

Asbestos Fibers Enhance the TMEM16A Channel Activity in Xenopus Oocytes

 ,
,  ,
,  and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Asbestos Fiber Suspensions
2.2. Xenopus Oocyte Preparation
2.3. Electrophysiological Recordings
2.4. Calcium Imaging
2.5. Isolation of a Membrane—Rich Fraction from Xenopus Oocytes
2.6. Isolation of a Membrane—Rich Fraction from the MeT5a Cell Line
2.7. Incubation of Xenopus Oocyte or MeT5a Membrane-Rich Fractions with Asbestos Fibers
2.8. Statistical Analysis
3. Results
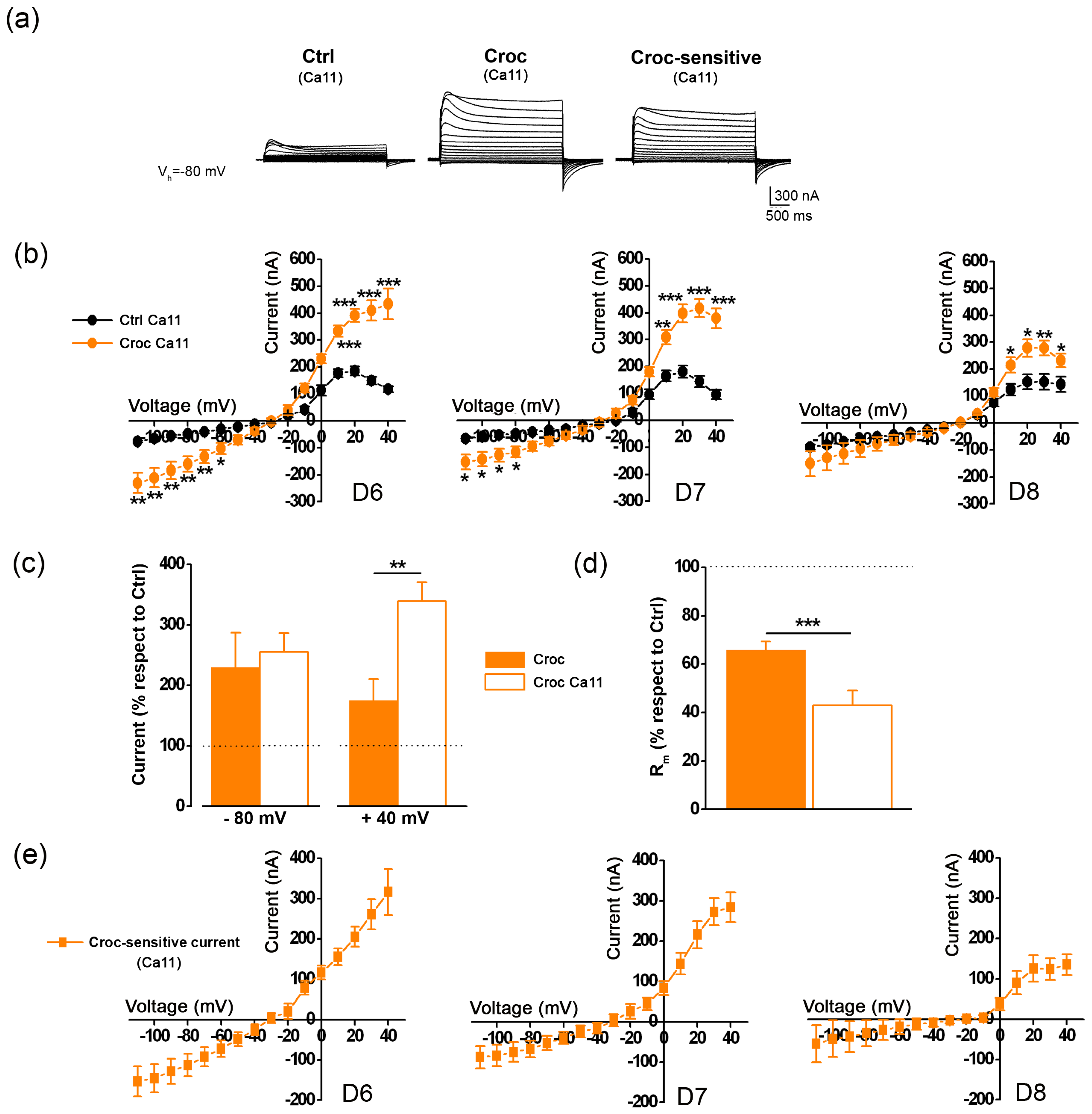
3.1. The Croc-Sensitive Current Is Mediated by TMEM16A Channels Endogenously Expressed in the Oocyte Cell Membrane
3.2. The Effect of Croc Exposure on the TMEM16A-Current Is Modulated by the [Ca2+]e
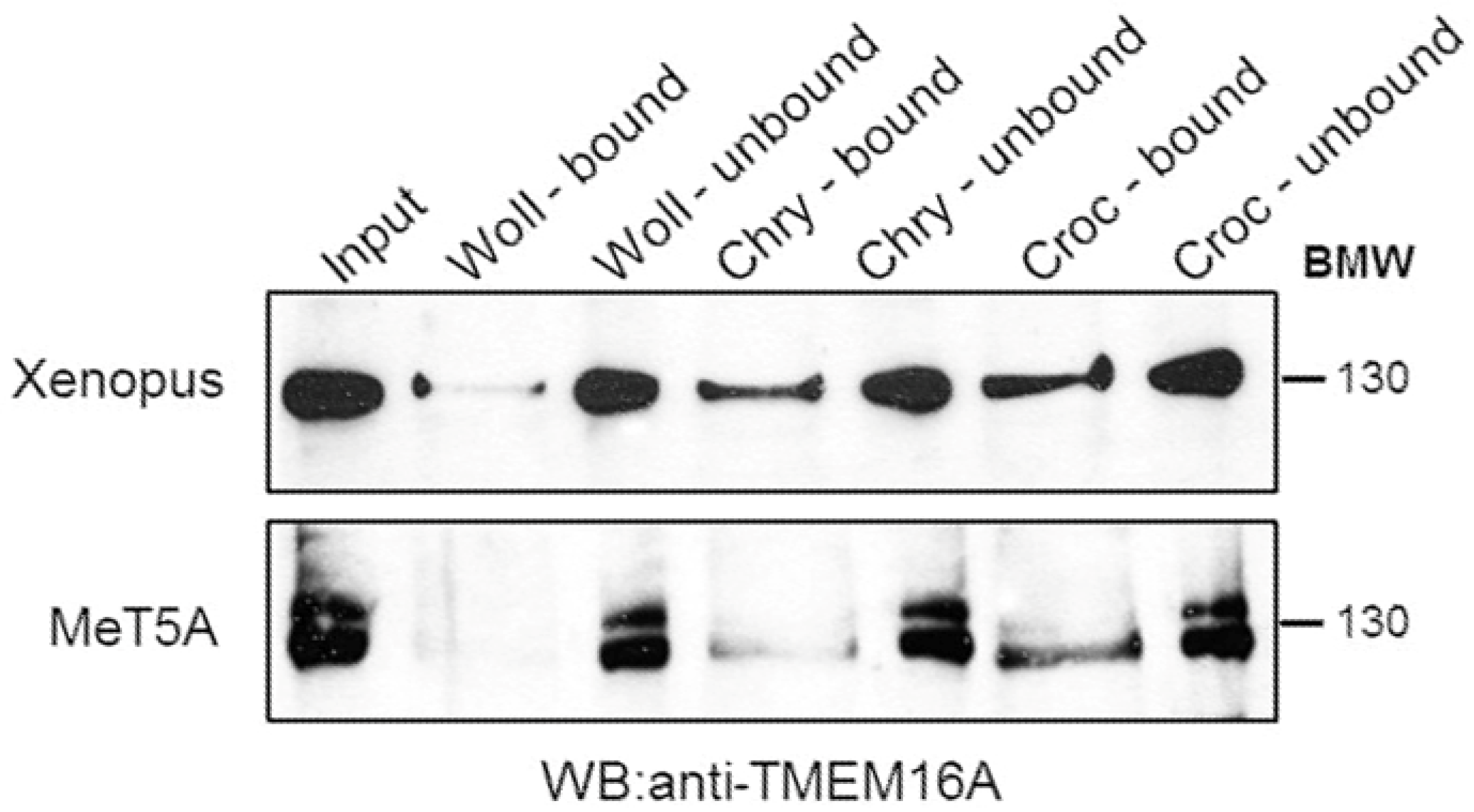
3.3. Western Blotting Experiments
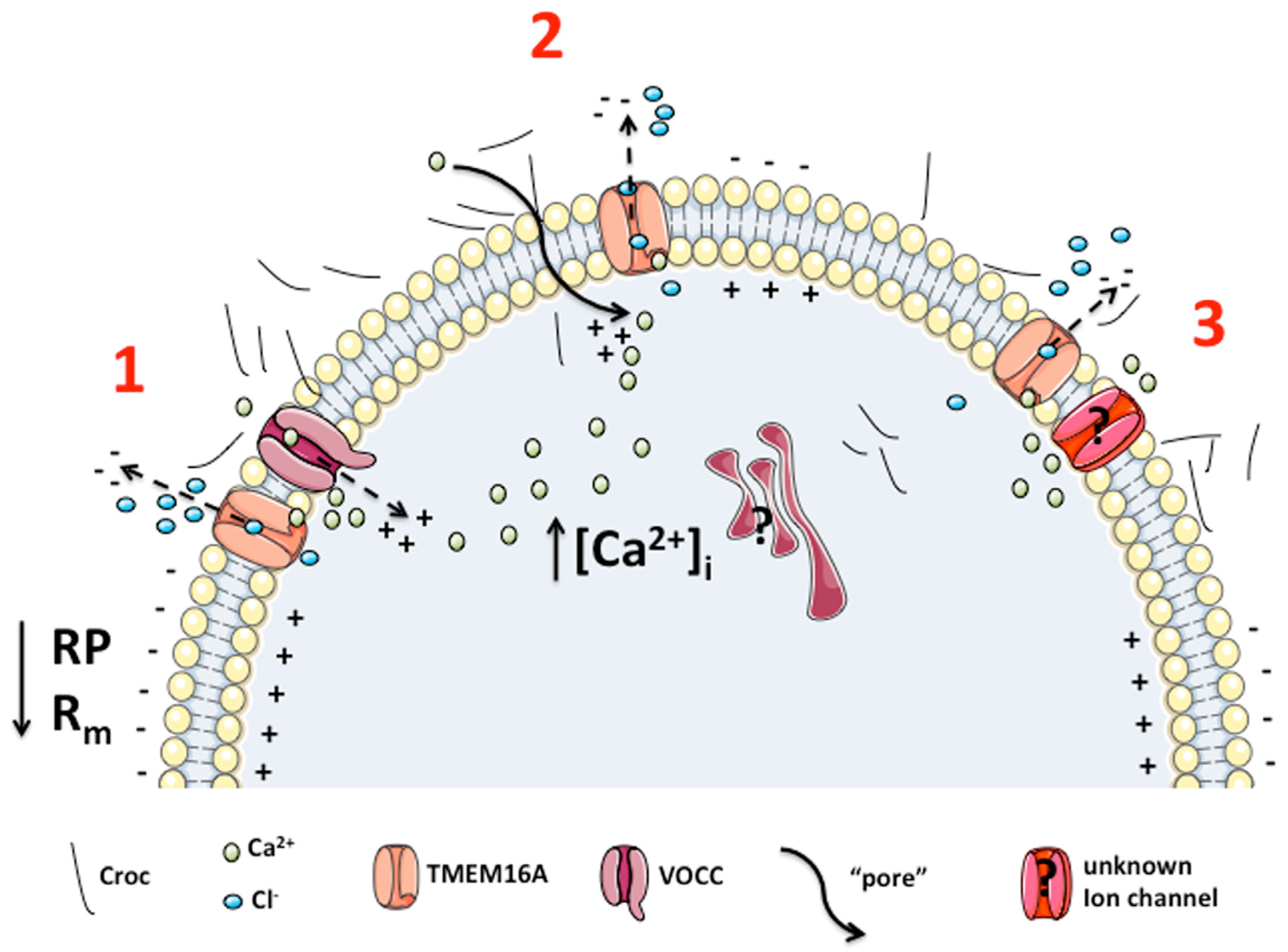
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Klebe, S.; Leigh, J.; Henderson, D.W.; Nurminen, M. Asbestos, smoking and lung cancer: An update. Int. J. Environ. Res. Public Health 2019, 17, 258. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Cheresh, P.; Kamp, D.W. Molecular basis of asbestos-induced lung disease. Annu. Rev. Pathol. Mech. Dis. 2013, 8, 161–187. [Google Scholar] [CrossRef] [PubMed]
- Bernareggi, A.; Ren, E.; Borelli, V.; Vita, F.; Constanti, A.; Zabucchi, G. Xenopus laevis oocytes as a model system for studying the interaction between asbestos fibres and cell membranes. Toxicol. Sci. 2015, 145, 263–272. [Google Scholar] [CrossRef] [PubMed]
- Bernareggi, A.; Conte, G.; Constanti, A.; Borelli, V.; Vita, F.; Zabucchi, G. On the mechanism of the electrophysiological changes and membrane lesions induced by asbestos fiber exposure in Xenopus laevis oocytes. Sci. Rep. 2019, 9, 2014. [Google Scholar] [CrossRef] [PubMed]
- Miledi, R. A calcium-dependent transient outward current in Xenopus laevis oocytes. Proc. R. Soc. Lond. B 1982, 215, 491–497. [Google Scholar] [CrossRef]
- Schroeder, B.C.; Cheng, T.; Jan, Y.N.; Jan, L.Y. Expression Cloning of TMEM16A as a Calcium-Activated Chloride Channel Subunit. Cell 2008, 134, 1019–1029. [Google Scholar] [CrossRef]
- Schreiber, R.; Ousingsawat, J.; Wanitchakool, P.; Sirianant, L.; Benedetto, R.; Reiss, K.; Kunzelmann, K.; Barrett, K.; Chang Chan, H.; Kunzelmann, K. Regulation of TMEM16A/ANO1 and TMEM16F/ANO6 ion currents and phospholipid scrambling by Ca2+ and plasma membrane lipid. J. Physiol. 2018, 596, 217–229. [Google Scholar] [CrossRef]
- Courjaret, R.; Hodeify, R.; Hubrack, S.; Ibrahim, A.; Dib, M.; Daas, S.; Machaca, K. The Ca 2+-activated Cl − channel Ano1 controls microvilli length and membrane surface area in the oocyte. Cell Sci. 2016, 129, 2548–2558. [Google Scholar] [CrossRef]
- Wang, H.; Zou, L.; Ma, K.; Yu, J.; Wu, H.; Wei, M.; Xiao, Q. Cell-specific mechanisms of TMEM16A Ca2+-activated chloride channel in cancer. Mol. Cancer 2017, 16, 152. [Google Scholar] [CrossRef]
- Terashima, H.; Picollo, A.; Accardi, A. Purified TMEM16A is sufficient to form Ca2+-activated Cl− channels. Proc. Natl. Acad. Sci. USA 2013, 110, 19354–19359. [Google Scholar] [CrossRef] [Green Version]
- Xiao, Q.; Yu, K.; Perez-Cornejo, P.; Cui, Y.; Arreola, J.; Hartzell, H.C. Voltage- and calcium-dependent gating of TMEM16A/Ano1 chloride channels are physically coupled by the first intracellular loop. Proc. Natl. Acad. Sci. USA 2011, 108, 8891–8896. [Google Scholar] [CrossRef]
- Liu, Y.; Zhang, H.; Huang, D.; Qi, J.; Xu, J.; Gao, H.; Du, X.; Gamper, N.; Zhang, H. Characterization of the effects of Cl − channel modulators on TMEM16A and bestrophin-1 Ca 2+ activated Cl—channels. Pflugers Arch. Eur. J. Physiol. 2015, 467, 1417–1430. [Google Scholar] [CrossRef]
- West, R.B.; Corless, C.L.; Chen, X.; Rubin, B.P.; Subramanian, S.; Montgomery, K.; Zhu, S.; Ball, C.A.; Nielsen, T.O.; Patel, R.; et al. The Novel Marker, DOG1, Is Expressed Ubiquitously in Gastrointestinal Stromal Tumors Irrespective of KIT or PDGFRA Mutation Status. Am. J. Pathol. 2004, 165, 107–113. [Google Scholar] [CrossRef]
- Espinosa, I.; Lee, C.H.; Kim, M.K.; Rouse, B.T.; Subramanian, S.; Montgomery, K.; Varma, S.; Corless, C.L.; Heinrich, M.C.; Smith, K.S.; et al. A Novel Monoclonal Antibody Against DOG1 is a Sensitive and Specific Marker for Gastrointestinal Stromal Tumors. Am. J. Surg. Pathol. 2008, 32, 210–218. [Google Scholar] [CrossRef]
- Ruiz, C.; Martins, J.R.; Rudin, F.; Schneider, S.; Dietsche, T.; Fischer, C.A.; Tornillo, L.; Terracciano, L.M.; Schreiber, R.; Bubendorf, L.; et al. Enhanced Expression of ANO1 in Head and Neck Squamous Cell Carcinoma Causes Cell Migration and Correlates with Poor Prognosis. PLoS ONE 2012, 7, e43265. [Google Scholar] [CrossRef]
- Ji, Q.; Guo, S.; Wang, X.; Pang, C.; Zhan, Y.; Chen, Y.; An, H. Recent advances in TMEM16A: Structure, function, and disease. J. Cell Physiol. 2019, 234, 7856–7873. [Google Scholar] [CrossRef]
- Crottès, D.; Jan, L.Y. The multifaceted role of TMEM16A in cancer. Cell Calcium 2019, 82, 102050. [Google Scholar] [CrossRef]
- Oh, S.J.; Hwang, S.J.; Jung, J.; Yu, K.; Kim, J.; Choi, J.Y.; Hartzell, H.C.; Roh, E.J.; Lee, C.J. MONNA, a Potent and Selective Blocker for Transmembrane Protein with Unknown Function 16/Anoctamin-1. Mol. Pharmacol. 2013, 84, 726–735. [Google Scholar] [CrossRef]
- Seo, Y.; Lee, H.K.; Park, J.; Jeon, D.K.; Jo, S.; Jo, M.; Namkung, W. Ani9, A Novel Potent Small-Molecule ANO1 Inhibitor with Negligible Effect on ANO2. PLoS ONE 2016, 11, e0155771. [Google Scholar] [CrossRef]
- Kohyama, N.; Shinohara, Y.; Suzuki, Y. Mineral phases and some reexamined characteristics of the International Union Against Cancer standard asbestos samples. Am. J. Ind. Med. 1996, 30, 515–528. [Google Scholar] [CrossRef]
- Governa, M.; Camilucci, L.; Amati, M.; Visonà, I.; Valentino, M.; Botta, G.C.; Campopiano, A.; Canizza, C. Wollastonite fibers in vitro generate reactive oxygen species able to lyse erythrocytes and activate the complement alternate pathway. Toxicol. Sci. 1998, 44, 32–38. [Google Scholar] [CrossRef] [PubMed]
- Clémençon, B.; Lüscher, B.P.; Fine, M.; Baumann, M.U.; Surbek, D.V.; Bonny, O.; Hediger, M.A. Expression, purification, and structural insights for the human uric acid transporter, GLUT9, using the Xenopus laevis oocytes system. PloS ONE 2014, 9, e108852. [Google Scholar] [CrossRef] [PubMed]
- Wozniak, K.L.; Wesley, A.; Phelps, W.A.; Tembo, M.; Lee, M.T.; Carlson, A.E. The TMEM16A channel mediates the fast polyspermy block in Xenopus laevis. J. Gen. Physiol. 2018, 150, 1249–1259. [Google Scholar] [CrossRef] [PubMed]
- Sobczak, K.; Bangel-Ruland, N.; Leier, G.; Weber, W.M. Endogenous transport systems in the Xenopus laevis oocyte plasma membrane. Methods 2010, 51, 183–189. [Google Scholar] [CrossRef]
- Lory, P.; Rassendren, F.A.; Richard, S.; Tiaho, F.; Nargeot, J. Characterization of voltage-dependent calcium channels expressed in Xenopus oocytes injected with mRNA from rat heart. J. Physiol. 1990, 429, 95–112. [Google Scholar] [CrossRef]
- Dascal, N.; Chilcott, G.; Lester, H.A. Recording of voltage and Ca(2+)-dependent currents in Xenopus oocytes using an intracellular perfusion method. J. Neurosci. Methods 1991, 39, 29–38. [Google Scholar] [CrossRef]
- Yao, Y.; Tsien, R.Y. Calcium current activated by depletion of calcium stores in Xenopus oocytes. J. Gen. Physiol. 1997, 6, 703–715. [Google Scholar] [CrossRef]
- Caputo, A.; Caci, E.; Ferrera, L.; Pedemonte, N.; Barsanti, C.; Sondo, E.; Pfeffer, U.; Ravazzolo, R.; Zegarra-Moran, O.; Galietta, L.J. TMEM16A, a membrane protein associated with calcium-dependent chloride channel activity. Science 2008, 322, 590–594. [Google Scholar] [CrossRef]
- Callamaras, N.; Parker, I. Ca(2+)-dependent activation of Cl(-) currents in Xenopus oocytes is modulated by voltage. Am. J. Physiol. Cell Physiol. 2000, 78, C667–C675. [Google Scholar] [CrossRef]
- Carvacho, I.; Piesche, M.; Maier, T.J.; Machaca, K. Ion Channel Function During Oocyte Maturation and Fertilization. Front. Cell Dev. Biol. 2018, 6, 63. [Google Scholar] [CrossRef] [Green Version]
- Nagai, H.; Ishihara, T.; Lee, W.-H.; Ohara, H.; Okazaki, Y.; Okawa, K.; Toyokuni, S. Asbestos surface provides a niche for oxidative modification. Cancer Sci. 2011, 102, 2118–2125. [Google Scholar] [CrossRef]
- Borelli, V.; Brochetta, C.; Melato, M.; Rizzardi, C.; Polentarutti, M.; Busatto, C.; Vita, F.; Abbate, R.; Gotter, R.; Zabucchi, G. A procedure for the isolation of asbestos bodies from lung tissue by exploiting their magnetic properties: A new approach to asbestos body study. J. Toxicol. Environ. Health A 2007, 70, 1232–1240. [Google Scholar] [CrossRef] [PubMed]
- Pascolo, L.; Borelli, V.; Canzonieri, V.; Gianoncelli, A.; Birarda, G.; Bedolla, D.E.; Salomé, M.; Vaccari, L.; Calligaro, C.; Cotte, M.; et al. Differential protein folding and chemical changes in lung tissues exposed to asbestos or particulates. Sci. Rep. 2015, 5, 12129. [Google Scholar] [CrossRef]
- Borelli, V.; Trevisan, E.; Vita, F.; Zabucchi, G. The Secretory Response of Rat Peritoneal Mast Cells on Exposure to Mineral Fibers. Int. J. Environ. Res. Public. Health 2018, 15, 104. [Google Scholar] [CrossRef]
- Maxim, L.D.; Niebo, R.; Utell, M.J.; McConnell, E.E.; Larosa, S.; Segrave, A.M. Wollastonite toxicity: An update. Inhal. Toxicol. 2014, 26, 95–112. [Google Scholar] [CrossRef]
- Lim, Y.; Kim, S.H.; Kim, K.A.; Oh, M.W.; Lee, K.H. Involvement of protein kinase C, phospholipase C, and protein tyrosine kinase pathways in oxygen radical generation by asbestos-stimulated alveolar macrophage. Environ. Health Perspect. 1997, 105, 1325–1327. [Google Scholar] [CrossRef]
- Song, Y.; Gao, J.; Guan, L.; Chen, X.; Gao, J.; Wang, K. Inhibition of ANO1/TMEM16A induces apoptosis in human prostate carcinoma cells by activating TNF-α signaling. Cell Death Dis. 2018, 9, 703. [Google Scholar] [CrossRef]
- IARC Working Group on the Evaluation of Carcinogenic Risks to Humans. A review of human carcinogens. In Arsenic, Metals, Fibres and Dusts; International Agency for Research on Cancer: Lyon, France, 2012; Volume 100C, ISBN 978-92-832-1320-8. Available online: https://www.ncbi.nlm.nih.gov/books/NBK304374/ (accessed on 26 January 2023).
- Doll, R. Mortality from lung cancer in asbestos workers. Br. J. Ind. Med. 1955, 12, 81–86. [Google Scholar] [CrossRef]
- Awan, K.H.; Hegde, R.; Cheever, V.J.; Carroll, W.; Khan, S.; Patil, S.; Warnakulasuriya, S. Oral and pharyngeal cancer risk associated with occupational carcinogenic substances: Systematic review. Head Neck 2018, 40, 2724–2732. [Google Scholar] [CrossRef]
- Chandrashekar, D.S.; Bashel, B.; Balasubramanya, S.A.H.; Creighton, C.J.; Ponce-Rodriguez, I.; Chakravarthi, B.V.S.K.; Varambally, S. UALCAN: A Portal for Facilitating Tumor Subgroup Gene Expression and Survival Analyses. Neoplasia 2017, 19, 649–658. [Google Scholar] [CrossRef]
- Jia, L.; Liu, W.; Guan, L.; Lu, M.; Wang, K. Inhibition of Calcium-Activated Chloride Channel ANO1/TMEM16A Suppresses Tumor Growth and Invasion in Human Lung Cancer. PLoS One 2015, 10, e0136584. [Google Scholar] [CrossRef] [PubMed]
- Perez-Cornejo, P.; Gokhale, A.; Duran, C.; Cui, Y.; Xiao, Q.; Hartzell, H.C.; Faundez, V. Anoctamin 1 (Tmem16A) Ca2+-activated chloride channel stoichiometrically interacts with an ezrin-radixin-moesin network. Proc. Natl. Acad. Sci. USA 2012, 109, 10376–10381. [Google Scholar] [CrossRef] [PubMed]
- Clucas, J.; Valderrama, F. ERM proteins in cancer progression. J. Cell Sci. 2015, 128, 267–275. [Google Scholar] [CrossRef] [PubMed]
- Cuddapah, V.A.; Sontheimer, H. Ion channels and tranporters [corrected] in cancer. 2. Ion channels and the control of cancer cell migration. Am. J. Physiol. Cell Physiol. 2011, 301, C541–C549. [Google Scholar] [CrossRef]
- Ito, F.; Kato, K.; Yanatori, I.; Murohara, T.; Toyokuni, S. Ferroptosis-dependent extracellular vesicles from macrophage contribute to asbestos-induced mesothelial carcinogenesis through loading ferritin. Redox Biol. 2021, 47, 102174. [Google Scholar] [CrossRef]
- Zangari, M.; Borelli, V.; Bernareggi, A.; Zabucchi, G. Asbestos fibers promote iron oxidation and compete with apoferritin enzymatic activity. J Toxicol Environ Health A 2023, 86, 69–73. [Google Scholar] [CrossRef]
- Neumann, V.; Löseke, S.; Tannapfel, A. Mesothelioma and analysis of tissue fiber content. Recent Results Cancer Res. 2011, 189, 79–95. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bernareggi, A.; Zangari, M.; Constanti, A.; Zacchi, P.; Borelli, V.; Mangogna, A.; Lorenzon, P.; Zabucchi, G. Asbestos Fibers Enhance the TMEM16A Channel Activity in Xenopus Oocytes. Membranes 2023, 13, 180. https://doi.org/10.3390/membranes13020180
Bernareggi A, Zangari M, Constanti A, Zacchi P, Borelli V, Mangogna A, Lorenzon P, Zabucchi G. Asbestos Fibers Enhance the TMEM16A Channel Activity in Xenopus Oocytes. Membranes. 2023; 13(2):180. https://doi.org/10.3390/membranes13020180
Chicago/Turabian StyleBernareggi, Annalisa, Martina Zangari, Andrew Constanti, Paola Zacchi, Violetta Borelli, Alessandro Mangogna, Paola Lorenzon, and Giuliano Zabucchi. 2023. "Asbestos Fibers Enhance the TMEM16A Channel Activity in Xenopus Oocytes" Membranes 13, no. 2: 180. https://doi.org/10.3390/membranes13020180
APA StyleBernareggi, A., Zangari, M., Constanti, A., Zacchi, P., Borelli, V., Mangogna, A., Lorenzon, P., & Zabucchi, G. (2023). Asbestos Fibers Enhance the TMEM16A Channel Activity in Xenopus Oocytes. Membranes, 13(2), 180. https://doi.org/10.3390/membranes13020180