

Membranes and Synaptosomes Used to Investigate Synaptic GABAergic Currents in Epileptic Patients

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Patients
2.2. Membranes and Synaptosomes’ Preparation and Intracellular Injection
2.3. Electrophysiological Recordings
2.4. Statistical Analysis
3. Results
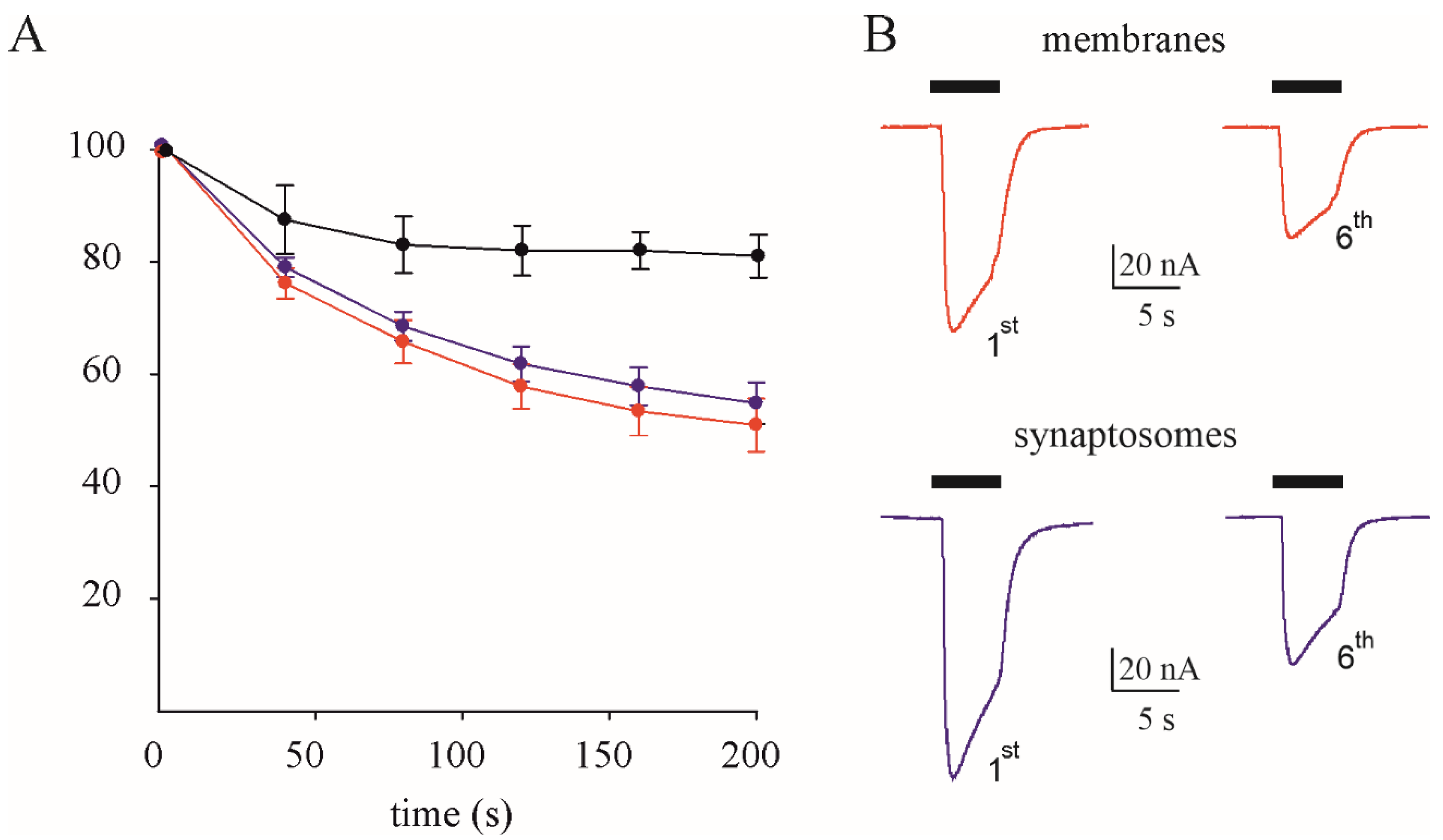
3.1. GABAA Current Rundown
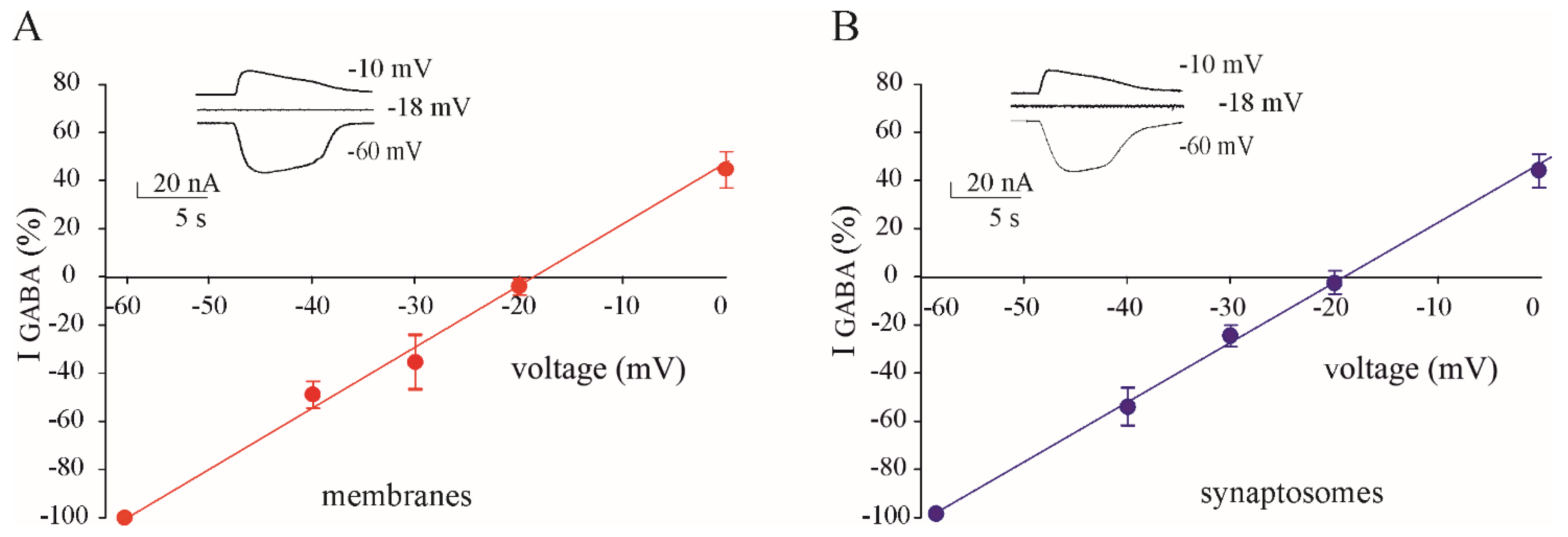
3.2. GABA Reversal Potential in Oocytes Injected with Membranes and Synaptosomes
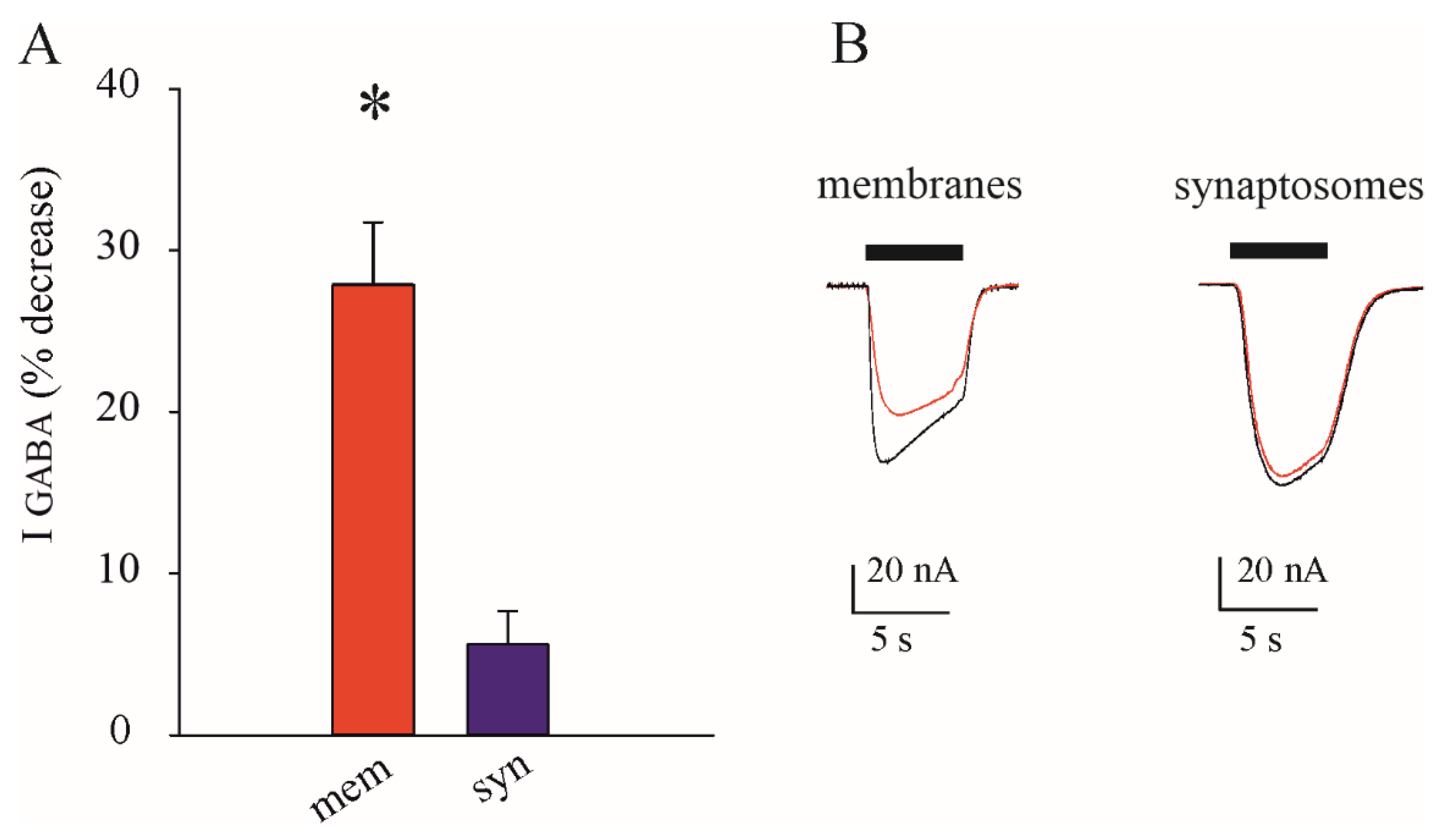
3.3. Lack of Inhibition of GABAARs in Synaptosomes after Zn2+ Application
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Thijs, R.D.; Surges, R.; O’Brien, T.J.; Sander, J.W. Epilepsy in Adults. Lancet 2019, 393, 689–701. [Google Scholar] [CrossRef] [PubMed]
- Fattorusso, A.; Matricardi, S.; Mencaroni, E.; Dell’Isola, G.B.; Di Cara, G.; Striano, P.; Verrotti, A. The Pharmacoresistant Epilepsy: An Overview on Existant and New Emerging Therapies. Front. Neurol. 2021, 12, 674483. [Google Scholar] [CrossRef] [PubMed]
- Ghit, A.; Assal, D.; Al-Shami, A.S.; Hussein, D.E.E. GABAA Receptors: Structure, Function, Pharmacology, and Related Disorders. J. Genet. Eng. Biotechnol. 2021, 19, 123. [Google Scholar] [CrossRef] [PubMed]
- Sallard, E.; Letourneur, D.; Legendre, P. Electrophysiology of Ionotropic GABA Receptors. Cell. Mol. Life Sci. 2021, 78, 5341–5370. [Google Scholar] [CrossRef] [PubMed]
- Farrant, M.; Nusser, Z. Variations on an Inhibitory Theme: Phasic and Tonic Activation of GABAA Receptors. Nat. Rev. Neurosci. 2005, 6, 215–229. [Google Scholar] [CrossRef] [PubMed]
- Markwardt, S.; Overstreet-Wadiche, L. GABAergic Signalling to Adult-generated Neurons. J. Physiol. 2008, 586, 3745–3749. [Google Scholar] [CrossRef]
- Rahmati, N.; Hoebeek, F.E.; Peter, S.; De Zeeuw, C.I. Chloride Homeostasis in Neurons With Special Emphasis on the Olivocerebellar System: Differential Roles for Transporters and Channels. Front. Cell. Neurosci. 2018, 12, 101. [Google Scholar] [CrossRef]
- Mortensen, M.; Smart, T.G. Extrasynaptic Aβ Subunit GABAA Receptors on Rat Hippocampal Pyramidal Neurons. J. Physiol. 2006, 577, 841–856. [Google Scholar] [CrossRef]
- Prenosil, G.A. Specific Subtypes of GABAA Receptors Mediate Phasic and Tonic Forms of Inhibition in Hippocampal Pyramidal Neurons. J. Neurophysiol. 2006, 96, 846–857. [Google Scholar] [CrossRef]
- Fritschy, J.-M. E/I Balance and GABAA Receptor Plasticity. Front. Mol. Neurosci. 2008, 1, 5. [Google Scholar] [CrossRef]
- Song, I.; Savtchenko, L.; Semyanov, A. Tonic Excitation or Inhibition Is Set by GABAA Conductance in Hippocampal Interneurons. Nat. Commun. 2011, 2, 376. [Google Scholar] [CrossRef]
- Kaila, K.; Price, T.J.; Payne, J.A.; Puskarjov, M.; Voipio, J. Cation-Chloride Cotransporters in Neuronal Development, Plasticity and Disease. Nat. Rev. Neurosci. 2014, 15, 637–654. [Google Scholar] [CrossRef] [PubMed]
- Pavlov, I.; Walker, M.C. Tonic GABAA Receptor-Mediated Signalling in Temporal Lobe Epilepsy. Neuropharmacology 2013, 69, 55–61. [Google Scholar] [CrossRef] [PubMed]
- Koh, W.; Kwak, H.; Cheong, E.; Lee, C.J. GABA Tone Regulation and Its Cognitive Functions in the Brain. Nat. Rev. Neurosci. 2023, 24, 523–539. [Google Scholar] [CrossRef] [PubMed]
- Thomas, P.; Mortensen, M.; Hosie, A.M.; Smart, T.G. Dynamic Mobility of Functional GABAA Receptors at Inhibitory Synapses. Nat. Neurosci. 2005, 8, 889–897. [Google Scholar] [CrossRef] [PubMed]
- Hannan, S.; Minere, M.; Harris, J.; Izquierdo, P.; Thomas, P.; Tench, B.; Smart, T.G. GABAAR Isoform and Subunit Structural Motifs Determine Synaptic and Extrasynaptic Receptor Localisation. Neuropharmacology 2020, 169, 107540. [Google Scholar] [CrossRef] [PubMed]
- Palma, E.; Ragozzino, D.A.; Di Angelantonio, S.; Spinelli, G.; Trettel, F.; Martinez-Torres, A.; Torchia, G.; Arcella, A.; Di Gennaro, G.; Quarato, P.P.; et al. Phosphatase Inhibitors Remove the Run-down of γ-Aminobutyric Acid Type A Receptors in the Human Epileptic Brain. Proc. Natl. Acad. Sci. USA 2004, 101, 10183–10188. [Google Scholar] [CrossRef] [PubMed]
- Palma, E.; Roseti, C.; Maiolino, F.; Fucile, S.; Martinello, K.; Mazzuferi, M.; Aronica, E.; Manfredi, M.; Esposito, V.; Cantore, G.; et al. GABAA-Current Rundown of Temporal Lobe Epilepsy Is Associated with Repetitive Activation of GABAA “Phasic” Receptors. Proc. Natl. Acad. Sci. USA 2007, 104, 20944–20948. [Google Scholar] [CrossRef] [PubMed]
- Ruffolo, G.; Cifelli, P.; Roseti, C.; Thom, M.; Van Vliet, E.A.; Limatola, C.; Aronica, E.; Palma, E. A Novel GABAergic Dysfunction in Human Dravet Syndrome. Epilepsia 2018, 59, 2106–2117. [Google Scholar] [CrossRef]
- Palma, E.; Amici, M.; Sobrero, F.; Spinelli, G.; Di Angelantonio, S.; Ragozzino, D.; Mascia, A.; Scoppetta, C.; Esposito, V.; Miledi, R.; et al. Anomalous Levels of Cl− Transporters in the Hippocampal Subiculum from Temporal Lobe Epilepsy Patients Make GABA Excitatory. Proc. Natl. Acad. Sci. USA 2006, 103, 8465–8468. [Google Scholar] [CrossRef]
- Falk-Petersen, C.B.; Tsonkov, T.M.; Nielsen, M.S.; Harpsøe, K.; Bundgaard, C.; Frølund, B.; Kristiansen, U.; Gloriam, D.E.; Wellendorph, P. Discovery of a New Class of Orthosteric Antagonists with Nanomolar Potency at Extrasynaptic GABAA Receptors. Sci. Rep. 2020, 10, 10078. [Google Scholar] [CrossRef]
- Etherington, L.-A.; Mihalik, B.; Pálvölgyi, A.; Ling, I.; Pallagi, K.; Kertész, S.; Varga, P.; Gunn, B.G.; Brown, A.R.; Livesey, M.R.; et al. Selective Inhibition of Extra-Synaptic A5-GABAA Receptors by S44819, a New Therapeutic Agent. Neuropharmacology 2017, 125, 353–364. [Google Scholar] [CrossRef]
- Olsen, R.W. Allosteric Ligands and Their Binding Sites Define γ-Aminobutyric Acid (GABA) Type A Receptor Subtypes. Adv. Pharmacol. 2015, 73, 167–202. [Google Scholar] [CrossRef] [PubMed]
- Bai, F.; Witzmann, F.A. Synaptosome Proteomics. In Subcellular Proteomics; Bertrand, E., Faupel, M., Eds.; Subcellular Biochemistry; Springer: Dordrecht, The Netherlands, 2007; Volume 43, pp. 77–98. ISBN 978-1-4020-5942-1. [Google Scholar]
- Gulyássy, P.; Puska, G.; Györffy, B.A.; Todorov-Völgyi, K.; Juhász, G.; Drahos, L.; Kékesi, K.A. Proteomic Comparison of Different Synaptosome Preparation Procedures. Amino Acids 2020, 52, 1529–1543. [Google Scholar] [CrossRef] [PubMed]
- Evans, G.J.O. The Synaptosome as a Model System for Studying Synaptic Physiology. Cold Spring Harb. Protoc. 2015, 2015, pdb.top074450. [Google Scholar] [CrossRef] [PubMed]
- Michaelis, M.L.; Jiang, L.; Michaelis, E.K. Isolation of Synaptosomes, Synaptic Plasma Membranes, and Synaptic Junctional Complexes. In Synapse Development; Methods in Molecular Biology; Poulopoulos, A., Ed.; Springer: New York, NY, USA, 2017; Volume 1538, pp. 107–119. ISBN 978-1-4939-6686-8. [Google Scholar]
- Ragozzino, D.; Palma, E.; Di Angelantonio, S.; Amici, M.; Mascia, A.; Arcella, A.; Giangaspero, F.; Cantore, G.; Di Gennaro, G.; Manfredi, M.; et al. Rundown of GABA Type A Receptors Is a Dysfunction Associated with Human Drug-Resistant Mesial Temporal Lobe Epilepsy. Proc. Natl. Acad. Sci. USA 2005, 102, 15219–15223. [Google Scholar] [CrossRef] [PubMed]
- Palma, E.; Torchia, G.; Limatola, C.; Trettel, F.; Arcella, A.; Cantore, G.; Di Gennaro, G.; Manfredi, M.; Esposito, V.; Quarato, P.P.; et al. BDNF Modulates GABAA Receptors Microtransplanted from the Human Epileptic Brain to Xenopus Oocytes. Proc. Natl. Acad. Sci. USA 2005, 102, 1667–1672. [Google Scholar] [CrossRef] [PubMed]
- Stell, B.M.; Mody, I. Receptors with Different Affinities Mediate Phasic and Tonic GABAA Conductances in Hippocampal Neurons. J. Neurosci. 2002, 22, RC223. [Google Scholar] [CrossRef] [PubMed]
- Semyanov, A.; Walker, M.C.; Kullmann, D.M.; Silver, R.A. Tonically Active GABAA Receptors: Modulating Gain and Maintaining the Tone. Trends Neurosci. 2004, 27, 262–269. [Google Scholar] [CrossRef] [PubMed]
- Ben-Ari, Y.; Khalilov, I.; Kahle, K.T.; Cherubini, E. The GABA Excitatory/Inhibitory Shift in Brain Maturation and Neurological Disorders. Neuroscientist 2012, 18, 467–486. [Google Scholar] [CrossRef]
- Liu, R.; Wang, J.; Liang, S.; Zhang, G.; Yang, X. Role of NKCC1 and KCC2 in Epilepsy: From Expression to Function. Front. Neurol. 2020, 10, 1407. [Google Scholar] [CrossRef] [PubMed]
- Hartmann, A.-M.; Nothwang, H.G. NKCC1 and KCC2: Structural Insights into Phospho-Regulation. Front. Mol. Neurosci. 2022, 15, 964488. [Google Scholar] [CrossRef]
- Braat, S.; Kooy, R.F. The GABAA Receptor as a Therapeutic Target for Neurodevelopmental Disorders. Neuron 2015, 86, 1119–1130. [Google Scholar] [CrossRef] [PubMed]
- Cohen, I.; Navarro, V.; Clemenceau, S.; Baulac, M.; Miles, R. On the Origin of Interictal Activity in Human Temporal Lobe Epilepsy In Vitro. Science 2002, 298, 1418–1421. [Google Scholar] [CrossRef]
- Pallud, J.; Le Van Quyen, M.; Bielle, F.; Pellegrino, C.; Varlet, P.; Cresto, N.; Baulac, M.; Duyckaerts, C.; Kourdougli, N.; Chazal, G.; et al. Cortical GABAergic Excitation Contributes to Epileptic Activities around Human Glioma. Sci. Transl. Med. 2014, 6, 244ra89. [Google Scholar] [CrossRef]
- Lauterborn, J.C.; Scaduto, P.; Cox, C.D.; Schulmann, A.; Lynch, G.; Gall, C.M.; Keene, C.D.; Limon, A. Increased Excitatory to Inhibitory Synaptic Ratio in Parietal Cortex Samples from Individuals with Alzheimer’s Disease. Nat. Commun. 2021, 12, 2603. [Google Scholar] [CrossRef]
- Hosie, A.M.; Dunne, E.L.; Harvey, R.J.; Smart, T.G. Zinc-Mediated Inhibition of GABAA Receptors: Discrete Binding Sites Underlie Subtype Specificity. Nat. Neurosci. 2003, 6, 362–369. [Google Scholar] [CrossRef]
- Ruffolo, G.; Cifelli, P.; Miranda-Lourenço, C.; De Felice, E.; Limatola, C.; Sebastião, A.M.; Diógenes, M.J.; Aronica, E.; Palma, E. Rare Diseases of Neurodevelopment: Maintain the Mystery or Use a Dazzling Tool for Investigation? The Case of Rett Syndrome. Neuroscience 2020, 439, 146–152. [Google Scholar] [CrossRef]
- Palma, E.; Ragozzino, D.; Angelantonio, S.D.; Mascia, A.; Maiolino, F.; Manfredi, M.; Cantore, G.; Esposito, V.; Di Gennaro, G.; Quarato, P.; et al. The Antiepileptic Drug Levetiracetam Stabilizes the Human Epileptic GABAA Receptors upon Repetitive Activation. Epilepsia 2007, 48, 1842–1849. [Google Scholar] [CrossRef] [PubMed]
- Moss, S.J.; Gorrie, G.H.; Amato, A.; Smart, T.G. Modulation of GABAA Receptors by Tyrosine Phosphorylation. Nature 1995, 377, 344–348. [Google Scholar] [CrossRef] [PubMed]
- Houston, C.M.; Lee, H.H.C.; Hosie, A.M.; Moss, S.J.; Smart, T.G. Identification of the Sites for CaMK-II-Dependent Phosphorylation of GABAA Receptors. J. Biol. Chem. 2007, 282, 17855–17865. [Google Scholar] [CrossRef] [PubMed]
- Ben-Ari, Y.; Cherubini, E. The GABA Polarity Shift and Bumetanide Treatment: Making Sense Requires Unbiased and Undogmatic Analysis. Cells 2022, 11, 396. [Google Scholar] [CrossRef] [PubMed]
- Talos, D.M.; Sun, H.; Kosaras, B.; Joseph, A.; Folkerth, R.D.; Poduri, A.; Madsen, J.R.; Black, P.M.; Jensen, F.E. Altered Inhibition in Tuberous Sclerosis and Type IIb Cortical Dysplasia. Ann. Neurol. 2012, 71, 539–551. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Patient | Age (y)/Gender | Epilepsy Onset (y) | Seizure Types | Pathology | Surgical Zone | ASMs |
|---|---|---|---|---|---|---|
| #1 | 27/M | 10 | FIAS | HS | Right temporal | CBZ; TPM |
| #2 | 26/M | 6 | FIAS | HS | Left temporal | LEV; LMT; TPM |
| #3 | 28/F | 14 | FAS | HS | Left temporal | CBZ; LMT |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gaeta, A.; Lissner, L.J.; Alfano, V.; Cifelli, P.; Morano, A.; Roseti, C.; Di Iacovo, A.; Aronica, E.; Palma, E.; Ruffolo, G. Membranes and Synaptosomes Used to Investigate Synaptic GABAergic Currents in Epileptic Patients. Membranes 2024, 14, 64. https://doi.org/10.3390/membranes14030064
Gaeta A, Lissner LJ, Alfano V, Cifelli P, Morano A, Roseti C, Di Iacovo A, Aronica E, Palma E, Ruffolo G. Membranes and Synaptosomes Used to Investigate Synaptic GABAergic Currents in Epileptic Patients. Membranes. 2024; 14(3):64. https://doi.org/10.3390/membranes14030064
Chicago/Turabian StyleGaeta, Alessandro, Lilian Juliana Lissner, Veronica Alfano, Pierangelo Cifelli, Alessandra Morano, Cristina Roseti, Angela Di Iacovo, Eleonora Aronica, Eleonora Palma, and Gabriele Ruffolo. 2024. "Membranes and Synaptosomes Used to Investigate Synaptic GABAergic Currents in Epileptic Patients" Membranes 14, no. 3: 64. https://doi.org/10.3390/membranes14030064
APA StyleGaeta, A., Lissner, L. J., Alfano, V., Cifelli, P., Morano, A., Roseti, C., Di Iacovo, A., Aronica, E., Palma, E., & Ruffolo, G. (2024). Membranes and Synaptosomes Used to Investigate Synaptic GABAergic Currents in Epileptic Patients. Membranes, 14(3), 64. https://doi.org/10.3390/membranes14030064