
Insights into Early Steps of Decanoic Acid Self-Assemblies under Prebiotic Temperatures Using Molecular Dynamics Simulations
 , ,
, ,
Abstract
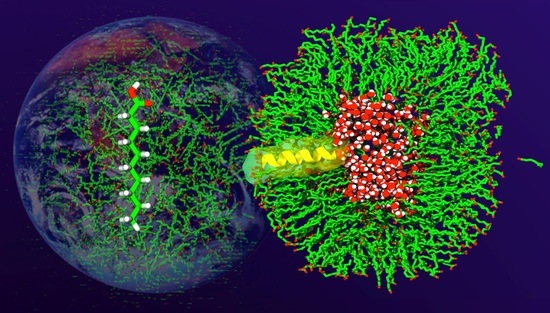
:
1. Introduction
2. Methods
2.1. Preparation of Decanoic Acid Vesicles
2.2. Dynamic Light Scattering (DLS) and Transmission Electron Microscopy
2.3. Molecules Built and Systems Set Up
2.4. Molecular Dynamics Simulations
2.5. Trajectory Analysis Tools
3. Results
3.1. Exploration of a DA Vesicle under Standard Conditions
3.2. DA Spontaneous Aggregation into Vesicles
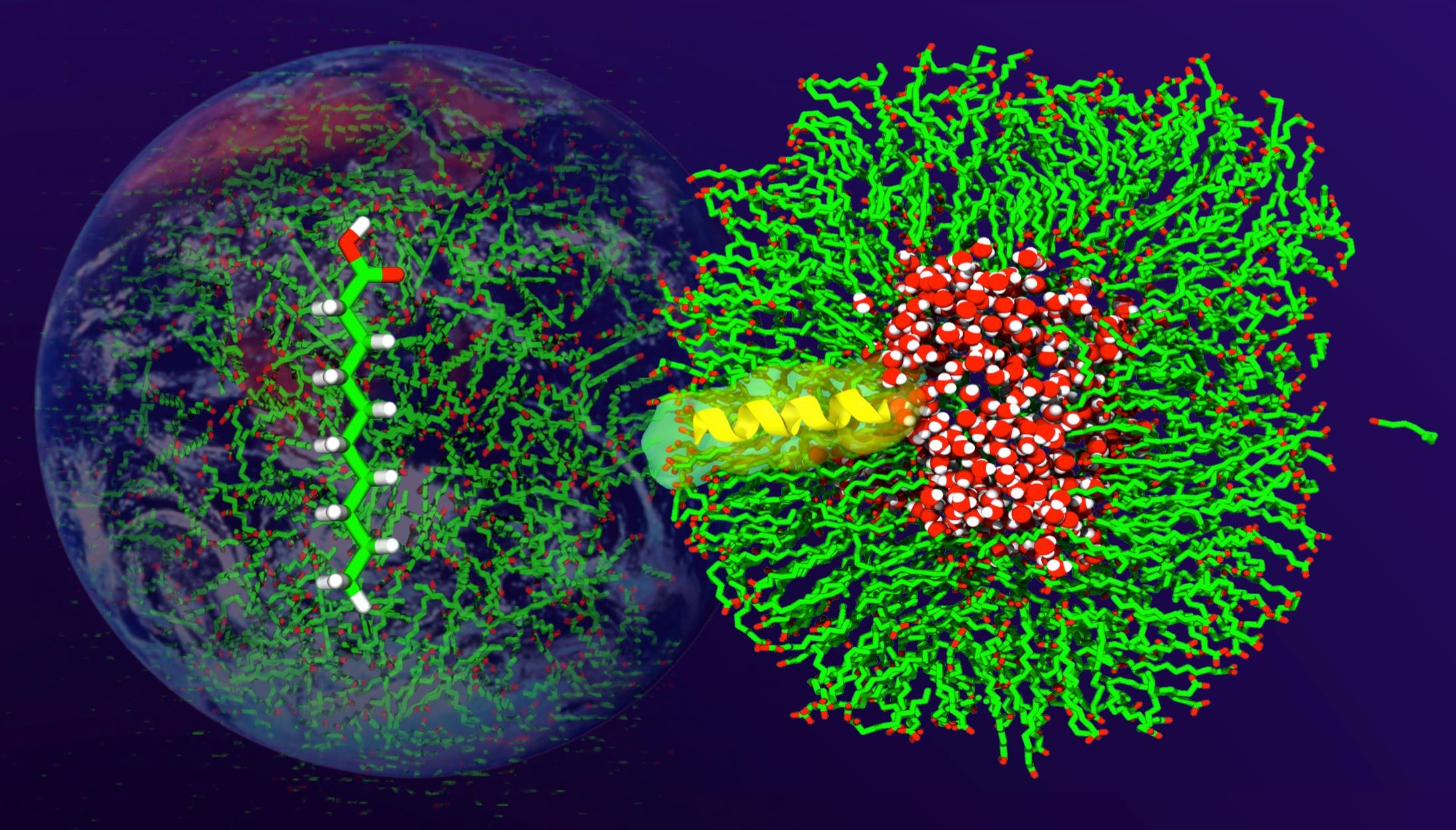
3.3. Spontaneous DA Vesicle Formation in a Wide Range of Temperatures
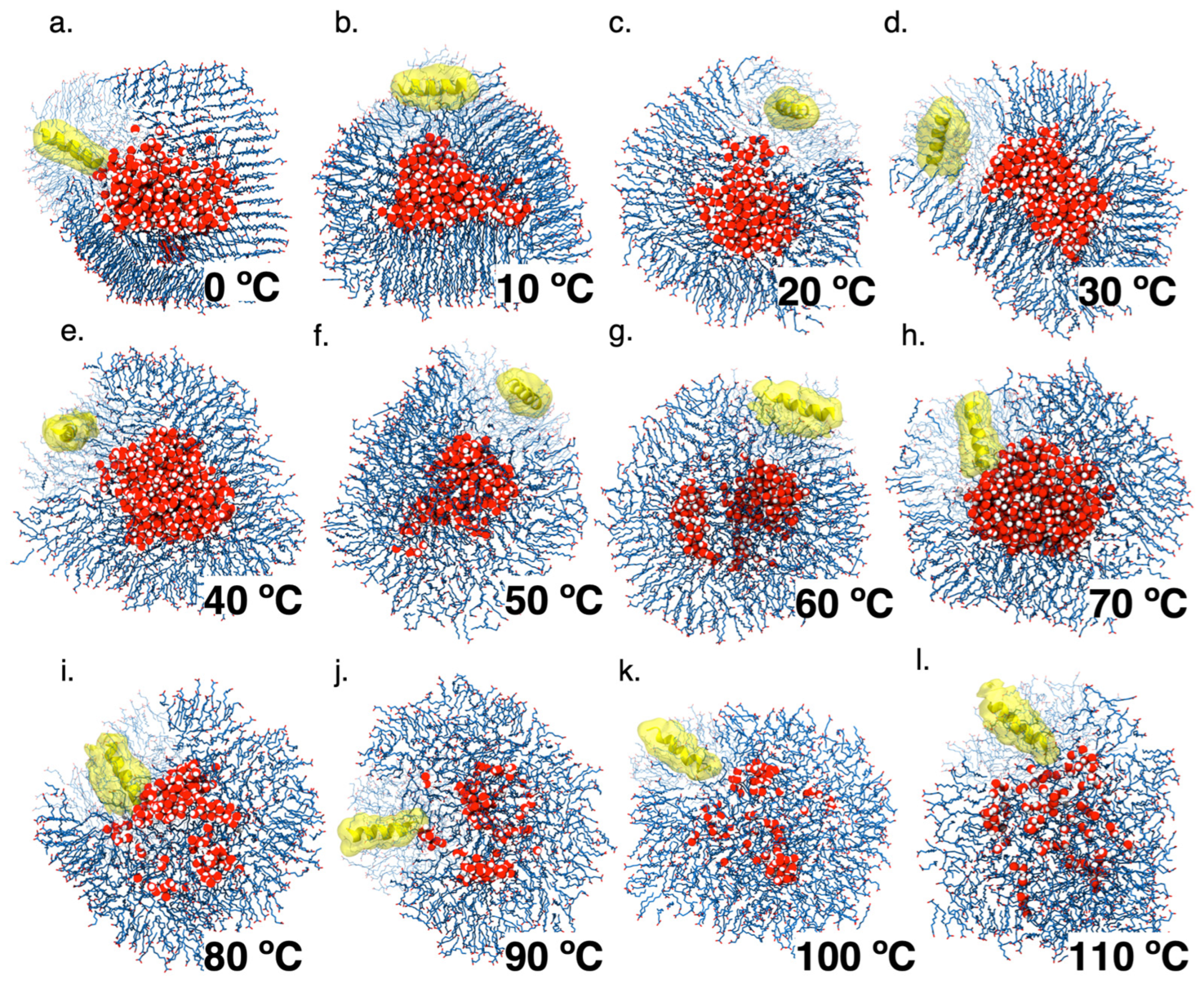
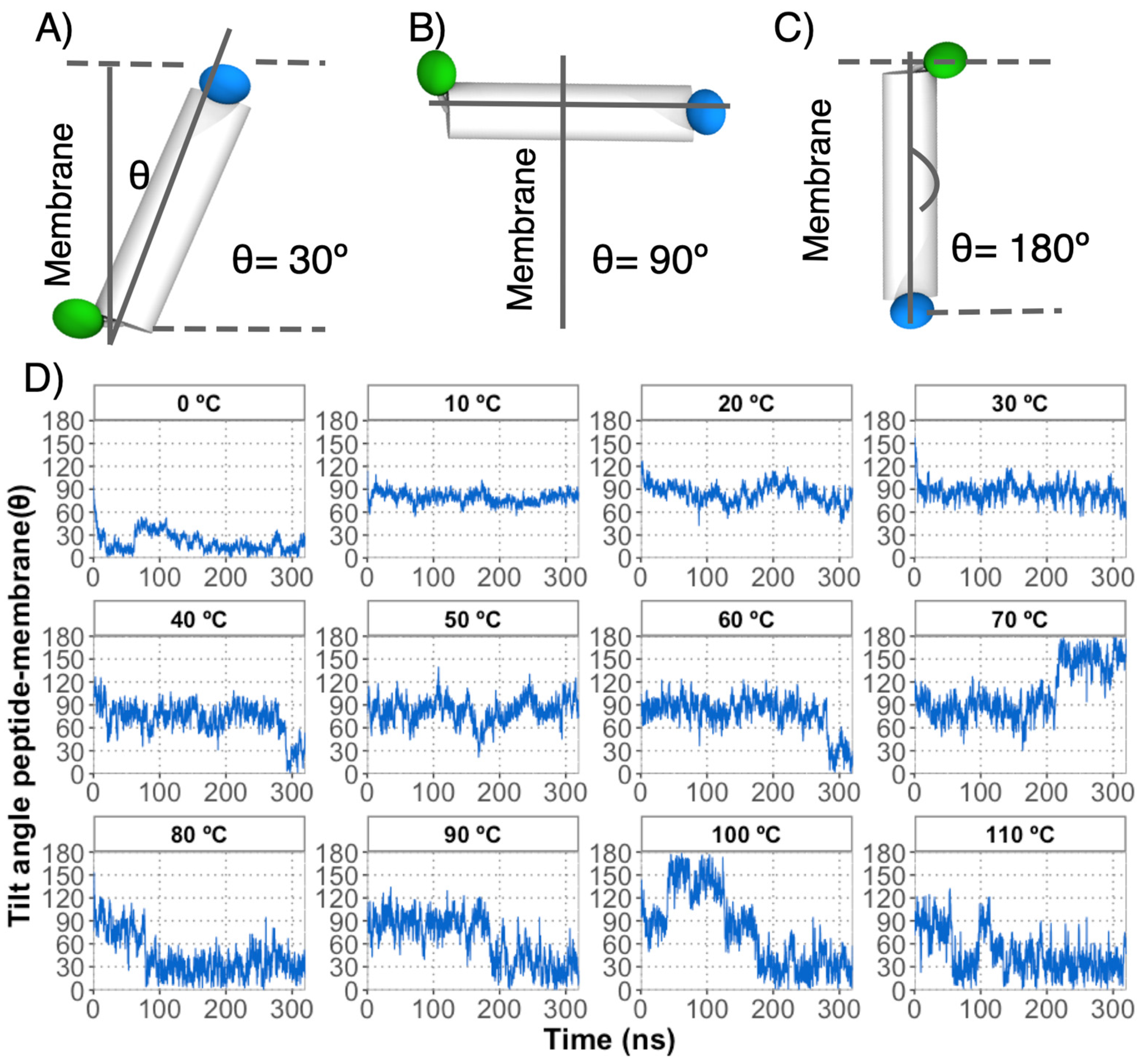
3.4. Peptide Interaction with Decanoic Acid Vesicles as a Key to Understanding the Encapsulation Process
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Black, R.A.; Blosser, M.C. A Self-Assembled Aggregate Composed of a Fatty Acid Membrane and the Building Blocks of Biological Polymers Provides a First Step in the Emergence of Protocells. Life 2016, 6, 33. [Google Scholar] [CrossRef] [PubMed]
- Chen, I.A.; Walde, P. From Self-Assembled Vesicles to Protocells. Cold Spring Harb. Perspect. Biol. 2010, 2, a002170. [Google Scholar] [CrossRef] [PubMed]
- Gözen, I.; Köksal, E.S.; Põldsalu, I.; Xue, L.; Spustova, K.; Pedrueza-Villalmanzo, E.; Ryskulov, R.; Meng, F.; Jesorka, A. Protocells: Milestones and Recent Advances. Small 2022, 18, 2106624. [Google Scholar] [CrossRef] [PubMed]
- Maurer, S.E.; Nguyen, G. Prebiotic Vesicle Formation and the Necessity of Salts. Orig. Life Evol. Biosph. 2016, 46, 215–222. [Google Scholar] [CrossRef] [PubMed]
- Schmitt-Kopplin, P.; Gabelica, Z.; Gougeon, R.D.; Fekete, A.; Kanawati, B.; Harir, M.; Gebefuegi, I.; Eckel, G.; Hertkorn, N. High molecular diversity of extraterrestrial organic matter in Murchison meteorite revealed 40 years after its fall. Proc. Natl. Acad. Sci. USA 2010, 107, 2763–2768. [Google Scholar] [CrossRef]
- Apel, C.L.; Deamer, D.W.; Mautner, M.N. Self-assembled vesicles of monocarboxylic acids and alcohols: Conditions for stability and for the encapsulation of biopolymers. Biochim. Biophys. Acta Biomembr. 2002, 1559, 1–9. [Google Scholar] [CrossRef]
- Kitadai, N. Thermodynamic Prediction of Glycine Polymerization as a Function of Temperature and pH Consistent with Experimentally Obtained Results. J. Mol. Evol. 2014, 78, 171–187. [Google Scholar] [CrossRef]
- Kitadai, N.; Maruyama, S. Origins of building blocks of life: A review. Geosci. Front. 2018, 9, 1117–1153. [Google Scholar] [CrossRef]
- Maurer, S.E.; Monnard, P.-A. Primitive Membrane Formation, Characteristics and Roles in the Emergent Properties of a Protocell. Entropy 2011, 13, 466–484. [Google Scholar] [CrossRef]
- Wellen, B.A.; Lach, E.A.; Allen, H.C. Surface pKa of octanoic, nonanoic, and decanoic fatty acids at the air–water interface: Applications to atmospheric aerosol chemistry. Phys. Chem. Chem. Phys. 2017, 19, 26551–26558. [Google Scholar] [CrossRef]
- Kanicky, J.R.; Shah, D.O. Effect of Premicellar Aggregation on the pKa of Fatty Acid Soap Solutions. Langmuir 2003, 19, 2034–2038. [Google Scholar] [CrossRef]
- Monnard, P.A.; Deamer, D.W. Membrane self-assembly processes: Steps toward the first cellular life. Anat. Rec. 2002, 268, 196–207. [Google Scholar] [CrossRef] [PubMed]
- Fitz, D.; Reiner, H.; Rode, B.M. Chemical evolution toward the origin of life. Pure Appl. Chem. 2007, 79, 2101–2117. [Google Scholar] [CrossRef]
- Holm, N.G. Chapter 1 Why are hydrothermal systems proposed as plausible environments for the origin of life? Orig. Life Evol. Biosph. 1992, 22, 5–14. [Google Scholar] [CrossRef]
- Deamer, D. The Role of Lipid Membranes in Life’s Origin. Life 2017, 7, 5. [Google Scholar] [CrossRef]
- Murillo-Sánchez, S.; Beaufils, D.; Mañas, J.M.G.; Pascal, R.; Ruiz-Mirazo, K. Fatty acids’ double role in the prebiotic formation of a hydrophobic dipeptide. Chem. Sci. 2016, 7, 3406–3413. [Google Scholar] [CrossRef]
- Adamala, K.; Szostak, J.W. Competition between model protocells driven by an encapsulated catalyst. Nat. Chem. 2013, 5, 495–501. [Google Scholar] [CrossRef]
- Wei, C.; Pohorille, A. Fast bilayer-micelle fusion mediated by hydrophobic dipeptides. Biophys. J. 2021, 120, 2330–2342. [Google Scholar] [CrossRef]
- Blake, R.E.; Chang, S.J.; Lepland, A. Phosphate oxygen isotopic evidence for a temperate and biologically active Archaean ocean. Nature 2010, 464, 1029–1032. [Google Scholar] [CrossRef]
- Catling, D.C.; Zahnle, K.J. The Archean atmosphere. Sci. Adv. 2020, 6, eaax1420. [Google Scholar] [CrossRef]
- Kahana, A.; Lancet, D. Protobiotic Systems Chemistry Analyzed by Molecular Dynamics. Life 2019, 9, 38. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Wei, Y.; Gao, F.; Yuan, S.; Liu, C. Origins of entropy change for the amphiphilic molecule in micellization: A molecular dynamics study. Phys. Chem. Chem. Phys. 2016, 18, 11357–11361. [Google Scholar] [CrossRef] [PubMed]
- Pohorille, A.; Deamer, D. Self-assembly and function of primitive cell membranes. Res. Microbiol. 2009, 160, 449–456. [Google Scholar] [CrossRef]
- Faramarzi, S.; Bonnett, B.; Scaggs, C.A.; Hoffmaster, A.; Grodi, D.; Harvey, E.; Mertz, B. Molecular Dynamics Simulations as a Tool for Accurate Determination of Surfactant Micelle Properties. Langmuir 2017, 33, 9934–9943. [Google Scholar] [CrossRef] [PubMed]
- Lebecque, S.; Crowet, J.; Nasir, M.; Deleu, M.; Lins, L. Molecular dynamics study of micelles properties according to their size. J. Mol. Graph. Model. 2017, 72, 6–15. [Google Scholar] [CrossRef]
- Martínez, L.; Andrade, R.; Birgin, E.G.; Martínez, J.M. Packmol: A package for building initial configurations for molecular dynamics simulations. J. Comput. Chem. 2009, 30, 2157–2164. [Google Scholar] [CrossRef] [PubMed]
- Salomon-Ferrer, R.; Case, D.A.; Walker, R.C. An overview of the Amber biomolecular simulation package. Wiley Interdiscip. Rev. Comput. Mol. Sci. 2012, 3, 198–210. [Google Scholar] [CrossRef]
- Tian, C.; Kasavajhala, K.; Belfon, K.A.A.; Raguette, L.; Huang, H.; Migues, A.N.; Bickel, J.; Wang, Y.; Pincay, J.; Wu, Q.; et al. ff19SB: Amino-Acid-Specific Protein Backbone Parameters Trained against Quantum Mechanics Energy Surfaces in Solution. J. Chem. Theory Comput. 2020, 16, 528–552. [Google Scholar] [CrossRef]
- Biller, J.R.; Elajaili, H.; Meyer, V.; Rosen, G.M.; Eaton, S.S.; Eaton, G.R. The Amber biomolecular simulation programs. J. Magn. Reson. 2013, 236, 47–56. [Google Scholar] [CrossRef]
- Giorgino, T. Computing diffusion coefficients in macromolecular simulations: The Diffusion Coefficient Tool for VMD. J. Open Source Softw. 2019, 4, 1698. [Google Scholar] [CrossRef]
- Piggot, T.J.; Allison, J.R.; Sessions, R.B.; Essex, J.W. On the Calculation of Acyl Chain Order Parameters from Lipid Simulations. J. Chem. Theory Comput. 2017, 13, 5683–5696. [Google Scholar] [CrossRef] [PubMed]
- Gurtovenko, A.A.; Miettinen, M.; Karttunen, M.; Vattulainen, I. Effect of Monovalent Salt on Cationic Lipid Membranes As Revealed by Molecular Dynamics Simulations. J. Phys. Chem. B 2005, 109, 21126–21134. [Google Scholar] [CrossRef] [PubMed]
- Romo, T.D.; Grossfield, A. LOOS: An extensible platform for the structural analysis of simulations. In Proceedings of the 2009 Annual International Conference of the IEEE Engineering in Medicine and Biology Society, Minneapolis, MN, USA, 3–6 September 2009; pp. 2332–2335. [Google Scholar] [CrossRef]
- Namani, T.; Walde, P. From Decanoate Micelles to Decanoic Acid/Dodecylbenzenesulfonate Vesicles. Langmuir 2005, 21, 6210–6219. [Google Scholar] [CrossRef] [PubMed]
- Kamp, F.; Zakim, D.; Zhang, F.; Noy, N.; Hamilton, J. Fatty Acid Flip-Flop in Phospholipid Bilayers Is Extremely Fast. Biochemistry 1995, 34, 11928–11937. [Google Scholar] [CrossRef]
- Budin, I.; Bruckner, R.J.; Szostak, J.W. Formation of Protocell-like Vesicles in a Thermal Diffusion Column. J. Am. Chem. Soc. 2009, 131, 9628–9629. [Google Scholar] [CrossRef]
- Cornell, C.E.; Black, R.A.; Williams, J.A.; Lee, K.K.; Drobny, G.P.; Keller, S.L.; Xue, M.; Litz, H.E.; Ramsay, A.; Gordon, M.; et al. Prebiotic amino acids bind to and stabilize prebiotic fatty acid membranes. Proc. Natl. Acad. Sci. USA 2019, 116, 17239–17244. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample * | Temperature °C | Radius (nm) | Thickness (nm) | % Inner Leaflets DA Monomers | 3D Diffusion Constant ** | SASA (nm2) |
|---|---|---|---|---|---|---|
| DA-0 | 0 | 3.55 | 1.44 | 20.9 | 0.89 | 5230 |
| DA-10 | 10 | 3.51 | 1.51 | 22.3 | 0.98 | 5219 |
| DA-20 | 20 | 3.53 | 1.51 | 21.4 | 1.77 | 5523 |
| DA-30 | 30 | 3.56 | 1.55 | 22.5 | 1.75 | 5784 |
| DA-40 | 40 | 3.61 | 1.52 | 22.4 | 1.39 | 6173 |
| DA-50 | 50 | 3.51 | 1.42 | 25.6 | 1.42 | 5883 |
| DA-60 | 60 | 3.67 | 1.54 | 21.9 | 6802 | |
| DA-70 | 70 | 3.59 | 1.45 | 23.4 | 6747 | |
| DA-80 | 80 | 3.61 | 1.47 | 24.2 | 7051 | |
| DA-90 | 90 | 3.59 | 1.45 | 23.4 | 7119 | |
| DA-100 | 100 | 3.61 | 1.47 | 23.0 | 7473 | |
| DA-110 | 110 | 3.64 | 1.51 | 21.4 | 7942 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sepulveda, R.V.; Sbarbaro, C.; Opazo, M.C.; Duarte, Y.; González-Nilo, F.; Aguayo, D. Insights into Early Steps of Decanoic Acid Self-Assemblies under Prebiotic Temperatures Using Molecular Dynamics Simulations. Membranes 2023, 13, 469. https://doi.org/10.3390/membranes13050469
Sepulveda RV, Sbarbaro C, Opazo MC, Duarte Y, González-Nilo F, Aguayo D. Insights into Early Steps of Decanoic Acid Self-Assemblies under Prebiotic Temperatures Using Molecular Dynamics Simulations. Membranes. 2023; 13(5):469. https://doi.org/10.3390/membranes13050469
Chicago/Turabian StyleSepulveda, Romina V., Christopher Sbarbaro, Ma Cecilia Opazo, Yorley Duarte, Fernando González-Nilo, and Daniel Aguayo. 2023. "Insights into Early Steps of Decanoic Acid Self-Assemblies under Prebiotic Temperatures Using Molecular Dynamics Simulations" Membranes 13, no. 5: 469. https://doi.org/10.3390/membranes13050469
APA StyleSepulveda, R. V., Sbarbaro, C., Opazo, M. C., Duarte, Y., González-Nilo, F., & Aguayo, D. (2023). Insights into Early Steps of Decanoic Acid Self-Assemblies under Prebiotic Temperatures Using Molecular Dynamics Simulations. Membranes, 13(5), 469. https://doi.org/10.3390/membranes13050469