
Increased Serum Levels of Activated Caspases in Murine and Human Biliary Atresia

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental BA Mouse Model
2.2. RRV and Real-Time PCR
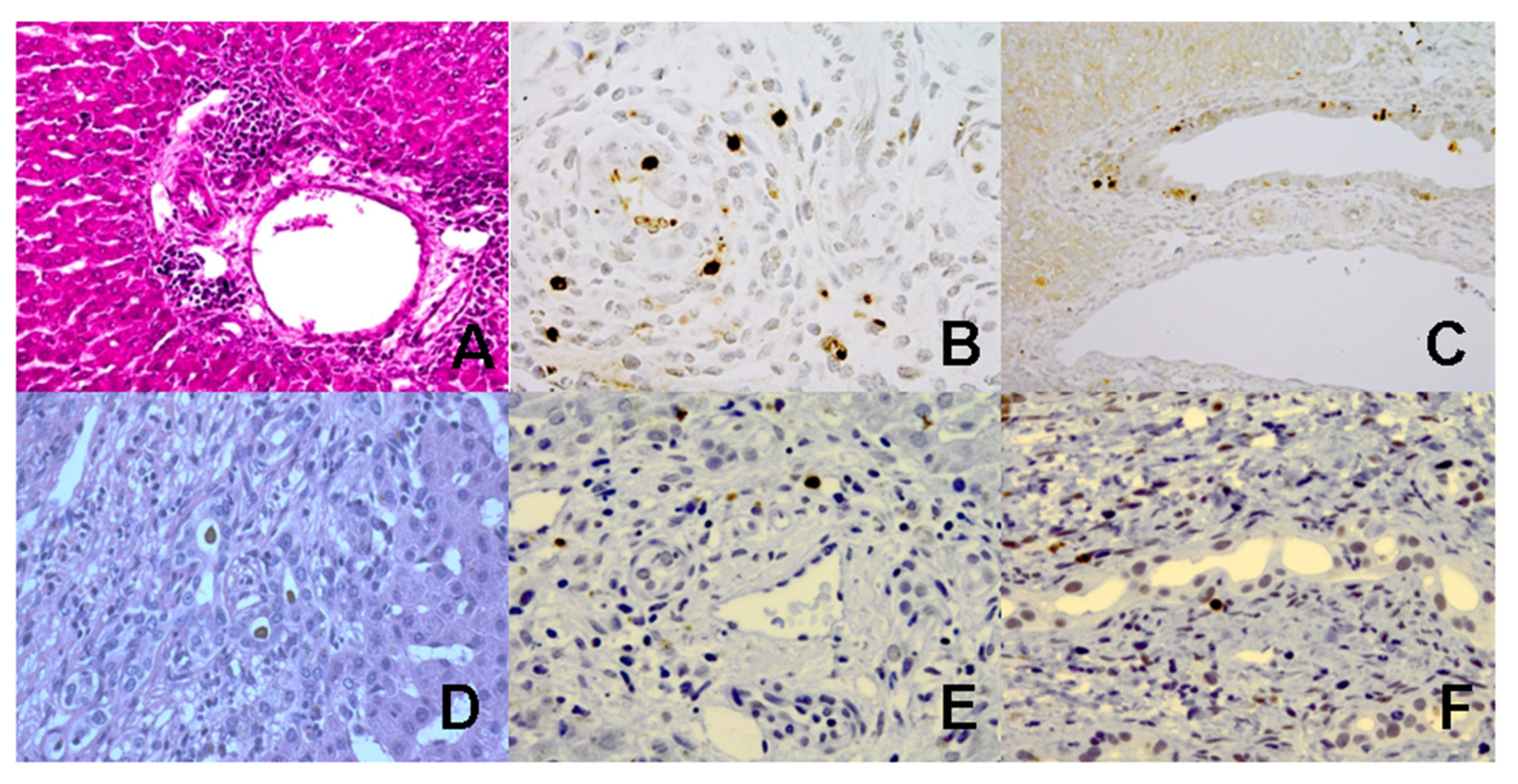
2.3. Immunohistochemical Staining
2.4. Detection of Caspase Activity in Mouse Liver Tissue
2.5. Detection of Caspase Activity in Mouse Sera
2.6. Detection of Caspase Activity in Human Sera
2.7. Patients Characteristics
2.8. Statistics
3. Results
3.1. Induction of BA and Cholestasis in RRV-Infected Mice
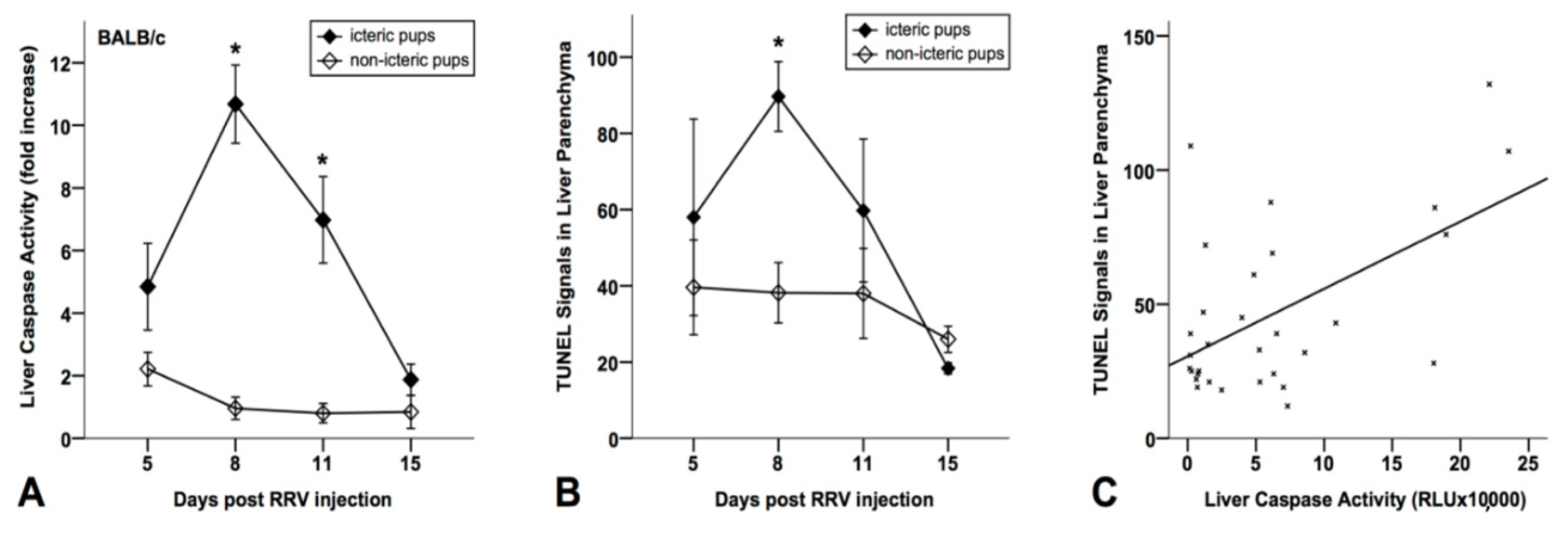
3.2. Increased Caspase Activation and Apoptosis in Liver Tissues from BA Mice
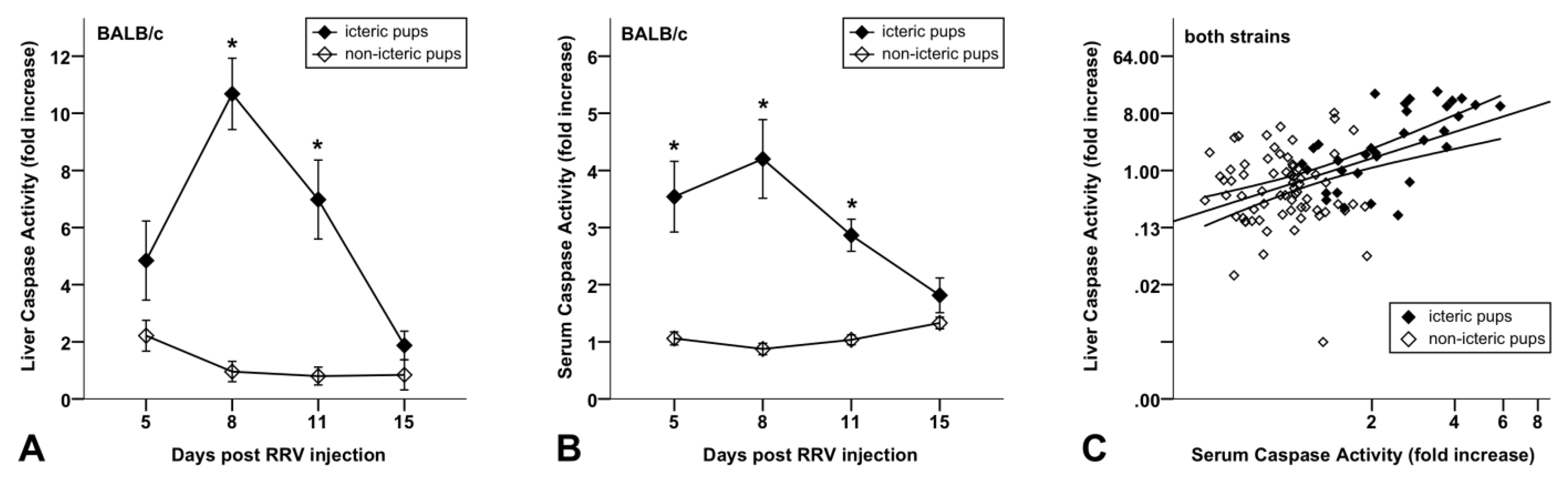
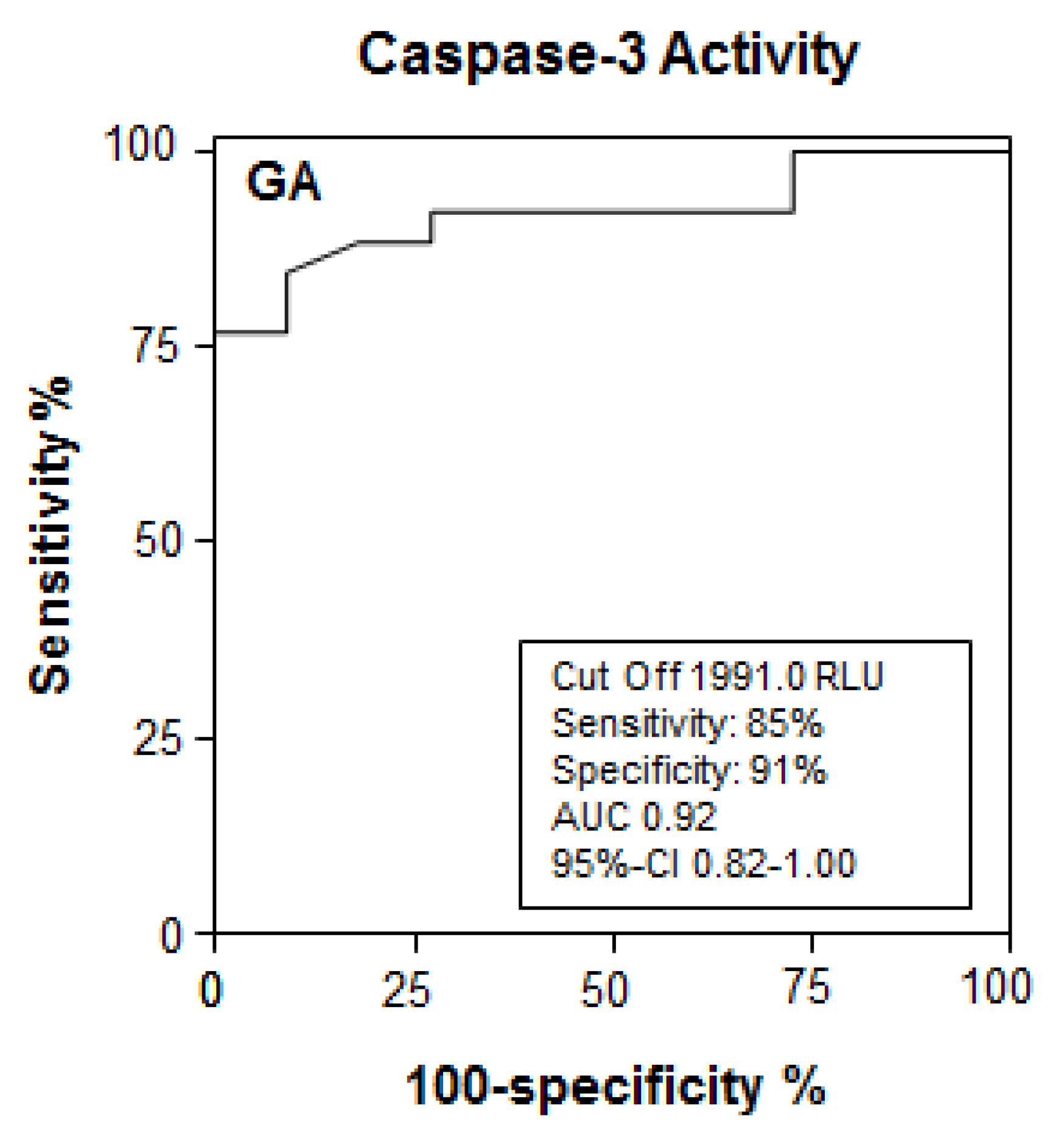
3.3. Increased Caspase Activity in Sera of BA Mice Compared to Non-Icteric Mice
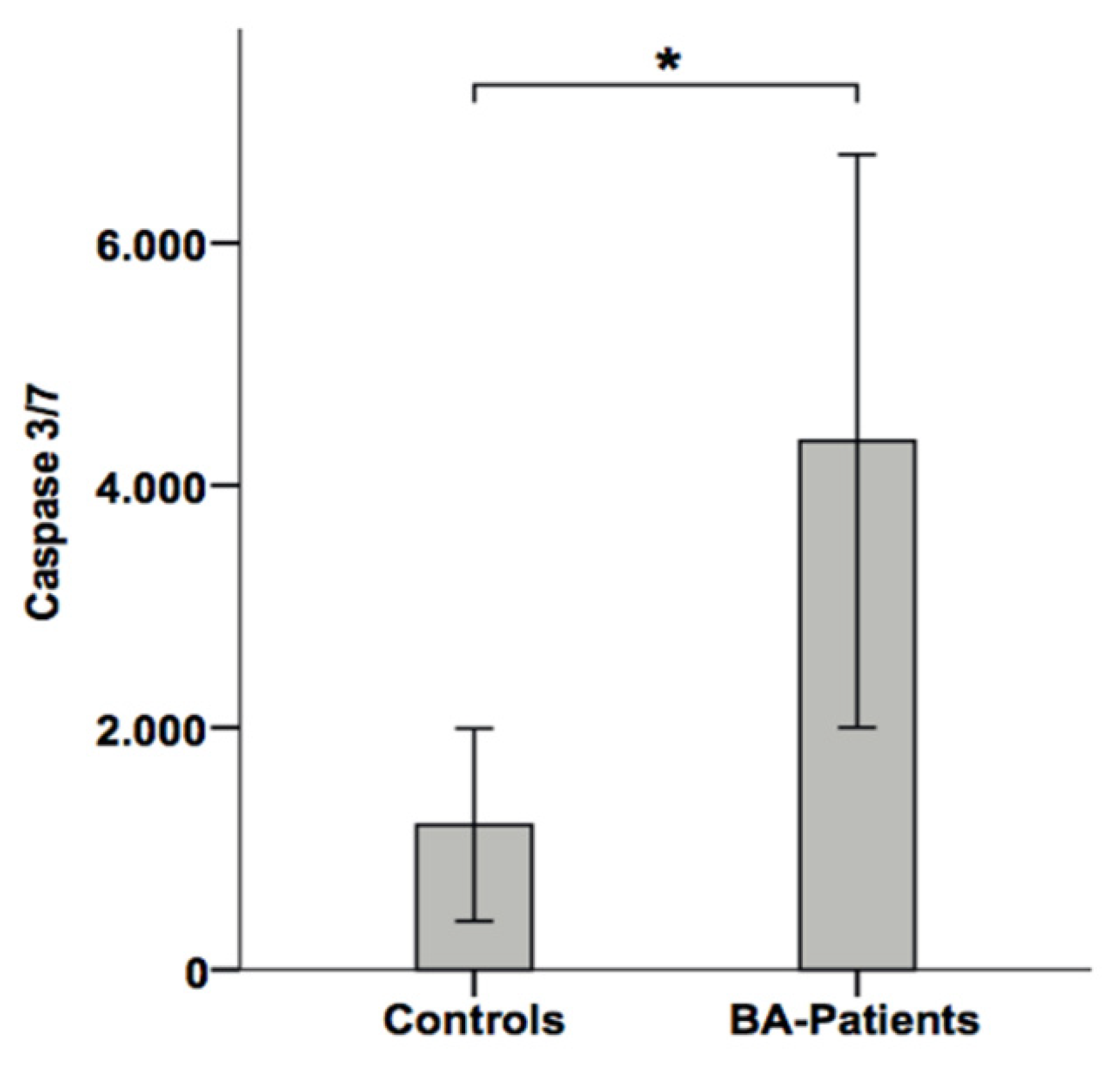
3.4. Apoptotic Liver Injury Is Increased in BA Patients and Associated with Elevated Caspase Activity in Serum
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
Abbreviations
| BA | Biliary Atresia |
| LTx | Liver Transplantation |
| RRV | Rhesus Rotavirus |
| CK | Cytokeratin |
| IFNγ | Interferon-Gamma |
| TNFα | Tumor Necrosis Factor-alpha |
| Pfu | Plaque Forming Units |
| ELISA | Enzyme-Linked Immunosorbent Assay |
| H.E. | Hematoxylin and Eosin |
| PCR | Polymerase Chain Reaction |
| GAPDH | Glyceraldehyde-3-Phosphate-Dehydrogenase |
| TUNEL | Terminal Deoxynucleotidyl Transferase-mediated Deoxyuridine Triphosphated Nick End-Labelling |
| FFPE | Formalin-Fixed, Paraffin Embedded |
| SEM | Standard Error of Mean |
| DAB | Diaminobenzidin |
| RLU | Relative Light Units |
| CTL | Cytotoxic T Cell |
References
- Chardot, C.; Buet, C.; Serinet, M.-O.; Golmard, J.-L.; Lachaux, A.; Roquelaure, B.; Gottrand, F.; Broué, P.; Dabadie, A.; Gauthier, F.; et al. Improving Outcomes of Biliary Atresia: French National Series 1986–2009. J. Hepatol. 2013, 58, 1209–1217. [Google Scholar] [CrossRef]
- Davenport, M.; Ong, E.; Sharif, K.; Alizai, N.; McClean, P.; Hadzic, N.; Kelly, D.A. Biliary Atresia in England and Wales: Results of Centralization and New Benchmark. J. Pediatr. Surg. 2011, 46, 1689–1694. [Google Scholar] [CrossRef] [PubMed]
- Nio, M.; Wada, M.; Sasaki, H.; Tanaka, H. Effects of Age at Kasai Portoenterostomy on the Surgical Outcome: A Review of the Literature. Surg. Today 2015, 45, 813–818. [Google Scholar] [CrossRef]
- Davenport, M. Biliary Atresia: Clinical Aspects. Semin. Pediatr. Surg. 2012, 21, 175–184. [Google Scholar] [CrossRef]
- Petersen, C. Pathogenesis and Treatment Opportunities for Biliary Atresia. Clin. Liver Dis. 2006, 10, 73–88. [Google Scholar] [CrossRef]
- Lien, T.-H.; Chang, M.-H.; Wu, J.-F.; Chen, H.-L.; Lee, H.-C.; Chen, A.-C.; Tiao, M.-M.; Wu, T.-C.; Yang, Y.-J.; Lin, C.-C.; et al. Effects of the Infant Stool Color Card Screening Program on 5-Year Outcome of Biliary Atresia in Taiwan. Hepatology 2011, 53, 202–208. [Google Scholar] [CrossRef] [PubMed]
- Shneider, B.L. Screening for Biliary Atresia: A Ray of Hope. Hepatology 2008, 47, 1105–1107. [Google Scholar] [CrossRef] [PubMed]
- Sokol, R.J. Biliary Atresia Screening: Why, When, and How? Pediatrics 2009, 123, e951–e952. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sokol, R.J.; Mack, C.L. Optimizing Outcomes and Bridging Biliary Atresia into Adulthood. Hepatology 2005, 41, 231–233. [Google Scholar] [CrossRef]
- Petersen, C. Biliary Atresia: The Animal Models. Semin. Pediatr. Surg. 2012, 21, 185–191. [Google Scholar] [CrossRef]
- Bezerra, J.A. Biliary Atresia—Translational Research on Key Molecular Processes Regulating Biliary Injury and Obstruction. Chang Gung Med. J. 2006, 29, 222–230. [Google Scholar] [PubMed]
- Mack, C.L. The Pathogenesis of Biliary Atresia: Evidence for a Virus-Induced Autoimmune Disease. Semin. Liver Dis. 2007, 27, 233–242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharland, A.; Gorrell, M.D. Cooperation of Innate and Adaptive Immunity in the Pathogenesis of Biliary Atresia: There’s a Killer on the Run. Hepatology 2009, 50, 2037–2040. [Google Scholar] [CrossRef]
- Malhi, H.; Gores, G.J. Cellular and Molecular Mechanisms of Liver Injury. Gastroenterology 2008, 134, 1641–1654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schulze-Osthoff, K.; Ferrari, D.; Los, M.; Wesselborg, S.; Peter, M.E. Apoptosis Signaling by Death Receptors. Eur. J. Biochem. 1998, 254, 439–459. [Google Scholar] [CrossRef]
- Fischer, U.; Jänicke, R.U.; Schulze-Osthoff, K. Many Cuts to Ruin: A Comprehensive Update of Caspase Substrates. Cell Death Differ. 2003, 10, 76–100. [Google Scholar] [CrossRef]
- Leers, M.P.; Kölgen, W.; Björklund, V.; Bergman, T.; Tribbick, G.; Persson, B.; Björklund, P.; Ramaekers, F.C.; Björklund, B.; Nap, M.; et al. Immunocytochemical Detection and Mapping of a Cytokeratin 18 Neo-Epitope Exposed during Early Apoptosis. J. Pathol. 1999, 187, 567–572. [Google Scholar] [CrossRef]
- Canbay, A.; Higuchi, H.; Bronk, S.F.; Taniai, M.; Sebo, T.J.; Gores, G.J. Fas Enhances Fibrogenesis in the Bile Duct Ligated Mouse: A Link between Apoptosis and Fibrosis. Gastroenterology 2002, 123, 1323–1330. [Google Scholar] [CrossRef]
- Funaki, N.; Sasano, H.; Shizawa, S.; Nio, M.; Iwami, D.; Ohi, R.; Nagura, H. Apoptosis and Cell Proliferation in Biliary Atresia. J. Pathol. 1998, 186, 429–433. [Google Scholar] [CrossRef]
- Liu, C.; Chiu, J.H.; Chin, T.; Wang, L.S.; Li, A.F.; Chow, K.C.; Wei, C. Expression of Fas Ligand on Bile Ductule Epithelium in Biliary Atresia—A Poor Prognostic Factor. J. Pediatr. Surg. 2000, 35, 1591–1596. [Google Scholar] [CrossRef]
- Carvalho, E.; Liu, C.; Shivakumar, P.; Sabla, G.; Aronow, B.; Bezerra, J.A. Analysis of the Biliary Transcriptome in Experimental Biliary Atresia. Gastroenterology 2005, 129, 713–717. [Google Scholar] [CrossRef] [PubMed]
- Leonhardt, J.; Stanulla, M.; von Wasielewski, R.; Skokowa, J.; Kübler, J.; Ure, B.M.; Petersen, C. Gene Expression Profile of the Infective Murine Model for Biliary Atresia. Pediatr. Surg. Int. 2006, 22, 84–89. [Google Scholar] [CrossRef]
- Erickson, N.; Mohanty, S.K.; Shivakumar, P.; Sabla, G.; Chakraborty, R.; Bezerra, J.A. Temporal-Spatial Activation of Apoptosis and Epithelial Injury in Murine Experimental Biliary Atresia. Hepatology 2008, 47, 1567–1577. [Google Scholar] [CrossRef]
- Leonhardt, J.; Kuebler, J.F.; Turowski, C.; Tschernig, T.; Geffers, R.; Petersen, C. Susceptibility to Experimental Biliary Atresia Linked to Different Hepatic Gene Expression Profiles in Two Mouse Strains. Hepatol. Res. Off. J. Jpn. Soc. Hepatol. 2010, 40, 196–203. [Google Scholar] [CrossRef]
- Petersen, C.; Grasshoff, S.; Luciano, L. Diverse Morphology of Biliary Atresia in an Animal Model. J. Hepatol. 1998, 28, 603–607. [Google Scholar] [CrossRef]
- Volkmann, X.; Fischer, U.; Bahr, M.J.; Ott, M.; Lehner, F.; Macfarlane, M.; Cohen, G.M.; Manns, M.P.; Schulze-Osthoff, K.; Bantel, H. Increased Hepatotoxicity of Tumor Necrosis Factor-Related Apoptosis-Inducing Ligand in Diseased Human Liver. Hepatology 2007, 46, 1498–1508. [Google Scholar] [CrossRef] [PubMed]
- Petersen, C.; Davenport, M. Aetiology of Biliary Atresia: What Is Actually Known? Orphanet J. Rare Dis. 2013, 8, 128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anthony, S.J.; Annunziato, R.A.; Fairey, E.; Kelly, V.L.; So, S.; Wray, J. Waiting for Transplant: Physical, Psychosocial, and Nutritional Status Considerations for Pediatric Candidates and Implications for Care. Pediatr. Transplant. 2014, 18, 423–434. [Google Scholar] [CrossRef]
- Mack, C.L.; Feldman, A.G.; Sokol, R.J. Clues to the Etiology of Bile Duct Injury in Biliary Atresia. Semin. Liver Dis. 2012, 32, 307–316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmed, A.F.; Ohtani, H.; Nio, M.; Funaki, N.; Shimaoka, S.; Nagura, H.; Ohi, R. CD8+ T Cells Infiltrating into Bile Ducts in Biliary Atresia Do Not Appear to Function as Cytotoxic T Cells: A Clinicopathological Analysis. J. Pathol. 2001, 193, 383–389. [Google Scholar] [CrossRef]
- Clarke, P.; Tyler, K.L. Apoptosis in Animal Models of Virus-Induced Disease. Nat. Rev. Microbiol. 2009, 7, 144–155. [Google Scholar] [CrossRef] [Green Version]
- Mohanty, S.K.; Lobeck, I.; Donnelly, B.; Dupree, P.; Walther, A.; Mowery, S.; Coots, A.; Bondoc, A.; Sheridan, R.M.; Poling, H.M.; et al. Rotavirus Reassortant—Induced Murine Model of Liver Fibrosis Parallels Human Biliary Atresia. Hepatology 2020, 71, 1316–1330. [Google Scholar] [CrossRef]
- Keyzer-Dekker, C.M.G.; Lind, R.C.; Kuebler, J.F.; Offerhaus, G.J.A.; ten Kate, F.J.W.; Morsink, F.H.M.; Verkade, H.J.; Petersen, C.; Hulscher, J.B.F. Liver fibrosis during the development of biliary atresia: Proof of principle in the murine model. J. Pediatr. Surg. 2015, 50, 1304–1309. [Google Scholar] [CrossRef]
- Tucker, R.M.; Feldman, A.G.; Fenner, E.K.; Mack, C.L. Regulatory T Cells Inhibit Th1 Cell-Mediated Bile Duct Injury in Murine Biliary Atresia. J. Hepatol. 2013, 59, 790–796. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Liu, Y.; Tang, S.; Yang, L.; Yang, J.; Cao, G.; Zhang, J.; Wang, X.; Mao, Y. Elevated Th17 Cells Accompanied by Decreased Regulatory T Cells and Cytokine Environment in Infants with Biliary Atresia. Pediatr. Surg. Int. 2013, 29, 1249–1260. [Google Scholar] [CrossRef] [PubMed]
- Veltkamp, C.; Anstaett, M.; Wahl, K.; Möller, S.; Gangl, S.; Bachmann, O.; Hardtke-Wolenski, M.; Länger, F.; Stremmel, W.; Manns, M.P.; et al. Apoptosis of Regulatory T Lymphocytes Is Increased in Chronic Inflammatory Bowel Disease and Reversed by Anti-TNFα Treatment. Gut 2011, 60, 1345–1353. [Google Scholar] [CrossRef] [PubMed]
- Canbay, A.; Feldstein, A.; Baskin-Bey, E.; Bronk, S.F.; Gores, G.J. The Caspase Inhibitor IDN-6556 Attenuates Hepatic Injury and Fibrosis in the Bile Duct Ligated Mouse. J. Pharmacol. Exp. Ther. 2004, 308, 1191–1196. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mouse Strain (N) | Healthy (%) | Biliary Atresia (%) |
|---|---|---|
| BALB/c (N = 91) | 28 (31%) | 63 (69%) |
| C57BL/6 (N = 101) | 79 (78%) | 22 (22%) |
| Total | 107 | 85 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Madadi-Sanjani, O.; Bohlen, G.; Wehrmann, F.; Andruszkow, J.; Khelif, K.; von Wasielewski, R.; Bantel, H.; Petersen, C. Increased Serum Levels of Activated Caspases in Murine and Human Biliary Atresia. J. Clin. Med. 2021, 10, 2718. https://doi.org/10.3390/jcm10122718
Madadi-Sanjani O, Bohlen G, Wehrmann F, Andruszkow J, Khelif K, von Wasielewski R, Bantel H, Petersen C. Increased Serum Levels of Activated Caspases in Murine and Human Biliary Atresia. Journal of Clinical Medicine. 2021; 10(12):2718. https://doi.org/10.3390/jcm10122718
Chicago/Turabian StyleMadadi-Sanjani, Omid, Gunnar Bohlen, Fabian Wehrmann, Julia Andruszkow, Karim Khelif, Reinhard von Wasielewski, Heike Bantel, and Claus Petersen. 2021. "Increased Serum Levels of Activated Caspases in Murine and Human Biliary Atresia" Journal of Clinical Medicine 10, no. 12: 2718. https://doi.org/10.3390/jcm10122718
APA StyleMadadi-Sanjani, O., Bohlen, G., Wehrmann, F., Andruszkow, J., Khelif, K., von Wasielewski, R., Bantel, H., & Petersen, C. (2021). Increased Serum Levels of Activated Caspases in Murine and Human Biliary Atresia. Journal of Clinical Medicine, 10(12), 2718. https://doi.org/10.3390/jcm10122718