1. Introduction
The electric properties of biomembranes play a pivotal role in cellular functions. The parameters of electrical equivalent circuits of biological and model bilayer membranes—capacitance and conductance and their dependence on external influences—have been intensively studied over the past 50 years. The dependence of the capacitance of lipid bilayer membranes with various solvents on the electric field was studied in early experiments [
1,
2,
3,
4,
5,
6]. The increase in capacitance
C under the action of an electric field was described by the empirical formula
. The coefficient of the nonlinearity of capacitance β for membranes with a solvent was estimated at 2–10 V
−2, for membranes without a solvent (“dry”) membranes 0.022 V
−2 [
1]. In studies of the nonlinear capacitance of the droplet interface bilayer (DIB), the value of the coefficient of nonlinearity depended on the type of solvent and was 25.8 V
−2 for membranes of diphytanoylphosphatidylcholine (DPhPC) in decane and 1.75 V
−2 in hexadecane [
7,
8]. The value of the coefficient for cells was estimated to be 0.108–0.135 V
−2 [
9]. The change in the capacitance of membranes under the action of an electric field is associated with various processes—electrostriction, a change in the thickness of the membrane as a result of solvent exclusion, membrane area increase [
1,
2,
3,
4,
5,
6], and thermal fluctuations [
10]. These processes have different relaxation times and, under the action of periodic electric fields, should manifest themselves in different frequency ranges. The dynamic current response of the membrane and its equivalent parameters must depend on the frequency of the alternating voltage. The nonlinearity coefficient β is expected to be maximum at low frequencies, since in this case all the processes occurring under the action of the electric field and responsible for the nonlinearity have time to take place. Inelastic (dissipative) effects of various natures, causing a delay in the polarization of the membrane and capable of causing hysteresis phenomena, should also manifest themselves in different frequency ranges.
Hysteresis effects on the current responses of the membrane to a periodic triangular voltage were noted in early experiments [
5,
6]. These phenomena are also discussed in works where DIB is used as a biological membrane model and dynamic capacitive currents in bilayer membranes with different solvents are studied [
7,
8,
11]. It was shown in [
11] that the existence of hysteresis is associated with voltage-dependent changes in the geometry of the bilayer, which depend on the viscosity of the solvent (decane and hexadecane were compared), temperature and the frequency of the command sinusoidal voltage. The authors first drew attention to a new aspect of studying the dynamic electrical characteristics of bilayer lipid membranes, showing that lipid DIB membranes with a solvent under the action of a periodic sinusoidal voltage exhibit the properties of memcapacitive (memory capacitive) systems. The concept of memristic systems proposed in [
12] was extended to capacitive elements—capacitors, the properties of which depend on the state and history of the system. A change in the capacitance of such systems under the action of an external control voltage can be associated with geometric changes in the systems and the influence of inelastic (dissipative) effects [
13]. Memcapacitors exhibit narrowed hysteresis loops in charge–voltage dependence. It is assumed that these systems can be used as synapses in artificial neural networks [
14,
15,
16,
17]. DIB membranes showing pinched hysteresis loops in charge–voltage coordinates, as well as in experimental capacitance–voltage dependences [
11], are almost the only example of the physical implementation of memcapacitance.
To evaluate the parameters characterizing the dynamic electrical properties of the model and biological membranes, periodic command voltage of various types is applied to them in voltage clamp mode: triangular or sinusoidal voltage, rectangular pulses, voltage ramp stimulus, or combinations thereof [
18]. Interpretation of experimental responses to these signals depends on the choice of an adequate equivalent circuit that allows one to estimate the characteristics of the membrane with varying accuracy. Traditionally, a membrane is thought of as a parallel connection between a capacitor and a resistor. Series resistance (access resistance) in the equivalent circuit of the membrane allows for a more accurate estimation of membrane parameters [
19,
20]. However, there are few models that describe the dynamic electrical behavior of membranes with non-linear capacitances. Basically, for the theoretical description of nonlinear and transient processes in the current response of the membrane, the capacitances of the classical equivalent circuit are assumed to be exponentially dependent on time in an explicit form [
21,
22] or described by first-order differential equations [
10,
11].
In this paper, we introduce a mathematical basis for interpreting experimental observations of the BLM input-output dynamics as memcapacitors with external control of a triangular-shaped periodic signal. For an analytical description of memcapacitor properties, we propose an equivalent circuit of a membrane in the form of a parallel connection of a viscoelastic capacitor [
23] and resistance modeling of the ion permeability of membranes. The ion permeability can increase in the presence of protein or lipid channels and affect the memcapacitance properties. Compiling a nonlinear differential equation for the equivalent circuit and solving it using the small parameter method, we obtain explicit analytical dependences of the current response of the membrane and pinched-hysteresis loops on time and applied voltage.
A comparison of analytical expressions with experimental current recordings of membranes confirms the adequacy of the equivalent circuit and the mathematical model. Explicit solutions for the current–voltage characteristics and pinched-hysteresis loops in comparison with the experimental data make it possible to estimate the parameters of the equivalent circuit and their dependence on the characteristics of the input signal and the properties of the membrane. The resulting formulas allow us to determine how these parameters control the memcapacitance behavior of the membrane and the amount of hysteresis in the capacitance–voltage coordinates.
We also quantify pinched hysteresis in terms of the work done by the control voltage, for which we obtain explicit formulas in terms of equivalent circuit parameters and control signal characteristics. Interpretation of the properties of a lipid bilayer membrane in the context of memcapacitance systems allows us to define a lipid bilayer membrane as an active memcapacitive device. The nonlinearity of a capacitance of a certain type, combined with viscous dissipative processes that cause a delay in membrane polarization, leads to the appearance of an insertion negative conductance proportional to the width of the hysteresis loop.
2. Materials and Methods
2.1. Electrical Model of a Viscoelastic Membrane: Solving a Nonlinear Differential Equation
The main processes leading to a noticeable non-linear change in the capacitance of a planar bilayer lipid membrane under an electric field are deformation of the membrane and a decrease in its thickness due to solvent extrusion, as well as an increase in the area of the membrane. These processes proceed quite slowly (relaxation times of solvent extrusion are proportional to its viscosity and amount to 5 ms–5 s [
5,
24,
25,
26]. When a cyclic electric field of corresponding frequencies is applied, these processes affect the dynamic behavior of the geometric dimensions of the membrane and, accordingly, its equivalent electrical parameters.
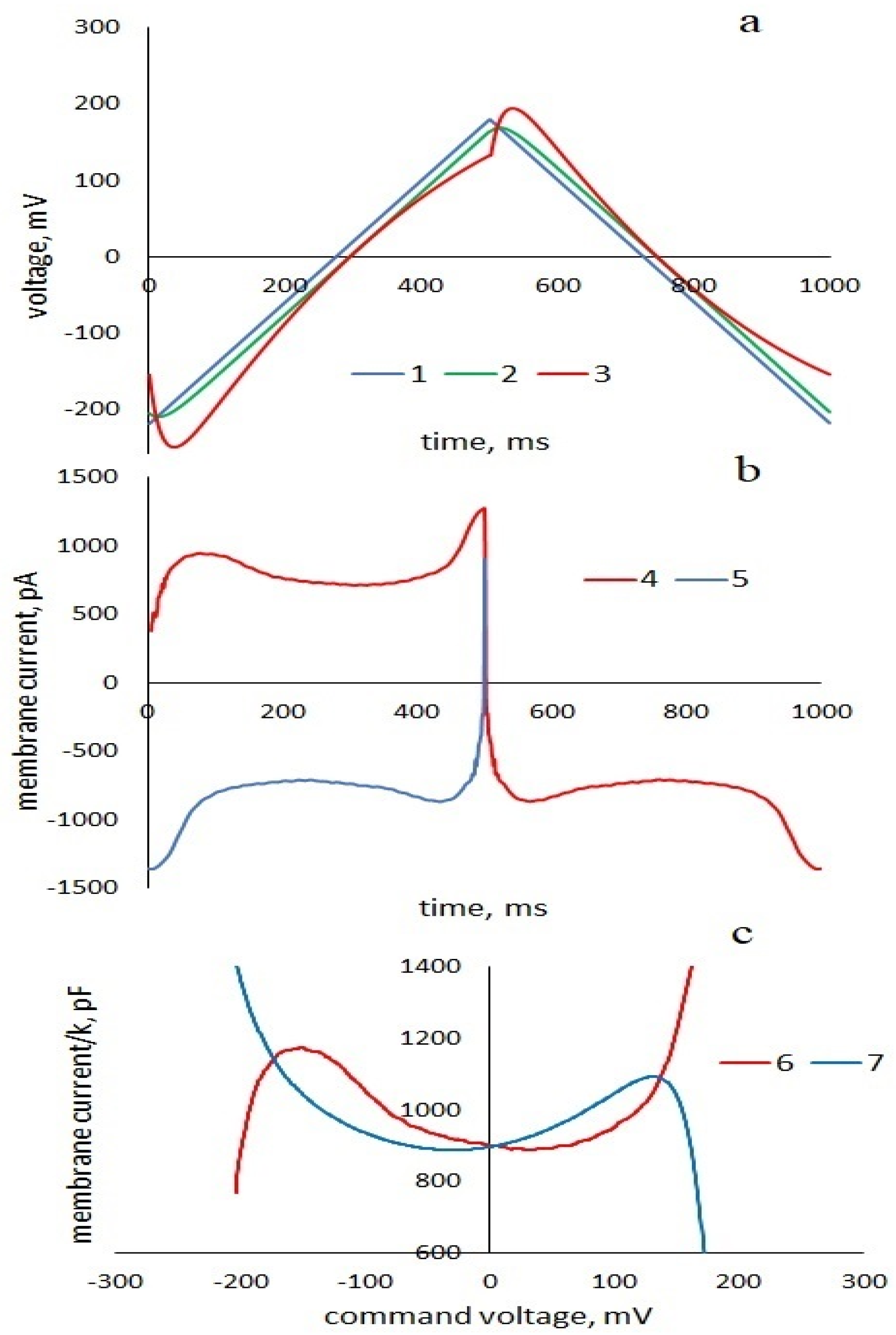
To analytically describe the dynamic current response of the membrane (
Figure 1) to a periodic triangular voltage with frequencies corresponding to the relaxation times, we consider the membrane as a viscoelastic capacitor [
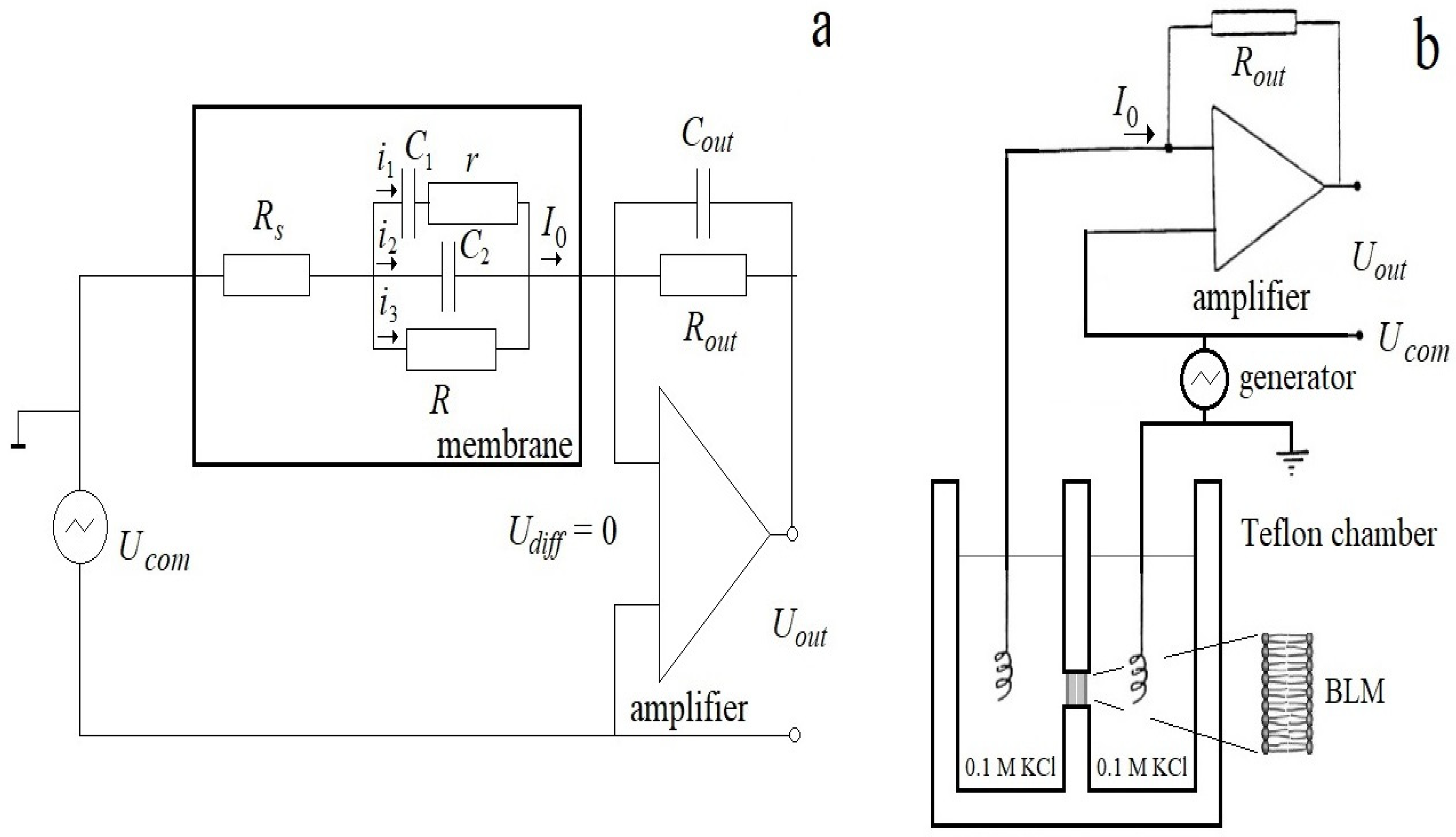
23]. Following the logic of this work, let us expand the total current response of the membrane into that caused by a purely elastic change in geometry and a non-linear current that depends on the prehistory of the process. The circuit of this current imitates the viscoelastic response of a section of the membrane in which the electrical displacement and polarization lag behind the field strength and depend on its value at the previous time. This occurs when the electric polarization does not have time to follow the changes in the electric field. The equivalent circuit of the membrane is shown in
Figure 2. The equivalent parameters of the membrane are the series connection of resistance r, proportional to the viscosity of the solvent, and the nonlinear capacitance
The material parameter
represents physically the fraction of the membrane, which in a given time range has time-independent deformation (solvent),
is the coefficient of capacitance nonlinearity for this part of the membrane,
is the capacitance of the entire membrane at zero voltage,
is the voltage across the capacitance
, physically proportional to the electrical displacement. Thus, the current circuit
represents a part of the membrane; the deformation and the corresponding change in capacitance are time-dependent processes due to viscous processes that prevent changes in the membrane geometry. The corresponding equation for the current is given by:
The capacitance in the second branch
is the capacitance of the part of the membrane that gives a purely elastic response, the nonlinearity of which can be neglected, since the coefficient of nonlinearity of “dry” membranes without solvent
[
1]. The membrane is purely elastic when ϰ = 1. The current in this branch is
where
U is the command voltage proportional to the field strength across the membrane.
The third branch represents ionic currents through the membrane, which arise when the ionic permeability of the membranes increases. In order to better agree with the experimental data, this model provides the possibility of the appearance of nonlinear conductance at high membrane voltages, which was observed in studies of both artificial [
27] and cell membranes [
28]. Following [
28], where the current–voltage profile of the cell was represented by a 9th order polynomial, we represent the ion current through the membrane as
where g is the linear conductance of the membrane, γ is an empirical coefficient of nonlinearity of membrane conductance.
Series resistance
(access resistance) is introduced into the equivalent circuits of the model and biological membranes to account for experimental artifacts of voltage clamp measurements. The effects described by the access resistance are associated with the experimental setup, the properties of the electrodes and electrolyte, as well as the relaxation properties of the membrane itself in response to the applied voltage [
19,
20]. Since here we take into account the relaxation properties of the membrane in the viscoelastic circuit and the resistance of the electrodes and electrolyte is low, for simplicity, we neglect this resistance in our analysis.
Let us write the Kirchhoff equations separately for the upward (up) and downward (down) half-periods of the triangular voltage for the viscoelastic circuit.
The right-hand sides of Equations (4) and (5) represent the command triangular voltage supplied from the generator, are voltages across capacitance of the viscoelastic part of the membrane, causing an elastic response, for two half-cycles of the command voltage, is triangular voltage amplitude, k is sweep ramp. The voltage on the right side is proportional to the field strength in the membrane, voltage proportional to the electrical displacement, which, due to viscous processes, lags behind the field strength.
Substituting (1) into Equations (4) and (5), respectively, we obtain two nonlinear differential equations for
and
, corresponding to the upward and downward the command voltage ramping
where
.
Solving Equations (6) and (7) using the small parameter method (see
Supplementary Material, Equations 3S–14S), we obtain expressions for the voltage across the capacitance C
1 of the viscoelastic branch for the upward and downward half-cycles of the command triangular voltage
Figure 1a shows the command voltage U(t), voltages
and
calculated by
Formulas (S9) and (S10) in the zero linear approximation (β = 0) and nonlinear voltages
and
Without taking into account transient processes, we can write
), which is equivalent to the electrical displacement lagging behind the field strength
D(t)~E(t−τ). Thus, the use of series resistance in the equivalent circuit describes a situation that occurs when the frequencies of electromagnetic fields correspond to the frequencies characteristic for establishing the electric polarization of a substance [
29].
The currents in the viscoelastic circuit
and
can be found from Equations (4) and (5), taking into account (8) and (9).
In order to find formulas for pinched hysteresis loops, we find expression for the time inverted current response of the viscoelastic part of the membrane to the downward half-period of the command voltage (
Figure 1b), substituting
into (11) and given that
:We express the time dependences of currents (10) and (12) as a function of command voltage by substituting . The currents (2), (3) are rewritten for the upward and downward half-periods: , .
Then, the total current response of the membrane to the upward and downward half-period of command voltage:
Total inverted current response to the downward half-period of command voltage
The hysteresis loop equations in the C, U coordinates are given by the expressions:
2.2. Experimental
Lipids and electrolytes. Azolectin and 1,2-diphytanoyl-sn-glycero-3-phosphocholine (Avanti Polar Lipids, Alabaster) were used for the formation of planar BLM. The bulk solution contained 0.1 M KCl. The reagent was of analytical grade.
Planar lipid bilayer membranes. The BLMs were formed according to [
30] over a 0.5 mm
2 circular hole in a 1 mm thick wall of a Teflon chamber at room temperature of 21 ± 1 °C. The wall separated two subchambers, each filled with 2.5 mL of the same electrolyte solution. The membrane-forming solution contained 30 mg of lipids dissolved in 1 mL of n-decane. Before each experiment, the vertical wall of the Teflon chamber was covered with a thin layer of dried membrane-forming solution. Once a small droplet (~0.1 µL) of lipid solution is placed below the hole, a bilayer is formed automatically in ~10 min. The formation of the bilayer was followed by capacitance measurements. To estimate the specific capacitance of the membrane, the area of the membrane formed on the hole was determined using a microscope. The specific capacitances of the studied membranes were in the range of 3–4 nF/mm
2.
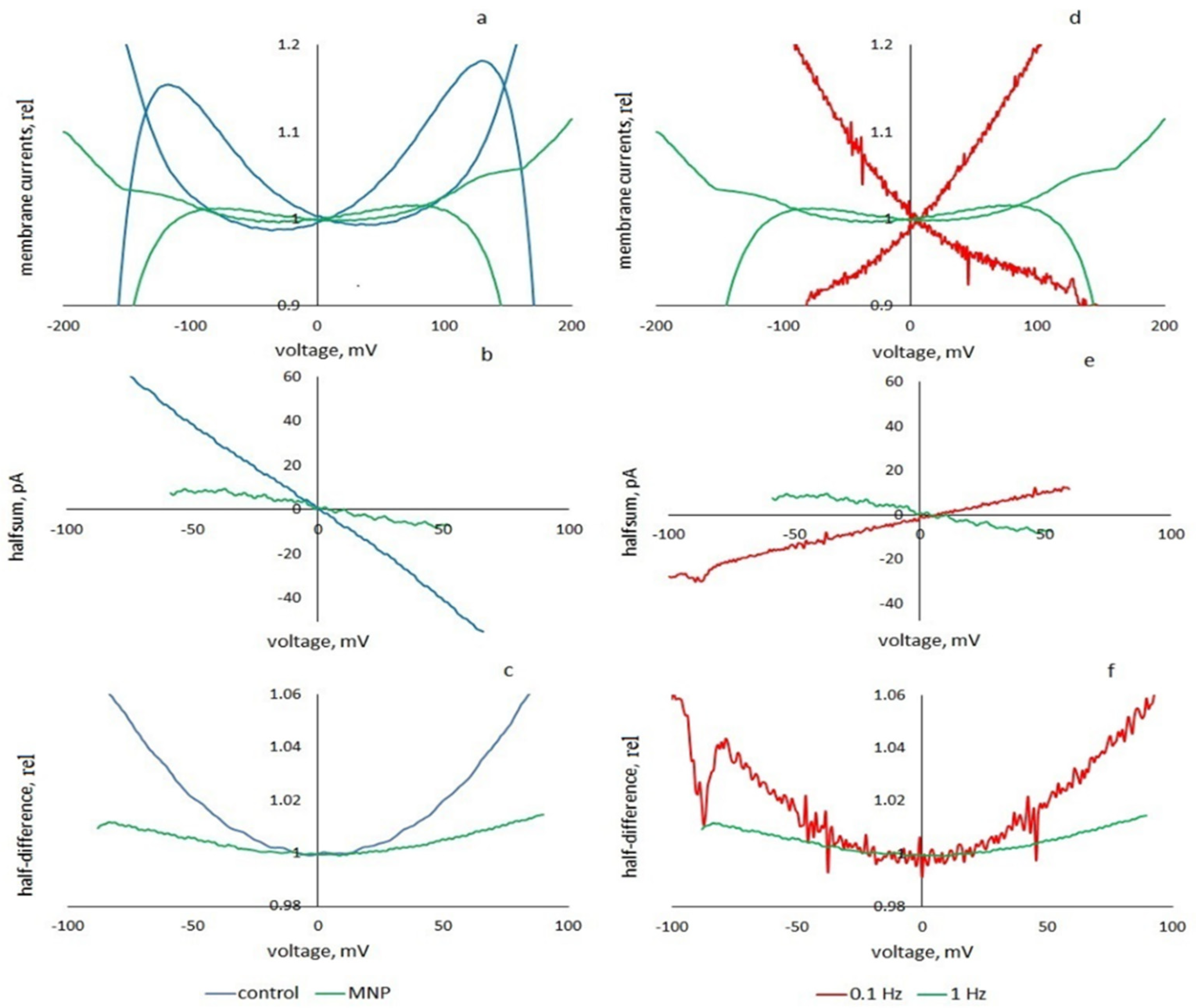
Nanoparticles. To change the membrane stiffness, hydrophobic cubic CoFe2O4 nanoparticles dispersed in toluene were added to the membrane solution at a concentration of 70 μg/mL (synthesized at MISIS, Moscow, Russia). The average diagonal of the nanoparticles was 14 nm.
Electrical measurements. Ag-AgCl STREF1 electrodes (OHAUS Corporation, Parsippany, New Jersey, USA) were placed in both compartments of the chamber. Transmembrane currents were detected with a VA-10X amplifier (NPI Electronics GmbH, Germany) in voltage clamp mode. Currents were recorded with a sampling rate of 1 kHz in a 16-digit ADC (L-Card, Moscow, Russia). The measurements were carried out in the voltage-clamp conditions. An AKIP 3409/1 arbitrary waveform generator (PRIST, Russia) was used to apply a triangular voltage to the membrane.
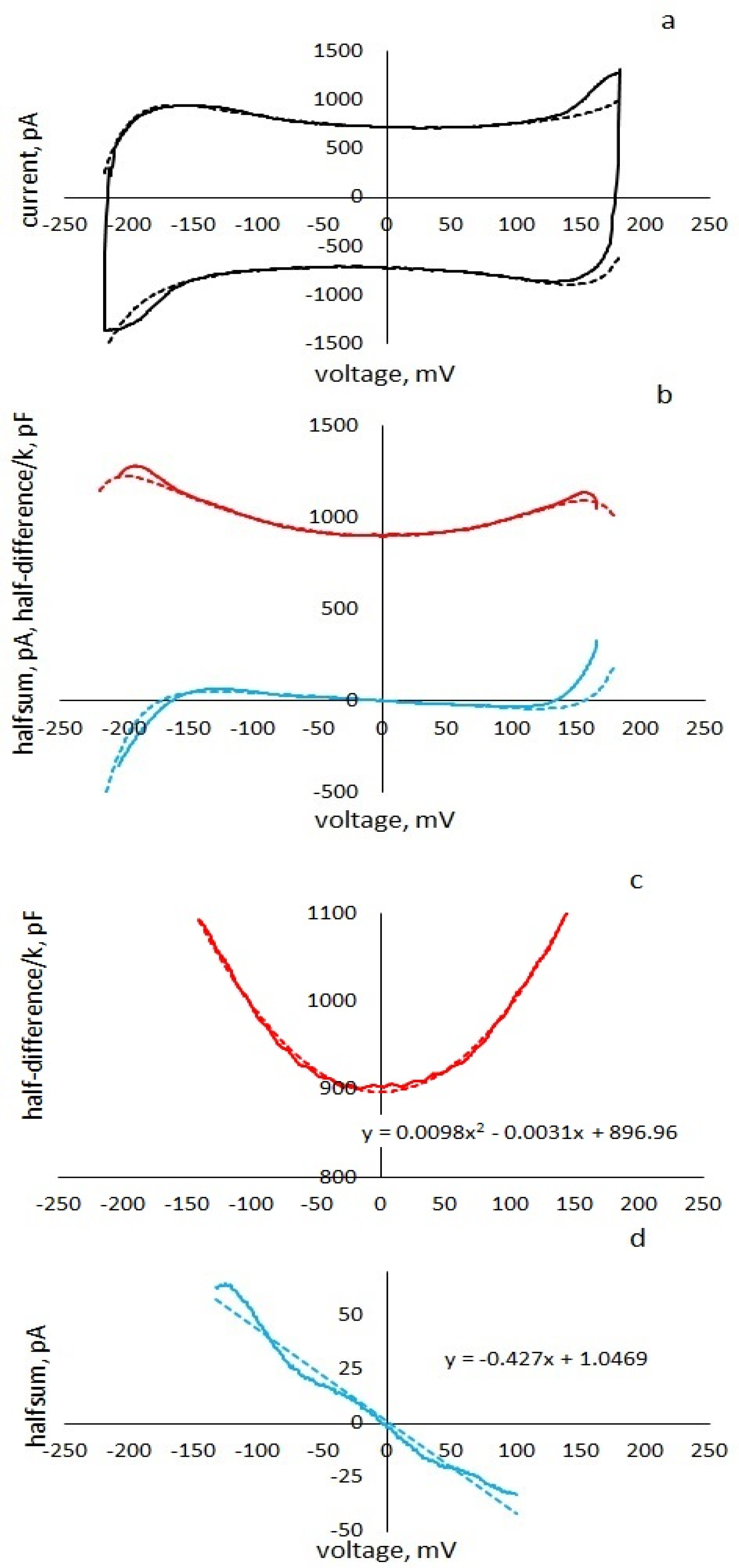
Description of the experiment. A periodic triangular voltage of various frequencies and amplitudes was applied to the membrane in voltage clamping mode, and the current responses of the membrane were recorded. Separately, the records of the current were distinguished in response to the upward and downward half-periods of the command voltage. The current responses to downward voltage ramping were inverted over time (
Figure 1b). The data were presented in the form
. The experimental pinched-hysteresis loops were plotted using the curves
and
in absolute or relative capacitance units.
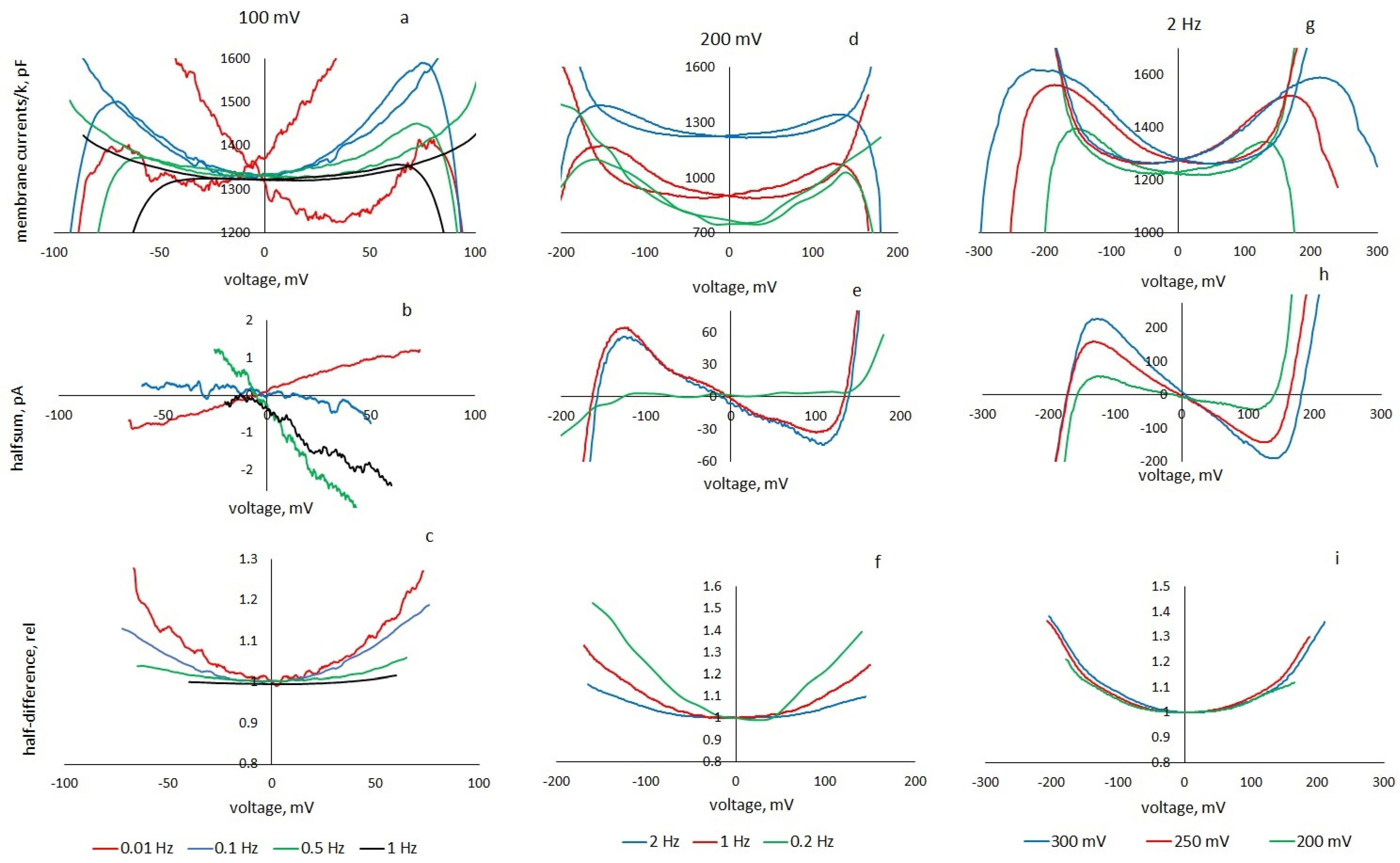
The measurements were carried out as follows: a triangular voltage with amplitudes of 50, 100, 130, 150, 175, 200, 225, 250, 275 and 300 mV was applied to the formed membranes; measurements were carried out at frequencies of 0.01, 0.1, 0.2, 0.35, 0.5, 0.66, 0.73, 0.8, 0.88, 1, 1.33, 1.5 and 2 Hz. For each amplitude of the triangular voltage, a current response was recorded with a duration of 20–30 periods. In each fragment, the current responses to the upward and downward half-periods of the command voltage were distinguished. To reduce the level of fluctuations at low frequencies, the signal was averaged over 30 ms. At low frequencies (<0.35 Hz), the membranes, as a rule, could not withstand high voltage; therefore, the results of measurements at the maximum amplitude before the membrane rupture were processed.
At a frequency of 0.01 Hz, two membranes were measured (triangular voltage amplitude 100 mV), at a frequency of 0.1 Hz- 2 membranes (100 mV), 1 membrane (150 mV) and 1 membrane (200 mV in the presence of particles). At a frequency of 0.2 Hz, 1 membrane was measured at amplitudes up to 200 mV, 1 membrane (150 mV) and 2 membranes (100 mV), at a frequency of 0.35 Hz—2 membranes (150 and 175 mV). At a frequency of 0.5 Hz, 4 membranes were measured at 130 mV and 2 membranes at 150 mV. A membrane was studied for which the rate of the membrane voltage sweep k = 0.8 V/s was the same for different frequencies and voltages: 0.66 Hz and 300 mV, 0.73 Hz and 275 mV, 0.8 Hz and 250 mV, 0.88 Hz and 225 mV, 1.33 Hz and 150 mV. At a frequency of 1 Hz, more than 20 membranes at 50 mV and 100 mV, 4 membranes at 150 mV and 14 membranes at 200 mV were recorded and processed. The current responses to a triangular voltage with an amplitude of 50 mV showed no nonlinearity or hysteresis, but we used them for transient analysis (in these cases, the signal was not averaged). At a frequency of 1.5 Hz, the current responses of the membrane were recorded at amplitudes up to 200 mV. At a frequency of 2 Hz, the currents of 4 membranes were recorded at command voltage amplitudes of up to 300 mV and of 6 membranes of up to 200 mV. The current responses to a triangular voltage with a frequency of 0.1 Hz and 1 Hz and an amplitude of 200 mV were studied for two membranes in the presence of nanoparticles.
4. Discussion
All putative processes affecting the nonlinear dependence of capacitance on voltage (electrostriction, solvent redistribution, thermal fluctuations) do not depend on the direction of the field and are approximated by an even degree polynomial. These effects are dissimilar in physical origins and differ in the intensity of the action and different time scales [
1,
2,
3,
4,
5,
10,
25,
26]. In the experiments we describe, which illustrate the adequacy of our nonlinear mathematical model, the main reason for the nonlinear response of the membrane to a triangular voltage at frequencies of 0.1–2 Hz and an amplitude of up to 300 mV is a decrease in the thickness and, possibly, an increase in the area of the bilayer. These changes in geometry arise due to the redistribution of the solvent between the microlenses and the boundary and the electrostrictive compression of the monolayers.
It is known that bilayer lipid membranes with a solvent allow the electric field to create large changes in geometry compared to “dry” membranes [
1,
7,
11] and, accordingly, give well-interpreted current responses to a periodic electrical stimulus. Although the solvent is not part of biological membranes, an artificial membrane can serve as a model for studying the influence of the characteristics of a periodic electrical field (amplitude, frequency, shape) on the nonlinear electrical characteristics of the membrane and their correlation with the properties of the membrane under various conditions.
In an equivalent circuit following the logic of work [
23] related to modeling viscoelastic dielectrics, we have divided the capacitive current of the membrane into two parts. The first part is the current through the purely elastic linear part of the membrane. The second part is the current through the viscoelastic nonlinear part, the polarization of which lags behind the command voltage. In the proposed equivalent circuit, this lag is provided by a resistance in series with the viscoelastic part of the capacitance. As a result of solving a nonlinear differential equation using the small parameter method, we obtain an analytical dependence of the current response of a membrane with quadratic capacitance nonlinearity to a periodic triangular voltage on time and membrane voltage. Already, the first approximation of the method makes it possible to adequately describe the main effects observed in the experiment. A comparison of the calculation with the experiment makes it possible to determine the parameters of the equivalent circuit.
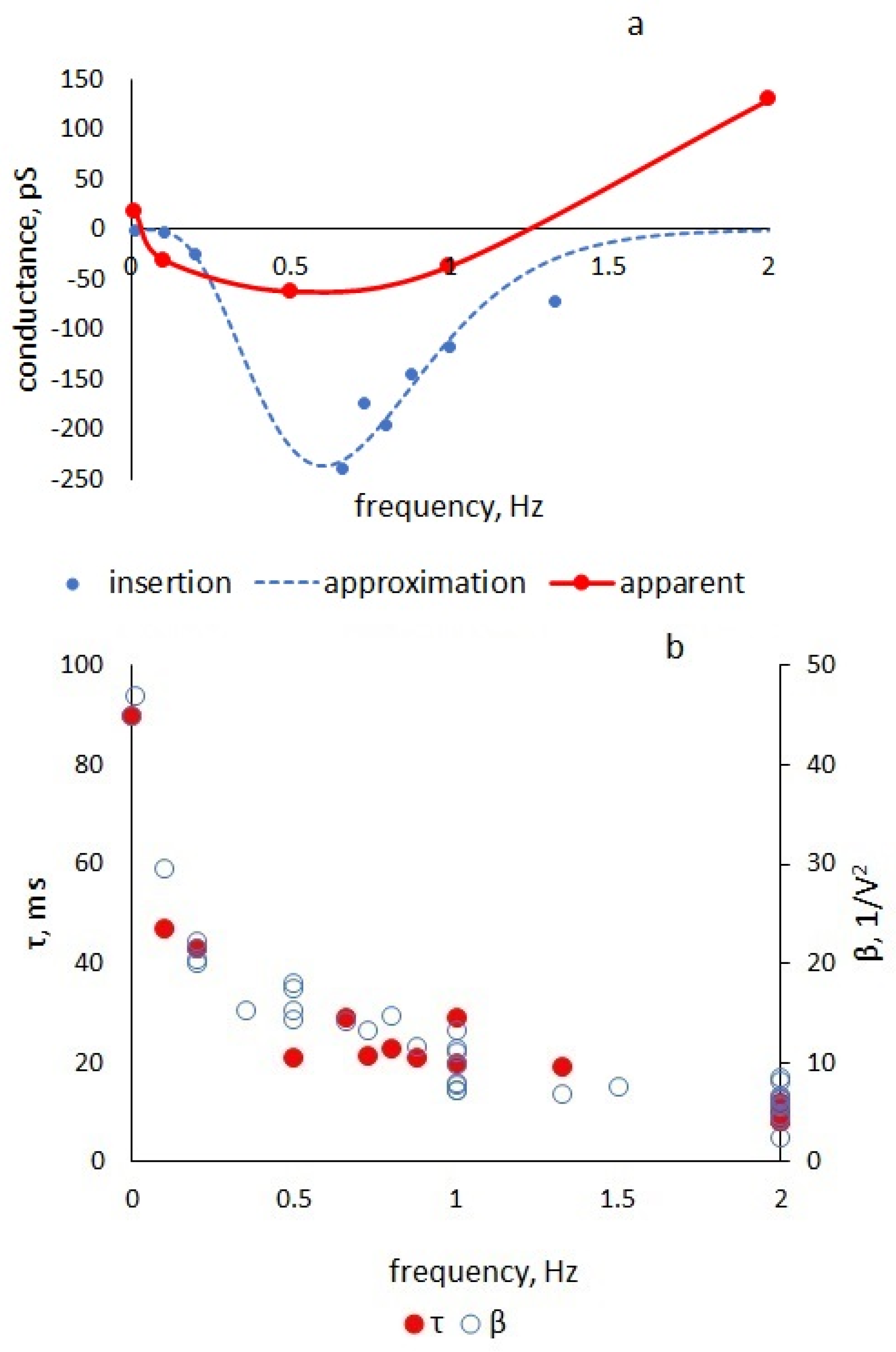
The nonlinearity coefficient
β is maximum in the static mode and decreases with increasing frequency due to a decrease in the number of processes that have time to follow changes in the alternating field. For example, at a frequency of 125 Hz [
10], the coefficient of nonlinearity for the capacitance of BLM from azolectin in decane was 0.6 V
−2, while in our measurements, it varied from 47 to 5 V
−2 in the range of 0.01–2 Hz (
Figure 6).
The series resistance in our equivalent circuit is responsible for the relaxation of the molecular processes occurring in the membrane under the action of an alternating electric field. An analysis of the equation solution in comparison with the experiment shows that the value of the equivalent series resistance enters the solution only through parameter τ, which characterizes the membrane polarization delay with respect to the applied voltage. This parameter, as well as the capacitance nonlinearity coefficient β, depend on the frequency of the periodic command voltage (
Figure 6) and on the relaxation times of capacitive processes occurring in the membrane under the action of an electric field. The frequency range in which we observe changes in the parameters corresponds to the relaxation times of the solvent redistribution and expansion of the plato border 5 ms–5 s [
4,
5,
24,
25,
26]. We use an empirical formula for the nonlinear capacitance in our model without considering the physical nature of the processes leading to nonlinearity. However, we believe that in our particular experimental situation τ depends on the viscosity of the BLM solvent, which is confirmed by studies of the capacitance characteristics for membranes with various solvents [
11].
A similarly shaped current response to a triangular voltage of higher frequencies and, accordingly, associated with other physical processes with corresponding relaxation times was discussed in [
10]. To take into account time dependences, capacitance was introduced into the model, which depends on time according to an exponential law in the same way as in [
22]. Since the processes preventing an instantaneous change in the polarization of the membrane under the action of voltage are associated with energy losses, it seems more physically adequate to use an equivalent circuit with a series of active resistance depending on the frequency.
The existence of hysteresis phenomena in the current response of a nonlinear membrane to a periodic triangular voltage was noted in early works [
5,
6]. These phenomena are also discussed in works where DIB is used as a biological membrane model and dynamic capacitive currents in membranes with different solvents are studied [
7,
8,
11]. It was shown in [
11] that the existence of hysteresis is associated with voltage-dependent changes in the bilayer geometry, which depend on the viscosity of the solvent (decane and hexadecane were compared), temperature and the frequency of the command sinusoidal voltage.
In this context, in [
11], it is proposed to consider DIB as the only variant of the physical implementation of a memcapacitor with a geometry that changes under the action of an electric field, where the effective distance between the capacitor plates or its area changes in some way under the action of the applied voltage.
The authors believe that the presence of so-called pinched hysteresis in the charge–voltage and capacitance–voltage coordinates makes it possible to assert that DIB with a solvent is a realistic and physically justified model for passive memcapacitance in the understanding of [
12,
13,
14]. It is hypothesized that the non-conductive lipid bilayer of cells (without ion channels) may also exhibit capacitive memory driven by voltage-dependent changes of the dielectric, dominated by the hydrophobic core of the bilayer. In DIB experiments, pinched hysteresis in the capacitance–voltage planes results from dynamic changes in interfacial area and hydrophobic thickness, each of which is nonlinearly dependent on voltage. They believe that studies of bilayer lipid membranes can predict new classes of biomimetic, low-power memelements based on soft, organic materials and biomolecules, which, in turn, aid in exploring capacitive memory and susceptibility in neuronal membranes.
We have shown that in the formula for the BLM current response to an applied triangular periodic voltage, due to a combination of nonlinearity and polarization time delays, insertion negative conductance appears, the effect of which is limited by the ion permeability of the membranes. The total apparent conductance is a measure of pinched hysteresis. When the negative insertion conductance is small or compensated for by the ionic conductance of the membranes, the hysteresis and, correspondingly, the memcapacitance properties disappear. Our model allows us to obtain analytical dependences of the curves forming hysteresis loops (expressions (16), (17)) and to analyze the causes and features of these loops.
Analyzing expressions (8, 9), we see that due to the presence of a series resistance in the viscous branch of the equivalent circuit, the voltage
lags from the command triangular voltage U, proportional to the field strength (
Figure 1). Because of this, hysteresis appears on the experimental and theoretical curves
in the presence of capacitance nonlinearity. The general form of the formulas for the current of both half-cycles (13), (14) corresponds to the condition memcapacitance
[
17] at
.
The presence in the current of a term with a negative sign, proportional to the voltage, is determined by the nonlinearity of the capacitance and the shift in the minimum of the capacitive current relative to zero of the command voltage associated with the delay in the polarization of the membrane. As a result (see
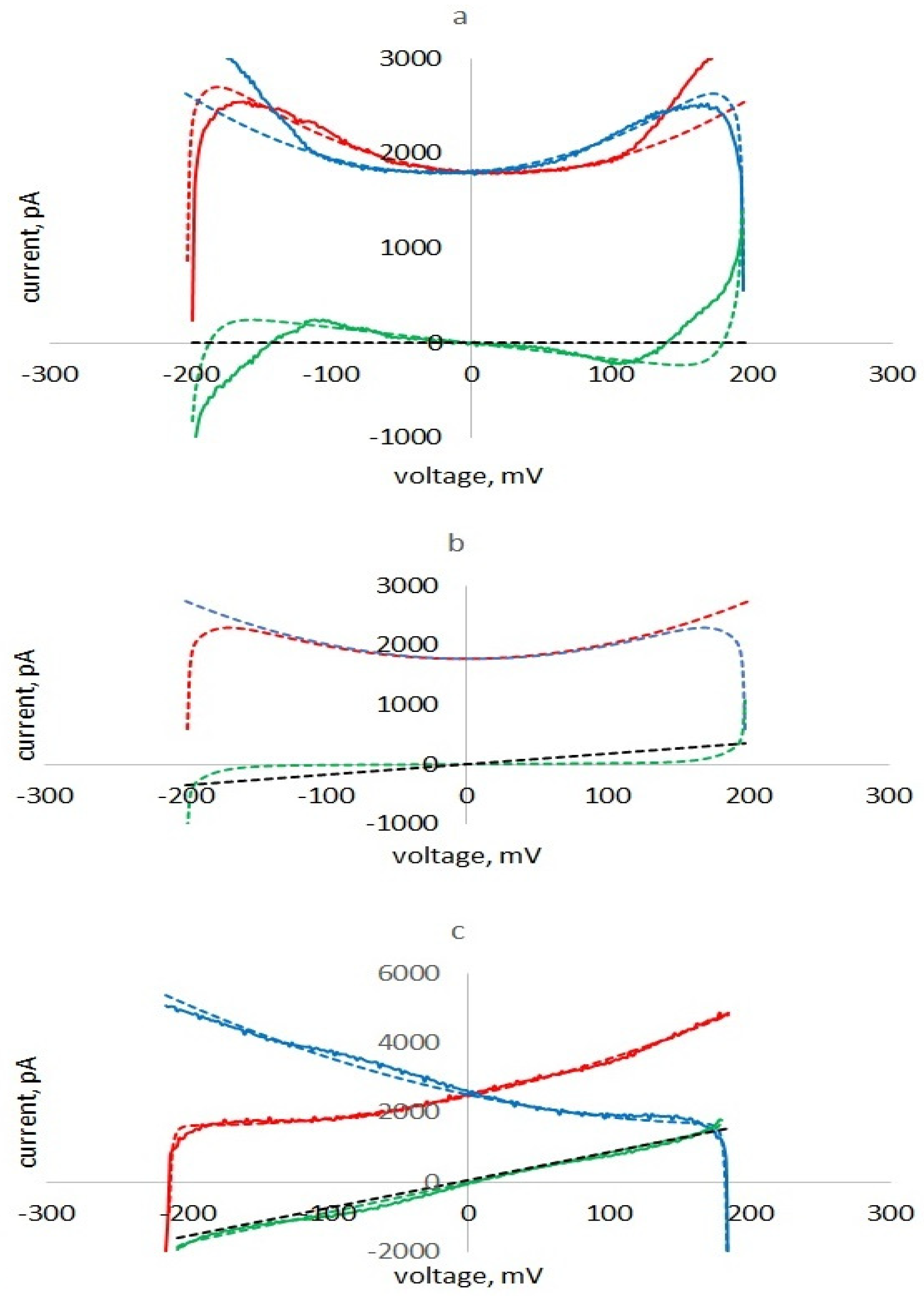
Figure 8a), the part of the half cycle under the left falling part of the parabola (the capacitor gives energy to the circuit) is greater than the part when the capacitor is charged (energy is taken). The difference between these energies is proportional to the negative insertion conductance, which reduces the losses in the membrane. The calculated current curves of two half-cycles, forming pinched hysteresis loops, are shown in
Figure 8a–c for different values of the membrane ionic conductance.
When
the term in expression (13) proportional to
is negative and the expression corresponds to the memcapacitance condition (
Figure 8a). The current minima in the upward and downward half cycles of the applied voltage are shifted to the right (see
Figure 8a). The energy released with a decrease in capacitance is greater than the energy taken from the generator with an increase in capacitance; the apparent conductance is negative, and hysteresis is observed.
When
the current minima in the upward and downward half cycles of applied voltage coincide and are observed at
= 0 (see
Figure 8b). The energy released when the capacitance decreases is equal to the energy taken from the generator with an increase in capacitance; the apparent conductance and the loop width are zero.
When
the term in expression (13) proportional to
is positive and hysteresis is not observed (
Figure 8c).
Thus, the observed pinched hysteresis is associated with nonlinear parametric capacitance effects and is limited by membrane ionic conductance. In
Figure 8a,c, for comparison, experimental curves for membranes with low (a) and high (c) ionic conductance are shown.
Let us calculate the work of the source in a nonlinearly viscous system (without taking into account transient processes), integrating over time over the period the product of the total current and the command voltage. Assuming in Formulas (13) and (14)
,
, we get
Under the conditions of the existence of pinched hysteresis the work is negative and the memcapacitance process is active.
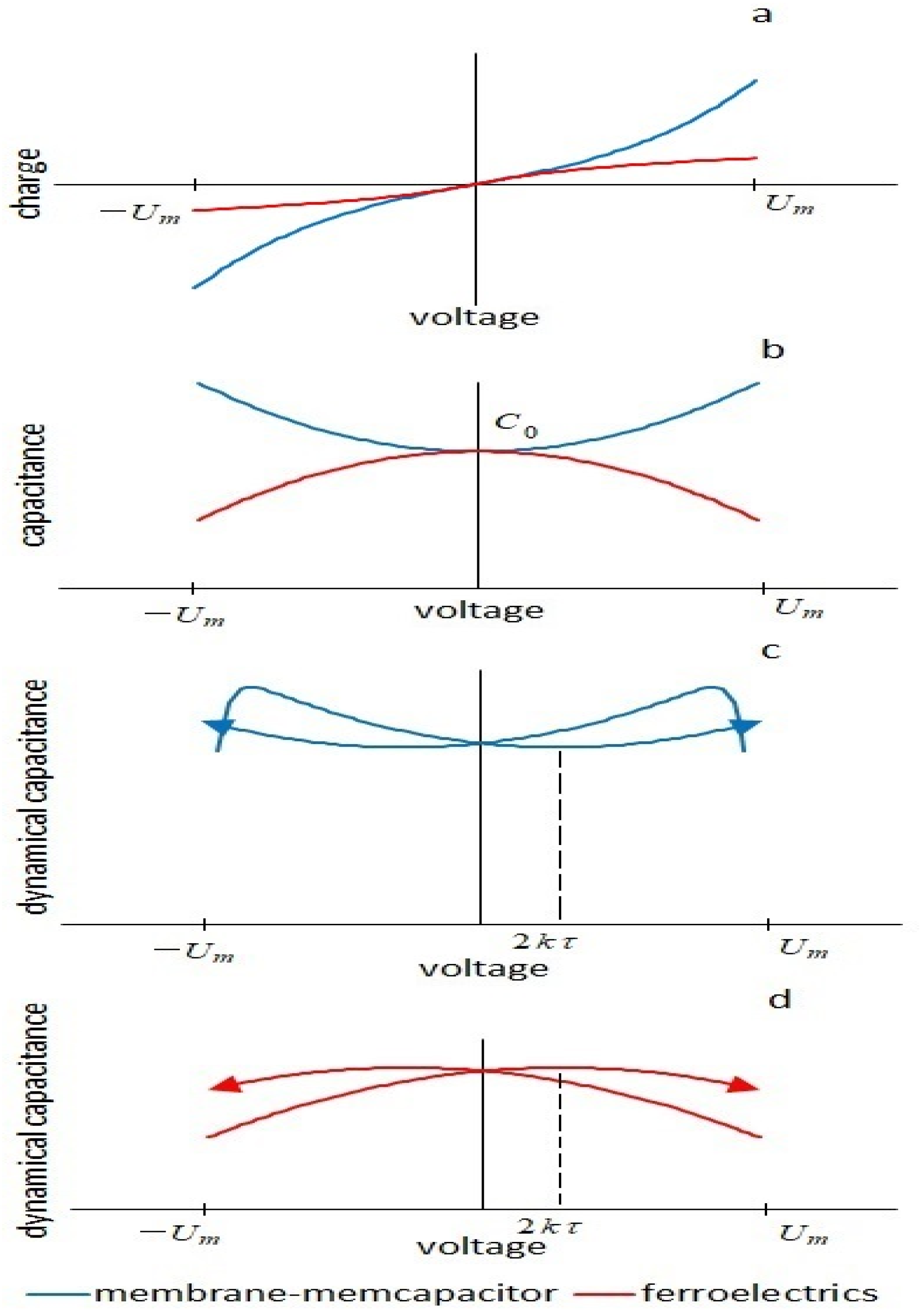
Thus, the combination of nonlinear geometrical changes in the membrane under the action of an electric field and a delay in the polarization of the membrane due to viscous processes does not lead to energy losses, for example, in a ferroelectric, but leads to the appearance of an insertion negative conductance. The importance of the shape of the curve of the nonlinear dependence of capacitance on voltage for viscoelastic effects was noted in [
23] when analyzing a dielectric elastomer. Nonlinear charge–voltage characteristics for a membrane
and a ferroelectric
are inverse functions.
The difference in the energetics of the processes can be shown by solving for a ferroelectric a similar nonlinear differential equation for
q, (see
Supplementary Material, Equations 15S–20S). Current response to the upward half-cycle of the triangular voltage
Comparing this expression with Formula (13), one can see that, unlike the capacitance of the membrane, the capacitance of a ferroelectric decreases with increasing voltage (in the first approximation, along a parabola). This leads to the fact that the insertion conductance in a circuit with ferroelectrics is positive and increases the losses in the system. There is no pinched hysteresis in such a system. Thus, capacitive pinched hysteresis requires a combination of nonlinear capacitance vs. voltage at which the capacitance increases with an increase in the electric field and a delay in the process of polarization of the dielectric, leading to an increase in the fraction of energy given off by the capacitor to the circuit during the upward half-cycle of the triangular voltage. Qualitatively, the energetics of the processes are shown in
Figure 9.



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}








