans. PLoS ONE 2009, 4, e6071. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Viratyosin, W.; Ingsriswang, S.; Pacharawongsakda, E.; Palittapongarnpim, P. Genome-wide subcellular localization of putative outer membrane and extracellular proteins in Leptospira interrogans serovar Lai genome using bioinformatics approaches. BMC Genom. 2008, 9, 181. [Google Scholar] [CrossRef] [Green Version]
- Hartwig, D.D.; Seixas, F.K.; Cerqueira, G.M.; McBride, A.J.; Dellagostin, O.A. Characterization of the immunogenic and antigenic potential of putative lipoproteins from Leptospira interrogans. Curr. Microbiol. 2011, 62, 1337–1341. [Google Scholar] [CrossRef]
- Koizumi, N.; Watanabe, H. Molecular cloning and characterization of a novel leptospiral lipoprotein with OmpA domain. FEMS Microbiol. Lett. 2003, 226, 215–219. [Google Scholar] [CrossRef] [Green Version]
- Hsu, S.H.; Chang, M.Y.; Lin, S.M.; Ko, Y.C.; Chou, L.F.; Tian, Y.C.; Hung, C.C.; Yang, C.W. Peptidoglycan mediates Leptospira outer membrane protein Loa22 to toll-like receptor 2 for inflammatory interaction: A novel innate immune recognition. Sci. Rep. 2021, 11, 1064. [Google Scholar] [CrossRef] [PubMed]
- Juncker, A.S.; Willenbrock, H.; Von Heijne, G.; Brunak, S.; Nielsen, H.; Krogh, A. Prediction of lipoprotein signal peptides in Gram-negative bacteria. Protein Sci. A Publ. Protein Soc. 2003, 12, 1652–1662. [Google Scholar] [CrossRef]
- Shin, S.; Lu, G.; Cai, M.; Kim, K.S. Escherichia coli outer membrane protein A adheres to human brain microvascular endothelial cells. Biochem. Biophys. Res. Commun. 2005, 330, 1199–1204. [Google Scholar] [CrossRef]
- Koebnik, R. Proposal for a peptidoglycan-associating alpha-helical motif in the C-terminal regions of some bacterial cell-surface proteins. Mol. Microbiol. 1995, 16, 1269–1270. [Google Scholar] [CrossRef] [PubMed]
- Grizot, S.; Buchanan, S.K. Structure of the OmpA-like domain of RmpM from Neisseria meningitidis. Mol. Microbiol. 2004, 51, 1027–1037. [Google Scholar] [CrossRef]
- Singh, S.P.; Williams, Y.U.; Miller, S.; Nikaido, H. The C-terminal domain of Salmonella enterica serovar typhimurium OmpA is an immunodominant antigen in mice but appears to be only partially exposed on the bacterial cell surface. Infect. Immun. 2003, 71, 3937–3946. [Google Scholar] [CrossRef] [Green Version]
- Pautsch, A.; Schulz, G.E. Structure of the outer membrane protein A transmembrane domain. Nat. Struct. Biol. 1998, 5, 1013–1017. [Google Scholar] [CrossRef]
- Torres, A.G.; Jeter, C.; Langley, W.; Matthysse, A.G. Differential binding of Escherichia coli O157:H7 to alfalfa, human epithelial cells, and plastic is mediated by a variety of surface structures. Appl. Environ. Microbiol. 2005, 71, 8008–8015. [Google Scholar] [CrossRef] [Green Version]
- Jeannin, P.; Bottazzi, B.; Sironi, M.; Doni, A.; Rusnati, M.; Presta, M.; Maina, V.; Magistrelli, G.; Haeuw, J.F.; Hoeffel, G.; et al. Complexity and complementarity of outer membrane protein A recognition by cellular and humoral innate immunity receptors. Immunity 2005, 22, 551–560. [Google Scholar] [CrossRef] [Green Version]
- Haake, D.A.; Matsunaga, J. Characterization of the leptospiral outer membrane and description of three novel leptospiral membrane proteins. Infect. Immun. 2002, 70, 4936–4945. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barbosa, A.S.; Abreu, P.A.E.; Neves, F.O.; Atzingen, M.V.; Watanabe, M.M.; Vieira, M.L.; Morais, Z.M.; Vasconcellos, S.A.; Nascimento, A.L.T.O. Newly identified leptospiral adhesin mediates attachment to laminin. Infect. Immun. 2006, 74, 6356–6364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fouts, D.E.; Matthias, M.A.; Adhikarla, H.; Adler, B.; Amorim-Santos, L.; Berg, D.E.; Bulach, D.; Buschiazzo, A.; Chang, Y.F.; Galloway, R.L.; et al. What makes a bacterial species pathogenic?: Comparative genomic analysis of the genus Leptospira. PLoS Negl. Trop. Dis. 2016, 10, e0004403. [Google Scholar] [CrossRef] [Green Version]
- Kobe, B.; Kajava, A.V. The leucine-rich repeat as a protein recognition motif. Curr. Opin. Struct. Biol. 2001, 11, 725–732. [Google Scholar] [CrossRef]
- Soundararajan, V.; Patel, N.; Subramanian, V.; Sasisekharan, V.; Sasisekharan, R. The many faces of the YopM effector from plague causative bacterium Yersinia pestis and its implications for host immune modulation. Innate Immun. 2011, 17, 548–557. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loimaranta, V.; Hytonen, J.; Pulliainen, A.T.; Sharma, A.; Tenovuo, J.; Stromberg, N.; Finne, J. Leucine-rich repeats of bacterial surface proteins serve as common pattern recognition motifs of human scavenger receptor gp340. J. Biol. Chem. 2009, 284, 18614–18623. [Google Scholar] [CrossRef] [Green Version]
- Seepersaud, R.; Hanniffy, S.B.; Mayne, P.; Sizer, P.; Le Page, R.; Wells, J.M. Characterization of a novel leucine-rich repeat protein antigen from group B streptococci that elicits protective immunity. Infect. Immun. 2005, 73, 1671–1683. [Google Scholar] [CrossRef] [Green Version]
- Schubert, W.D.; Urbanke, C.; Ziehm, T.; Beier, V.; Machner, M.P.; Domann, E.; Wehland, J.; Chakraborty, T.; Heinz, D.W. Structure of internalin, a major invasion protein of Listeria monocytogenes, in complex with its human receptor E-cadherin. Cell 2002, 111, 825–836. [Google Scholar] [CrossRef] [Green Version]
- Bernal-Bayard, J.; Cardenal-Munoz, E.; Ramos-Morales, F. The Salmonella type III secretion effector, salmonella leucine-rich repeat protein (SlrP), targets the human chaperone ERdj3. J. Biol. Chem. 2010, 285, 16360–16368. [Google Scholar] [CrossRef] [Green Version]
- Marino, M.; Braun, L.; Cossart, P.; Ghosh, P. Structure of the lnlB leucine-rich repeats, a domain that triggers host cell invasion by the bacterial pathogen L. monocytogenes. Mol. Cell 1999, 4, 1063–1072. [Google Scholar] [CrossRef]
- Liu, C.I.; Cheng, T.L.; Chen, S.Z.; Huang, Y.C.; Chang, W.T. LrrA, a novel leucine-rich repeat protein involved in cytoskeleton remodeling, is required for multicellular morphogenesis in Dictyostelium discoideum. Dev. Biol. 2005, 285, 238–251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ikegami, A.; Honma, K.; Sharma, A.; Kuramitsu, H.K. Multiple functions of the leucine-rich repeat protein LrrA of Treponema denticola. Infect. Immun. 2004, 72, 4619–4627. [Google Scholar] [CrossRef] [Green Version]
- Miras, I.; Saul, F.; Nowakowski, M.; Weber, P.; Haouz, A.; Shepard, W.; Picardeau, M. Structural characterization of a novel subfamily of leucine-rich repeat proteins from the human pathogen Leptospira interrogans. Acta Crystallogr. Sect. D Biol. Crystallogr. 2015, 71, 1351–1359. [Google Scholar] [CrossRef] [PubMed]
- Doxey, A.C.; McConkey, B.J. Prediction of molecular mimicry candidates in human pathogenic bacteria. Virulence 2013, 4, 453–466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bella, J.; Hindle, K.L.; McEwan, P.A.; Lovell, S.C. The leucine-rich repeat structure. Cell. Mol. Life Sci. CMLS 2008, 65, 2307–2333. [Google Scholar] [CrossRef] [PubMed]
- Nitipan, S.; Sritrakul, T.; Kunjantarachot, A.; Prapong, S. Identification of epitopes in Leptospira borgpetersenii leucine-rich repeat proteins. Infect. Genet. Evol. J. Mol. Epidemiol. Evol. Genet. Infect. Dis. 2013, 14, 46–57. [Google Scholar] [CrossRef] [PubMed]
- Shevchenko, D.V.; Akins, D.R.; Robinson, E.; Li, M.; Popova, T.G.; Cox, D.L.; Radolf, J.D. Molecular characterization and cellular localization of TpLRR, a processed leucine-rich repeat protein of Treponema pallidum, the syphilis spirochete. J. Bacteriol. 1997, 179, 3188–3195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, A.; Sojar, H.T.; Glurich, I.; Honma, K.; Kuramitsu, H.K.; Genco, R.J. Cloning, expression, and sequencing of a cell surface antigen containing a leucine-rich repeat motif from Bacteroides forsythus ATCC 43037. Infect. Immun. 1998, 66, 5703–5710. [Google Scholar] [CrossRef] [Green Version]
- Sanchez-Beato, A.R.; Lopez, R.; Garcia, J.L. Molecular characterization of PcpA: A novel choline-binding protein of Streptococcus pneumoniae. FEMS Microbiol. Lett. 1998, 164, 207–214. [Google Scholar] [CrossRef] [Green Version]
- Chou, L.F.; Chen, T.W.; Ko, Y.C.; Pan, M.J.; Tian, Y.C.; Chiu, C.H.; Tang, P.; Hung, C.C.; Yang, C.W. Potential impact on kidney infection: A whole-genome analysis of Leptospira santarosai serovar Shermani. Emerg Microbes Infect. 2014, 3, e82. [Google Scholar] [CrossRef]
- Hsu, S.H.; Chu, C.H.; Tian, Y.C.; Chang, M.Y.; Chou, L.F.; Sun, Y.J.; Yang, C.W. Crystal structure of Leptospira leucine-rich repeat 20 reveals a novel E-cadherin binding protein to induce NGAL expression in HK2 cells. Biochem. J. 2020, 477, 4313–4326. [Google Scholar] [CrossRef] [PubMed]
- Hsu, S.H.; Chou, L.F.; Hong, C.H.; Chang, M.Y.; Tsai, C.Y.; Tian, Y.C.; Yang, H.Y.; Yang, C.W. Crosstalk between E-cadherin/beta-catenin and NF-kappaB signaling pathways: The regulation of host-pathogen interaction during leptospirosis. Int. J. Mol. Sci. 2021, 22, 13132. [Google Scholar] [CrossRef] [PubMed]
- Verma, A.; Artiushin, S.; Matsunaga, J.; Haake, D.A.; Timoney, J.F. LruA and LruB, novel lipoproteins of pathogenic Leptospira interrogans associated with equine recurrent uveitis. Infect. Immun. 2005, 73, 7259–7266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verma, A.; Kumar, P.; Babb, K.; Timoney, J.F.; Stevenson, B. Cross-reactivity of antibodies against leptospiral recurrent uveitis-associated proteins A and B (LruA and LruB) with eye proteins. PLoS Negl. Trop. Dis. 2010, 4, e778. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palaniappan, R.U.; McDonough, S.P.; Divers, T.J.; Chen, C.S.; Pan, M.J.; Matsumoto, M.; Chang, Y.F. Immunoprotection of recombinant leptospiral immunoglobulin-like protein A against Leptospira interrogans serovar Pomona infection. Infect. Immun. 2006, 74, 1745–1750. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choy, H.A.; Kelley, M.M.; Chen, T.L.; Moller, A.K.; Matsunaga, J.; Haake, D.A. Physiological osmotic induction of Leptospira interrogans adhesion: LigA and LigB bind extracellular matrix proteins and fibrinogen. Infect. Immun. 2007, 75, 2441–2450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, Y.P.; Chang, Y.F. A domain of the Leptospira LigB contributes to high affinity binding of fibronectin. Biochem. Biophys. Res. Commun. 2007, 362, 443–448. [Google Scholar] [CrossRef]
- Lin, M.H.; Chang, Y.C.; Hsiao, C.D.; Huang, S.H.; Wang, M.S.; Ko, Y.C.; Yang, C.W.; Sun, Y.J. LipL41, a hemin binding protein from Leptospira santarosai serovar Shermani. PLoS ONE 2013, 8, e83246. [Google Scholar] [CrossRef] [Green Version]
- Gautam, R.; Guptill, L.F.; Wu, C.C.; Potter, A.; Moore, G.E. Spatial and spatio-temporal clustering of overall and serovar-specific Leptospira microscopic agglutination test (MAT) seropositivity among dogs in the United States from 2000 through 2007. Prev Vet Med 2010, 96, 122–131. [Google Scholar] [CrossRef]
- Levett, P.N. Usefulness of serologic analysis as a predictor of the Infecting serovar in patients with severe leptospirosis. Clin. Infect. Dis. 2003, 36, 447–452. [Google Scholar] [CrossRef]
- Smythe, L.D.; Wuthiekanun, V.; Chierakul, W.; Suputtamongkol, Y.; Tiengrim, S.; Dohnt, M.F.; Symonds, M.L.; Slack, A.T.; Apiwattanaporn, A.; Chueasuwanchai, S.; et al. The microscopic agglutination test (MAT) is an unreliable predictor of infecting Leptospira serovar in Thailand. Am. J. Trop. Med. Hyg. 2009, 81, 695–697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohd Ali, M.R.; Sum, J.S.; Aminuddin Baki, N.N.; Choong, Y.S.; Nor Amdan, N.A.; Amran, F.; Lim, T.S. Development of monoclonal antibodies against recombinant LipL21 protein of pathogenic Leptospira through phage display technology. Int. J. Biol. Macromol 2021, 168, 289–300. [Google Scholar] [CrossRef] [PubMed]
- Goris, M.G.; Boer, K.R.; Duarte, T.A.; Kliffen, S.J.; Hartskeerl, R.A. Human leptospirosis trends, The Netherlands, 1925–2008. Emerg Infect. Dis. 2013, 19, 371–378. [Google Scholar] [CrossRef] [PubMed]
- Mohd Ali, M.R.; Mohd Safee, A.W.; Ismail, N.H.; Abu Sapian, R.; Mat Hussin, H.; Ismail, N.; Yean Yean, C. Development and validation of pan-Leptospira Taqman qPCR for the detection of Leptospira spp. in clinical specimens. Mol. Cell Probes 2018, 38, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Miotto, B.A.; da Hora, A.S.; Taniwaki, S.A.; Brandao, P.E.; Heinemann, M.B.; Hagiwara, M.K. Development and validation of a modified TaqMan based real-time PCR assay targeting the lipl32 gene for detection of pathogenic Leptospira in canine urine samples. Braz. J. Microbiol. 2018, 49, 584–590. [Google Scholar] [CrossRef] [PubMed]
- Esteves, L.M.; Bulhoes, S.M.; Branco, C.C.; Carreira, T.; Vieira, M.L.; Gomes-Solecki, M.; Mota-Vieira, L. Diagnosis of human leptospirosis in a clinical setting: Real-time PCR high resolution melting analysis for detection of Leptospira at the onset of disease. Sci. Rep. 2018, 8, 9213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chirathaworn, C.; Chantaramalai, T.; Sereemaspun, A.; Kongthong, N.; Suwancharoen, D. Detection of Leptospira in urine using anti-Leptospira-coated gold nanoparticles. Comp. Immunol. Microbiol. Infect. Dis. 2011, 34, 31–34. [Google Scholar] [CrossRef]
- Saengjaruk, P.; Chaicumpa, W.; Watt, G.; Bunyaraksyotin, G.; Wuthiekanun, V.; Tapchaisri, P.; Sittinont, C.; Panaphut, T.; Tomanakan, K.; Sakolvaree, Y.; et al. Diagnosis of human leptospirosis by monoclonal antibody-based antigen detection in urine. J. Clin. Microbiol. 2002, 40, 480–489. [Google Scholar] [CrossRef]
- Fernandes, C.P.; Seixas, F.K.; Coutinho, M.L.; Vasconcellos, F.A.; Seyffert, N.; Croda, J.; McBride, A.J.; Ko, A.I.; Dellagostin, O.A.; Aleixo, J.A. Monoclonal antibodies against LipL32, the major outer membrane protein of pathogenic Leptospira: Production, characterization, and testing in diagnostic applications. Hybridoma 2007, 26, 35–41. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hsu, S.-H.; Yang, C.-W. Insight into the Structure, Functions, and Dynamics of the Leptospira Outer Membrane Proteins with the Pathogenicity. Membranes 2022, 12, 300. https://doi.org/10.3390/membranes12030300
Hsu S-H, Yang C-W. Insight into the Structure, Functions, and Dynamics of the Leptospira Outer Membrane Proteins with the Pathogenicity. Membranes. 2022; 12(3):300. https://doi.org/10.3390/membranes12030300
Chicago/Turabian StyleHsu, Shen-Hsing, and Chih-Wei Yang. 2022. "Insight into the Structure, Functions, and Dynamics of the Leptospira Outer Membrane Proteins with the Pathogenicity" Membranes 12, no. 3: 300. https://doi.org/10.3390/membranes12030300
APA StyleHsu, S. -H., & Yang, C. -W. (2022). Insight into the Structure, Functions, and Dynamics of the Leptospira Outer Membrane Proteins with the Pathogenicity. Membranes, 12(3), 300. https://doi.org/10.3390/membranes12030300