
Application of Immersed Membrane Bioreactor for Semi-Continuous Production of Polyhydroxyalkanoates from Organic Waste-Based Volatile Fatty Acids

 ,
,  , ,
, ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Microorganism and Preculture Preparation
2.2. Organic Residues-Derived VFAs
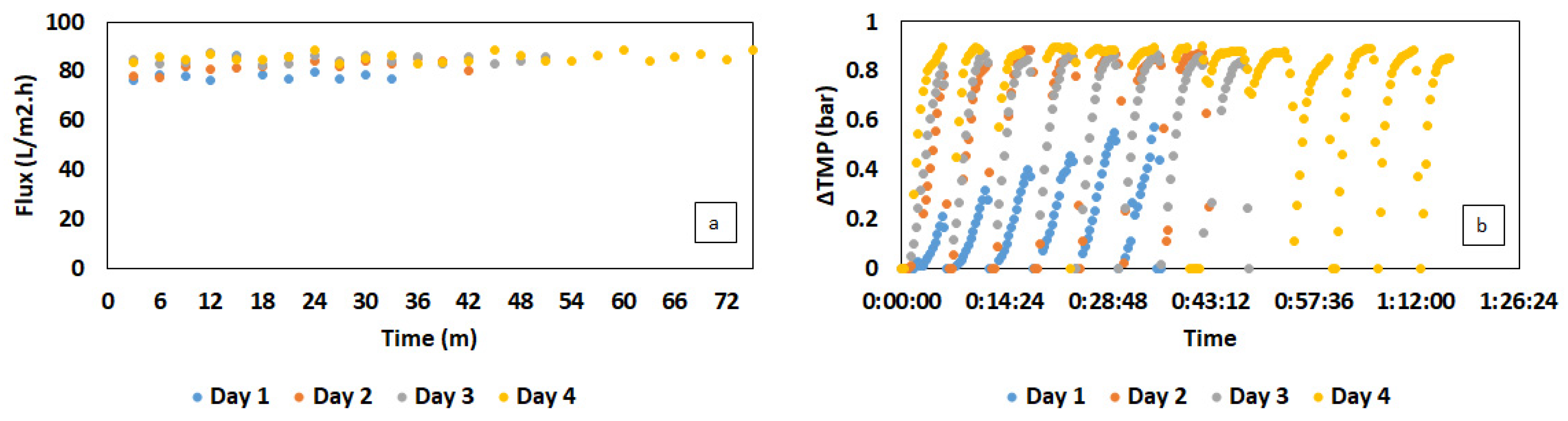
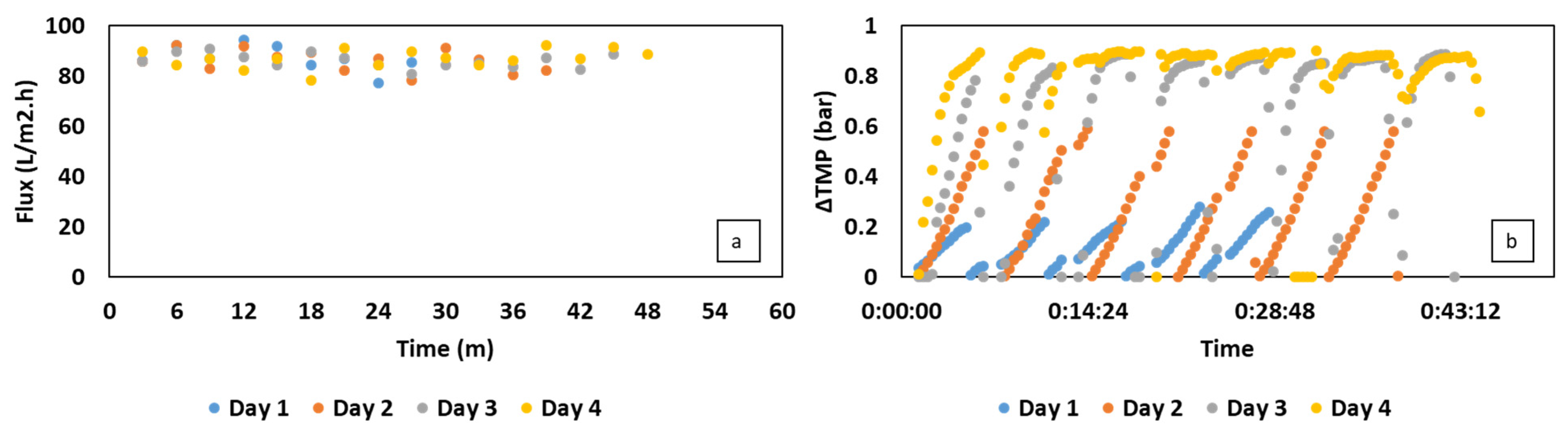
2.3. Immersed Membrane Bioreactor Setup
2.4. Semi-Continuous Cultivations
2.5. Analytical Methods
2.6. PHA Characterization
3. Results and Discussion
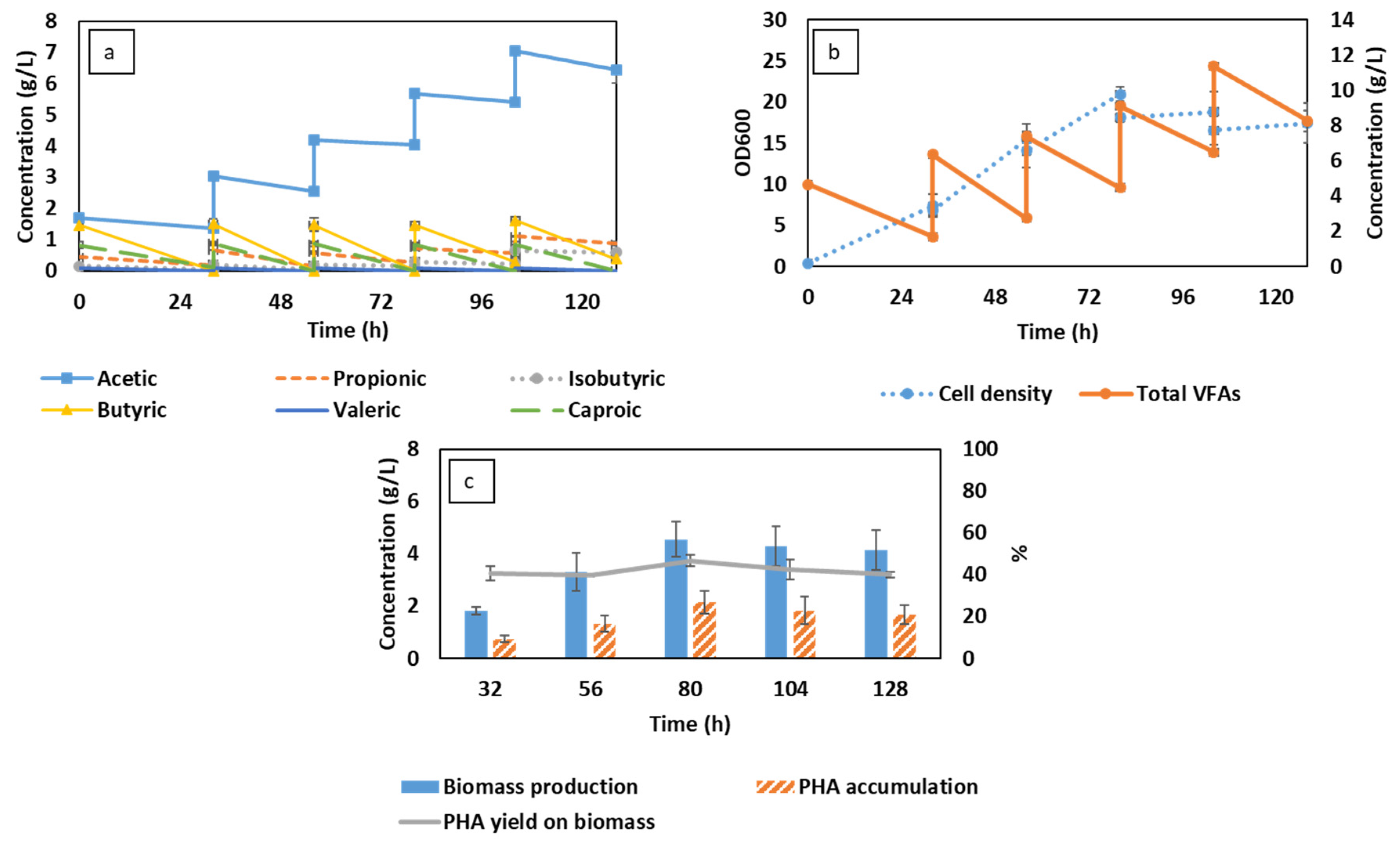
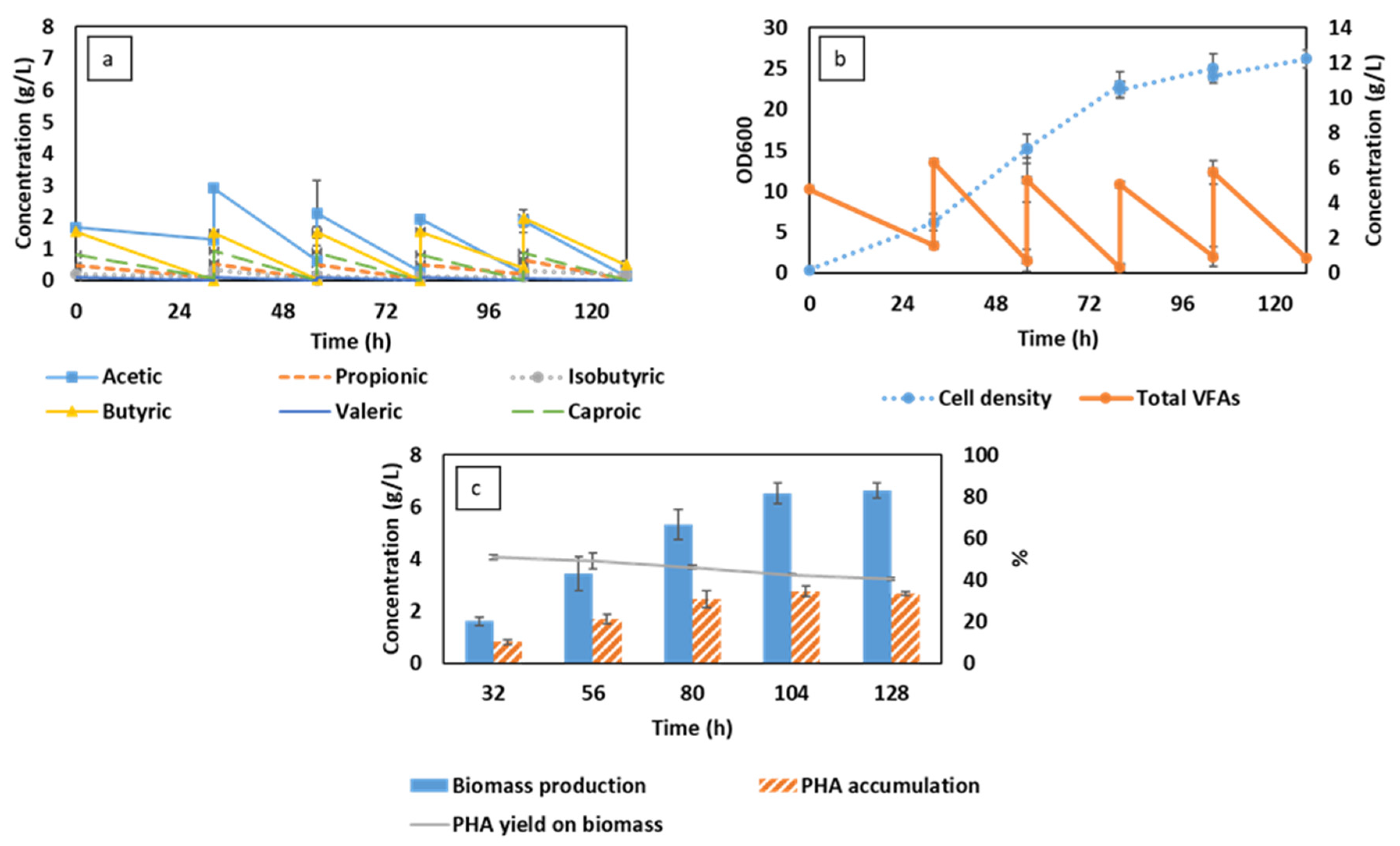
3.1. Semi-Continuous Cultivation of C. necator Cultivation Using Synthetic VFAs in CSTR
3.2. Semi-Continuous Cultivation of C. necator Using Synthetic VFAs in iMBR
3.3. C. necator Cultivation Using Organic Residues-Derived VFAs in iMBR
3.4. PHA Characterization
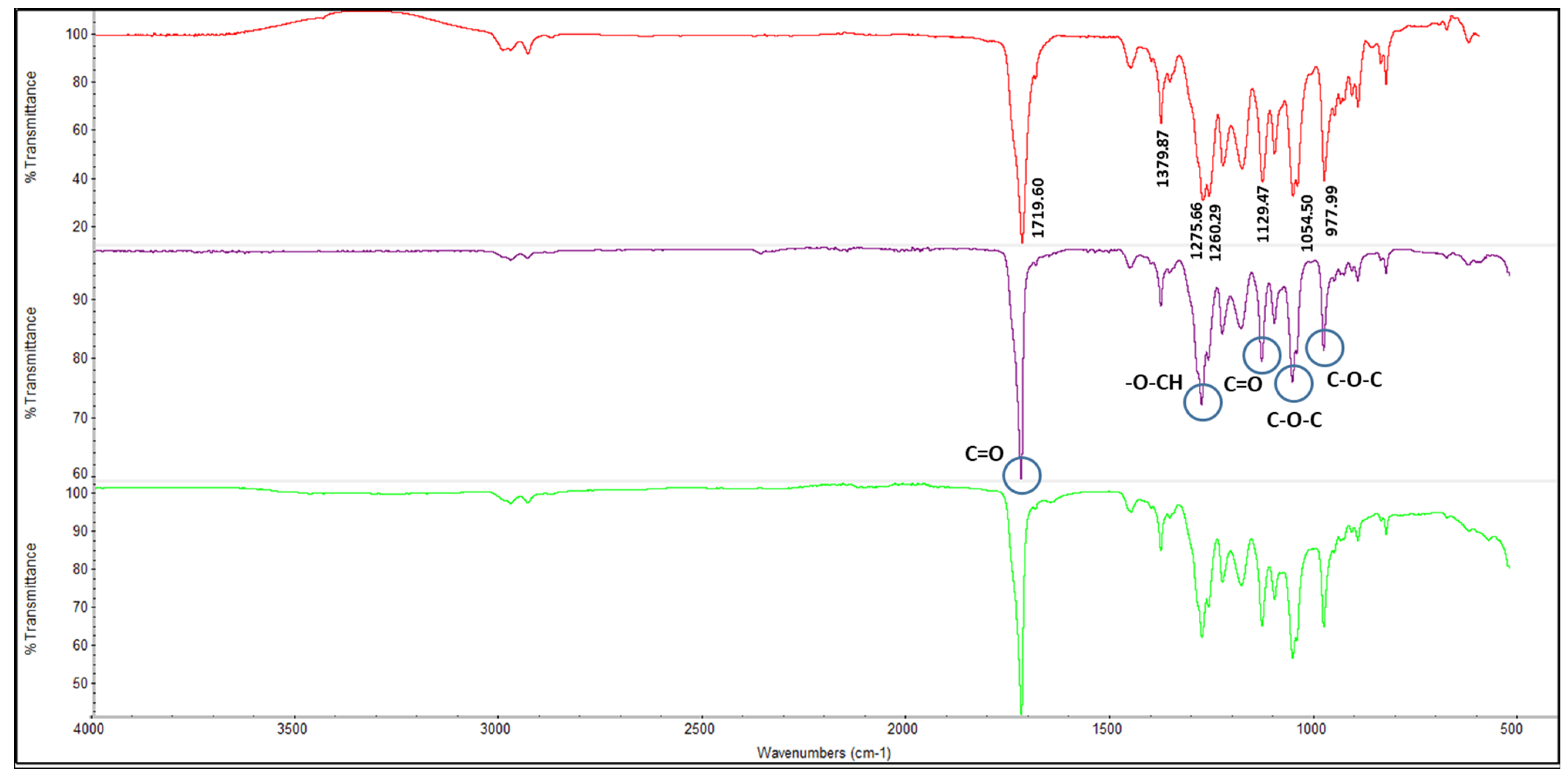
3.4.1. Fourier-Transform Infrared Spectroscopy (FTIR) Analysis
3.4.2. Thermogravimetric and Differential Scanning Calorimetry Analysis (TGA and DSC)
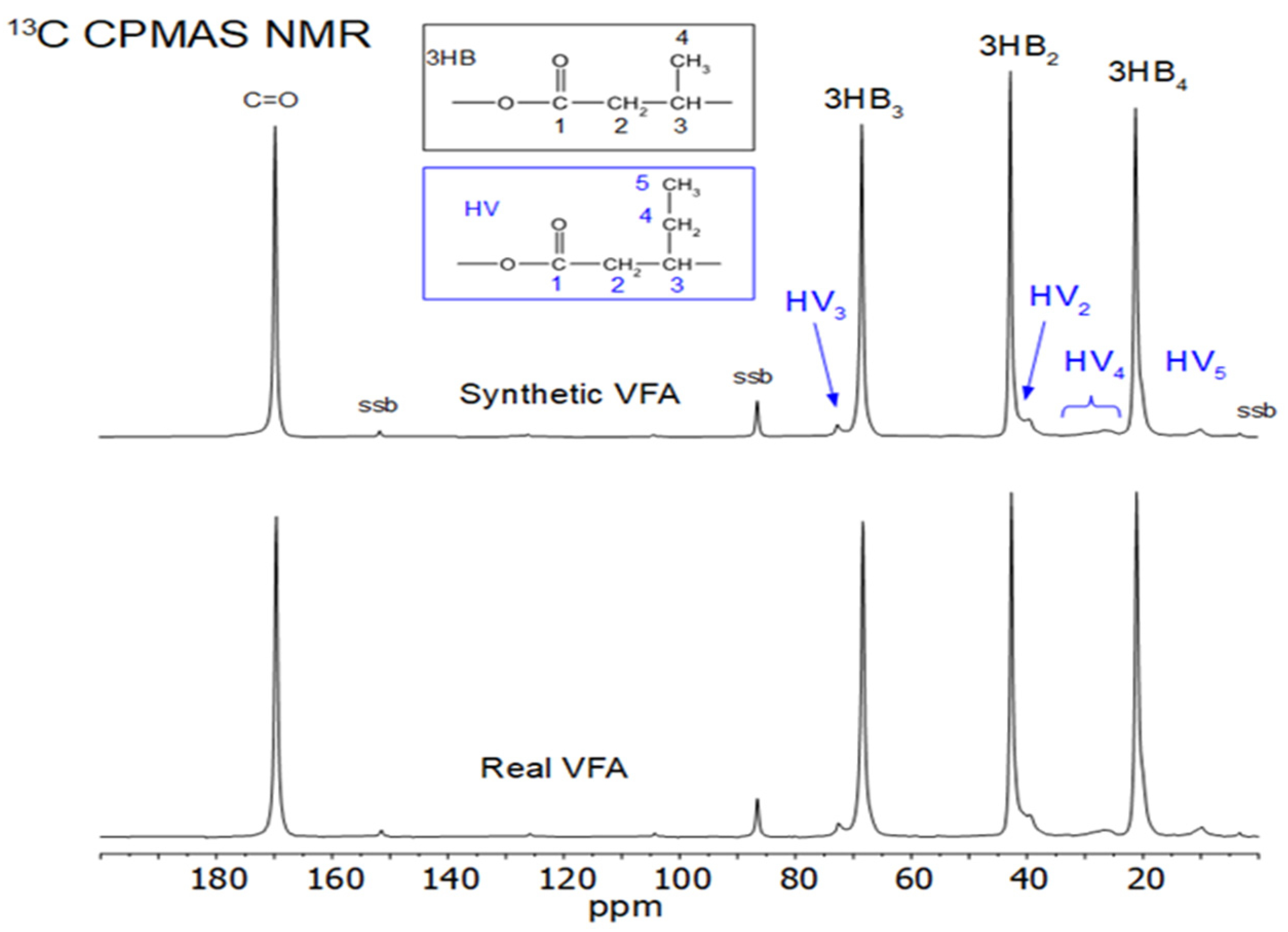
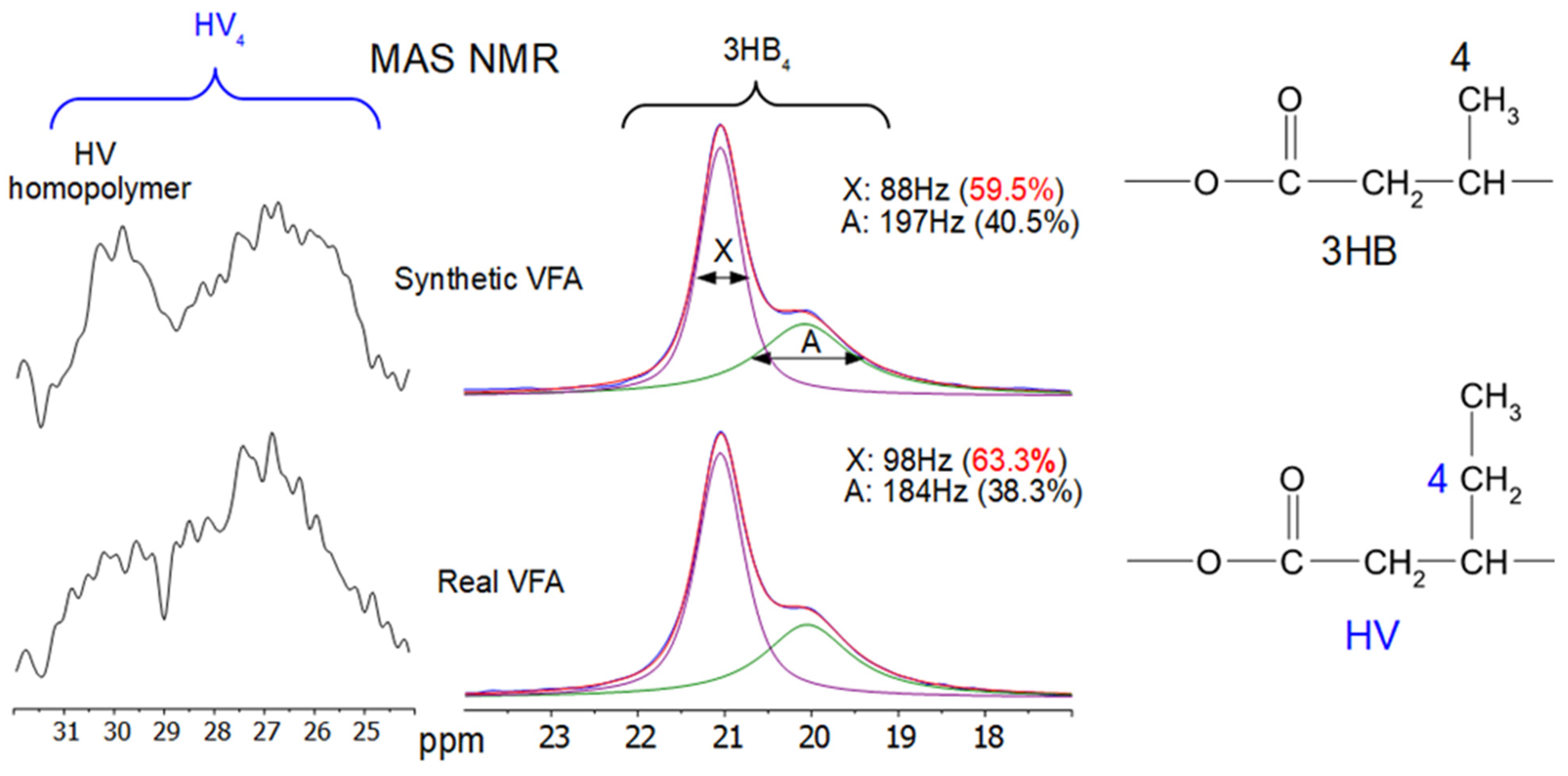
3.4.3. Nuclear Magnetic Resonance (NMR) Analysis
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Costa, S.S.; Miranda, A.L.; de Morais, M.G.; Costa, J.A.V.; Druzian, J.I. Microalgae as source of polyhydroxyalkanoates (PHAs) —A review. Int. J. Biol. Macromol. 2019, 131, 536–547. [Google Scholar] [CrossRef] [PubMed]
- Khok, Y.S.; Baidurah, S. Renewable biomass feedstocks for production of sustainable biodegradable polymer. Curr. Opin. Green Sustain. Chem. 2021, 27, 100412. [Google Scholar]
- Saratale, G.D.; Saratale, R.G.; Varjani, S.; Cho, S.-K.; Ghodake, G.S.; Kadam, A.; Mulla, S.I.; Bharagava, R.N.; Kim, D.-S.; Shin, H.S. Development of ultrasound aided chemical pretreatment methods to enrich saccharification of wheat waste biomass for polyhydroxybutyrate production and its characterization. Ind. Crops Prod. 2020, 150, 112425. [Google Scholar] [CrossRef]
- Chong, J.W.R.; Yew, G.Y.; Khoo, K.S.; Ho, S.-H.; Show, P.L. Recent advances on food waste pretreatment technology via microalgae for source of polyhydroxyalkanoates. J. Environ. Manag. 2021, 293, 112782. [Google Scholar] [CrossRef] [PubMed]
- Solaiman, D.K.; Ashby, R.D.; Foglia, T.A.; Marmer, W.N. Conversion of agricultural feedstock and coproducts into poly(hydroxyalkanoates). Appl. Microbiol. Biotechnol. 2006, 71, 783–789. [Google Scholar] [CrossRef]
- Saratale, R.G.; Cho, S.-K.; Saratale, G.D.; Kumar, M.; Bharagava, R.N.; Varjani, S.; Kadam, A.A.; Ghodake, G.S.; Palem, R.R.; Mulla, S.I.; et al. An Overview of Recent Advancements in Microbial Polyhydroxyalkanoates (PHA) Production from Dark Fermentation Acidogenic Effluents: A Path to an Integrated Bio-Refinery. Polymers 2021, 13, 4297. [Google Scholar] [CrossRef]
- Wainaina, S.; Parchami, M.; Mahboubi, A.; Horváth, I.S.; Taherzadeh, M.J. Food waste-derived volatile fatty acids platform using an immersed membrane bioreactor. Bioresour. Technol. 2019, 274, 329–334. [Google Scholar] [CrossRef]
- Agnihotri, S.; Yin, D.M.; Mahboubi, A.; Sapmaz, T.; Varjani, S.; Qiao, W.; Koseoglu-Imer, D.Y.; Taherzadeh, M.J. A Glimpse of the World of Volatile Fatty Acids Production and Application: A review. Bioengineered 2022, 13, 1249–1275. [Google Scholar] [CrossRef]
- Ramos-Suarez, M.; Zhang, Y.; Outram, V. Current perspectives on acidogenic fermentation to produce volatile fatty acids from waste. Rev. Environ. Sci. Bio/Technol. 2021, 20, 439–478. [Google Scholar] [CrossRef]
- Wainaina, S.; Horváth, I.S.; Taherzadeh, M.J. Biochemicals from food waste and recalcitrant biomass via syngas fermentation: A review. Bioresour. Technol. 2018, 248, 113–121. [Google Scholar] [CrossRef]
- Szacherska, K.; Oleskowicz-Popiel, P.; Ciesielski, S.; Mozejko-Ciesielska, J. Volatile Fatty Acids as Carbon Sources for Polyhydroxyalkanoates Production. Polymers 2021, 13, 321. [Google Scholar] [CrossRef]
- Gutschmann, B.; Huang, B.; Santolin, L.; Thiele, I.; Neubauer, P.; Riedel, S.L. Native feedstock options for the polyhydroxyalkanoate industry in Europe: A review. Microbiol. Res. 2022, 264, 127177. [Google Scholar] [CrossRef]
- Perez-Zabaleta, M.; Atasoy, M.; Khatami, K.; Eriksson, E.; Cetecioglu, Z. Bio-based conversion of volatile fatty acids from waste streams to polyhydroxyalkanoates using mixed microbial cultures. Bioresour. Technol. 2021, 323, 124604. [Google Scholar] [CrossRef]
- Du, G.; Si, Y.; Yu, J. Inhibitory effect of medium-chain-length fatty acids on synthesis of polyhydroxyalkanoates from volatile fatty acids by Ralstonia eutropha. Biotechnol. Lett. 2001, 23, 1613–1617. [Google Scholar] [CrossRef]
- López, J.C.; Arnáiz, E.; Merchán, L.; Lebrero, R.; Muñoz, R. Biogas-based polyhydroxyalkanoates production by Methylocystis hirsuta: A step further in anaerobic digestion biorefineries. Chem. Eng. J. 2018, 333, 529–536. [Google Scholar] [CrossRef]
- Vishnuvardhan Reddy, S.; Thirumala, M.; Mahmood, S.K. Production of PHB and P (3HB-co-3HV) biopolymers by Bacillus megaterium strain OU303A isolated from municipal sewage sludge. World J. Microbiol. Biotechnol. 2009, 25, 391–397. [Google Scholar] [CrossRef]
- Yun, J.; Sawant, S.; Kim, B.S. Production of Polyhydroxyalkanoates by Ralstonia Eutropha from Volatile Fatty Acids. Korean J. Chem. Eng. 2013, 30, 2223–2227. [Google Scholar] [CrossRef]
- Liu, S. Chapter 11—How Cells Grow. In Bioprocess Engineering, 2nd ed.; Liu, S., Ed.; Elsevier: Amsterdam, The Netherlands, 2017; pp. 629–697. [Google Scholar]
- Zeb, B.S.; Mahmood, Q.; Ping, Z.; Lin, Q.; Lu, H.; Tingting, C.; Abbas, G. Assessment of toxicity of volatile fatty acids to Photobacterium phosphoreum. Microbiology 2014, 83, 510–515. [Google Scholar] [CrossRef]
- Agustín Martinez, G.; Bertin, L.; Scoma, A.; Rebecchi, S.; Braunegg, G.; Fava, F. Production of polyhydroxyalkanoates from dephenolised and fermented olive mill wastewaters by employing a pure culture of Cupriavidus necator. Biochem. Eng. J. 2015, 97, 92–100. [Google Scholar] [CrossRef]
- Quinlan, A.V. A semicontinuous culture model that links cell growth to extracellular nutrient concentration. Biotechnol. Bioeng. 1986, 28, 1455–1461. [Google Scholar] [CrossRef]
- Westman, J.O.; Franzén, C.J. Current progress in high cell density yeast bioprocesses for bioethanol production. Biotechnol. J. 2015, 10, 1185–1195. [Google Scholar] [CrossRef] [PubMed]
- Carstensen, F.; Apel, A.; Wessling, M. In situ product recovery: Submerged membranes vs. external loop membranes. J. Membr. Sci. 2012, 394–395, 1–36. [Google Scholar] [CrossRef]
- Mahboubi Soufiani, A.; Uwineza, C.; Doyen, W.; De Wever, H.; Taherzadeh, M. Intensification of lignocellulosic bioethanol production process using continuous double-staged immersed membrane bioreactors. Bioresour. Technol. 2019, 296, 122314. [Google Scholar] [CrossRef]
- Rahman, T.U.; Roy, H.; Islam, M.R.; Tahmid, M.; Fariha, A.; Mazumder, A.; Tasnim, N.; Pervez, M.N.; Cai, Y.; Naddeo, V.; et al. The Advancement in Membrane Bioreactor (MBR) Technology toward Sustainable Industrial Wastewater Management. Membranes 2023, 13, 181. [Google Scholar] [CrossRef]
- Wainaina, S.; Awasthi, M.K.; Horváth, I.S.; Taherzadeh, M.J. Anaerobic digestion of food waste to volatile fatty acids and hydrogen at high organic loading rates in immersed membrane bioreactors. Renew. Energy 2020, 152, 1140–1148. [Google Scholar] [CrossRef]
- Mahboubi, A.; Ylitervo, P.; Doyen, W.; De Wever, H.; Molenberghs, B.; Taherzadeh, M.J. Continuous bioethanol fermentation from wheat straw hydrolysate with high suspended solid content using an immersed flat sheet membrane bioreactor. Bioresour. Technol. 2017, 241, 296–308. [Google Scholar] [CrossRef] [PubMed]
- Vu, D.H.; Wainaina, S.; Taherzadeh, M.J.; Åkesson, D.; Ferreira, J.A. Production of polyhydroxyalkanoates (PHAs) by Bacillus megaterium using food waste acidogenic fermentation-derived volatile fatty acids. Bioengineered 2021, 12, 2480–2498. [Google Scholar] [CrossRef] [PubMed]
- Policastro, G.; Panico, A.; Fabbricino, M. Improving biological production of poly(3-hydroxybutyrate-co-3-hydroxyvalerate) (PHBV) co-polymer: A critical review. Rev. Environ. Sci. Bio/Technol. 2021, 20, 479–513. [Google Scholar] [CrossRef]
- Raza, Z.A.; Tariq, M.R.; Majeed, M.I.; Banat, I.M. Recent developments in bioreactor scale production of bacterial polyhydroxyalkanoates. Bioprocess Biosyst. Eng. 2019, 42, 901–919. [Google Scholar] [CrossRef]
- Vu, D.H.; Mahboubi, A.; Root, A.; Heinmaa, I.; Taherzadeh, M.J.; Åkesson, D. Thorough Investigation of the Effects of Cultivation Factors on Polyhydroalkanoates (PHAs) Production by Cupriavidus necator from Food Waste-Derived Volatile Fatty Acids. Fermentation 2022, 8, 605. [Google Scholar] [CrossRef]
- Kovárová-Kovar, K.; Egli, T. Growth kinetics of suspended microbial cells: From single-substrate-controlled growth to mixed-substrate kinetics. Microbiol. Mol. Biol. Rev. 1998, 62, 646–666. [Google Scholar] [CrossRef] [PubMed]
- Setiadi, T.; Aznury, M.; Trianto, A.; Pancoro, A. Production of polyhydroxyalkanoate (PHA) by Ralstonia eutropha JMP 134 with volatile fatty acids from palm oil mill effluent as precursors. Water Sci. Technol. 2015, 72, 1889–1895. [Google Scholar] [CrossRef] [PubMed]
- Gaveau, A.; Coetsier, C.; Roques, C.; Bacchin, P.; Dague, E.; Causserand, C. Bacteria transfer by deformation through microfiltration membrane. J. Membr. Sci. 2017, 523, 446–455. [Google Scholar] [CrossRef]
- Pervez, M.N.; Bilgiç, B.; Mahboubi, A.; Uwineza, C.; Zarra, T.; Belgiorno, V.; Naddeo, V.; Taherzadeh, M.J. Double-stage membrane-assisted anaerobic digestion process intensification for production and recovery of volatile fatty acids from food waste. Sci. Total Environ. 2022, 825, 154084. [Google Scholar] [CrossRef]
- Iorhemen, O.T.; Hamza, R.A.; Tay, J.H. Membrane Bioreactor (MBR) Technology for Wastewater Treatment and Reclamation: Membrane Fouling. Membranes 2016, 6, 33. [Google Scholar] [CrossRef]
- He, Z.; Miller, D.; Kasemset, S.; Paul, D.; Freeman, B. The Effect of Permeate Flux on Membrane Fouling during Microfiltration of Oily Water. J. Membr. Sci. 2016, 525, 25–34. [Google Scholar] [CrossRef]
- Khomlaem, C.; Aloui, H.; Singhvi, M.; Kim, B.S. Production of polyhydroxyalkanoates and astaxanthin from lignocellulosic biomass in high cell density membrane bioreactor. Chem. Eng. J. 2023, 451, 138641. [Google Scholar] [CrossRef]
- Aloui, H.; Khomlaem, C.; Torres, C.A.V.; Freitas, F.; Reis, M.A.M.; Kim, B.S. Enhanced co-production of medium-chain-length polyhydroxyalkanoates and phenazines from crude glycerol by high cell density cultivation of Pseudomonas chlororaphis in membrane bioreactor. Int. J. Biol. Macromol. 2022, 211, 545–555. [Google Scholar] [CrossRef]
- Chakraborty, P.; Gibbons, W.; Muthukumarappan, K. Conversion of volatile fatty acids into polyhydroxyalkanoate by Ralstonia eutropha. J. Appl. Microbiol. 2009, 106, 1996–2005. [Google Scholar] [CrossRef]
- Jaramillo-Sánchez, R.; Alcaraz-Zapata, W. Limitations on production methods for PHAs obtention: A systematic review. Dyna 2020, 87, 193–203. [Google Scholar]
- Khumwanich, P.; Napathorn, S.; Suwannasilp, B. Polyhydroxyalkanoate Production with a Feast/Famine Feeding Regime Using Sludge from Wastewater Treatment Plants of the Food and Beverage Industry. J. Biobased Mater. Bioenergy 2014, 8, 641–647. [Google Scholar]
- Meneses, L.; Craveiro, R.; Jesus, A.R.; Reis, M.A.M.; Freitas, F.; Paiva, A. Supercritical CO2 Assisted Impregnation of Ibuprofen on Medium-Chain-Length Polyhydroxyalkanoates (mcl-PHA). Molecules 2021, 26, 4772. [Google Scholar] [CrossRef]
- Muneer, F.; Rasul, I.; Qasim, M.; Sajid, A.; Nadeem, H. Optimization, Production and Characterization of Polyhydroxyalkanoate (PHA) from Indigenously Isolated Novel Bacteria. J. Polym. Environ. 2022, 30, 3523–3533. [Google Scholar] [CrossRef]
- Khomlaem, C.; Aloui, H.; Deshmukh, A.R.; Yun, J.-H.; Kim, H.-S.; Napathorn, S.C.; Kim, B.S. Defatted Chlorella biomass as a renewable carbon source for polyhydroxyalkanoates and carotenoids co-production. Algal Res. 2020, 51, 102068. [Google Scholar] [CrossRef]
- Khomlaem, C.; Aloui, H.; Kim, B.S. Biosynthesis of Polyhydroxyalkanoates from Defatted Chlorella Biomass as an Inexpensive Substrate. Appl. Sci. 2021, 11, 1094. [Google Scholar] [CrossRef]
- Khomlaem, C.; Aloui, H.; Oh, W.-G.; Kim, B.S. High cell density culture of Paracoccus sp. LL1 in membrane bioreactor for enhanced co-production of polyhydroxyalkanoates and astaxanthin. Int. J. Biol. Macromol. 2021, 192, 289–297. [Google Scholar]
- Nygaard, D.; Yashchuk, O.; Noseda, D.G.; Araoz, B.; Hermida, É.B. Improved fermentation strategies in a bioreactor for enhancing poly(3-hydroxybutyrate) (PHB) production by wild type Cupriavidus necator from fructose. Heliyon 2021, 7, e05979. [Google Scholar] [CrossRef]
- Ntaikou, I.; Koumelis, I.; Kamilari, M.; Iatridi, Z.; Tsitsilianis, C.; Lyberatos, G. Effect of nitrogen limitation on polyhydroxyalkanoates production efficiency, properties and microbial dynamics using a soil-derived mixed continuous culture. Int. J. Biobased Plast. 2019, 1, 31–47. [Google Scholar] [CrossRef]
- Rosengart, A.; Cesário, M.T.; de Almeida, M.C.M.D.; Raposo, R.S.; Espert, A.; de Apodaca, E.D.; da Fonseca, M.M.R. Efficient P(3HB) extraction from Burkholderia sacchari cells using non-chlorinated solvents. Biochem. Eng. J. 2015, 103, 39–46. [Google Scholar] [CrossRef]
- VanderHart, D.L.; Orts, W.J.; Marchessault, R.H. 13C NMR Determination of the Degree of Cocrystallization in Random Copolymers of Poly(.beta.-hydroxybutyrate-co-.beta.-hydroxyvalerate). Macromolecules 1995, 28, 6394–6400. [Google Scholar]
- Kamiya, N.; Sakurai, M.; Inoue, Y.; Chujo, R.; Doi, Y. Study of cocrystallization of poly(3-hydroxybutyrate-co-3-hydroxyvalerate) by solid-state high-resolution carbon-13 NMR spectroscopy and differential scanning calorimetry. Macromolecules 1991, 24, 2178–2182. [Google Scholar] [CrossRef]
- Nieuwendaal, R.C.; Snyder, C.R.; DeLongchamp, D.M. Measuring Order in Regioregular Poly(3-hexylthiophene) with Solid-State 13C CPMAS NMR. ACS Macro Lett. 2014, 3, 130–135. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Components | Concentration (g/L) |
|---|---|
| Acetic acid | 3.01 |
| Propionic acid | 0.80 |
| Butyric acid | 4.15 |
| Isobutyric acid | 0.07 |
| Valeric acid | 0.13 |
| Isovaleric acid | 0.00 |
| Caproic acid | 0.65 |
| Ammonium | 0.84 |
| Total VFAs | 8.81 |
| Samples | Tonset (°C) | Tmax (°C) | Total Mass Loss (%) | Tm (°C) | Melting Enthalpy (J/g) | Degree of Crystallinity (%) |
|---|---|---|---|---|---|---|
| Synthetic VFAs | 246.8.6 ± 1.8 | 297.6 ± 2.0 | 98.1 | 146.2 ± 0.5 | 34.5 ± 1.3 | 23.8 |
| Organic waste-based VFA effluent | 253.16 ± 1.2 | 302.3 ± 0.8 | 89.1 | 143.4 ± 0.9 | 13.8 ± 0.8 | 9.6 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vu, D.H.; Mahboubi, A.; Root, A.; Heinmaa, I.; Taherzadeh, M.J.; Åkesson, D. Application of Immersed Membrane Bioreactor for Semi-Continuous Production of Polyhydroxyalkanoates from Organic Waste-Based Volatile Fatty Acids. Membranes 2023, 13, 569. https://doi.org/10.3390/membranes13060569
Vu DH, Mahboubi A, Root A, Heinmaa I, Taherzadeh MJ, Åkesson D. Application of Immersed Membrane Bioreactor for Semi-Continuous Production of Polyhydroxyalkanoates from Organic Waste-Based Volatile Fatty Acids. Membranes. 2023; 13(6):569. https://doi.org/10.3390/membranes13060569
Chicago/Turabian StyleVu, Danh H., Amir Mahboubi, Andrew Root, Ivo Heinmaa, Mohammad J. Taherzadeh, and Dan Åkesson. 2023. "Application of Immersed Membrane Bioreactor for Semi-Continuous Production of Polyhydroxyalkanoates from Organic Waste-Based Volatile Fatty Acids" Membranes 13, no. 6: 569. https://doi.org/10.3390/membranes13060569
APA StyleVu, D. H., Mahboubi, A., Root, A., Heinmaa, I., Taherzadeh, M. J., & Åkesson, D. (2023). Application of Immersed Membrane Bioreactor for Semi-Continuous Production of Polyhydroxyalkanoates from Organic Waste-Based Volatile Fatty Acids. Membranes, 13(6), 569. https://doi.org/10.3390/membranes13060569