
Fatty Acid Binding Protein 3 (FABP3) and Apolipoprotein E4 (ApoE4) as Lipid Metabolism-Related Biomarkers of Alzheimer’s Disease

 ,
,  ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Biochemical Measurements
2.2. Statistical Analysis
3. Results
3.1. Concentrations of Potential Lipid-Related Proteins as Biomarkers Candidates
3.2. Associations between CSF Levels of FABP3, ApoE4 and Neurochemical Dementia Biomarkers (Aβ42/40 Ratio, Tau, pTau181)
3.3. Diagnostic Usefulness of Candidate Biomarkers
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dhiman, K.; Blennow, K.; Zetterberg, H.; Martins, R.N.; Gupta, V.B. Cerebrospinal fluid biomarkers for understanding multiple aspects of Alzheimer’s disease pathogenesis. Cell. Mol. Life Sci. 2019, 76, 1833–1863. [Google Scholar] [CrossRef]
- El Kadmiri, N.; Said, N.; Slassi, I.; El Moutawakil, B.; Nadifi, S. Biomarkers for Alzheimer Disease: Classical and Novel Candidates’ Review. Neuroscience 2018, 370, 181–190. [Google Scholar] [CrossRef]
- Kao, Y.-C.; Ho, P.-C.; Tu, Y.-K.; Jou, I.-M.; Tsai, K.-J. Lipids and Alzheimer’s Disease. Int. J. Mol. Sci. 2020, 21, 1505. [Google Scholar] [CrossRef]
- Park, S.A.; Han, S.M.; Kim, C.E. New fluid biomarkers tracking non-amyloid-β and non-tau pathology in Alzheimer’s disease. Exp. Mol. Med. 2020, 52, 556–568. [Google Scholar] [CrossRef] [Green Version]
- Grimm, M.O.W.; Michaelson, D.M.; Hartmann, T. Omega-3 fatty acids, lipids, and apoE lipidation in Alzheimer’s disease: A rationale for multi-nutrient dementia prevention. J. Lipid Res. 2017, 58, 2083–2101. [Google Scholar] [CrossRef] [Green Version]
- Cheng, A.; Shinoda, Y.; Yamamoto, T.; Miyachi, H.; Fukunaga, K. Development of FABP3 ligands that inhibit arachidonic acid-induced α-synuclein oligomerization. Brain Res. 2019, 1707, 190–197. [Google Scholar] [CrossRef]
- Farrer, L.A.; Cupples, L.A.; Haines, J.L.; Hyman, B.; Kukull, W.A.; Mayeux, R.; Myers, R.H.; Pericak-Vance, M.A.; Risch, N.; van Duijn, C.M. Effects of age, sex, and ethnicity on the association between apolipoprotein E genotype and Alzheimer disease. A meta-analysis. APOE and Alzheimer Disease Meta Analysis Consortium. JAMA 1997, 278, 1349–1356. [Google Scholar] [CrossRef] [PubMed]
- Serrano-Pozo, A.; Das, S.; Hyman, B.T. APOE and Alzheimer’s disease: Advances in genetics, pathophysiology, and therapeutic approaches. Lancet Neurol. 2021, 20, 68–80. [Google Scholar] [CrossRef]
- Liu, C.-C.; Kanekiyo, T.; Xu, H.; Bu, G. Apolipoprotein E and Alzheimer disease: Risk, mechanisms and therapy. Nat. Rev. Neurol. 2013, 9, 106–118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tachibana, M.; Holm, M.-L.; Liu, C.-C.; Shinohara, M.; Aikawa, T.; Oue, H.; Yamazaki, Y.; Martens, Y.A.; Murray, M.E.; Sullivan, P.M.; et al. APOE4-mediated amyloid-β pathology depends on its neuronal receptor LRP1. J. Clin. Investig. 2019, 129, 1272–1277. [Google Scholar] [CrossRef]
- Minta, K.; Brinkmalm, G.; Janelidze, S.; Sjödin, S.; Portelius, E.; Stomrud, E.; Zetterberg, H.; Blennow, K.; Hansson, O.; Andreasson, U. Quantification of total apolipoprotein E and its isoforms in cerebrospinal fluid from patients with neurodegenerative diseases. Alzheimer’s Res. Ther. 2020, 12, 19. [Google Scholar] [CrossRef]
- Desikan, R.S.; Thompson, W.K.; Holland, D.; Hess, C.P.; Brewer, J.B.; Zetterberg, H.; Blennow, K.; Andreassen, O.A.; McEvoy, L.K.; Hyman, B.T.; et al. Heart fatty acid binding protein and Aβ-associated Alzheimer’s neurodegeneration. Mol. Neurodegener. 2013, 8, 39. [Google Scholar] [CrossRef] [Green Version]
- Mouchard, A.; Boutonnet, M.-C.C.; Mazzocco, C.; Biendon, N.; Macrez, N. ApoE-fragment/Aβ heteromers in the brain of patients with Alzheimer’s disease. Sci. Rep. 2019, 9, 3989. [Google Scholar] [CrossRef] [Green Version]
- Verghese, P.B.; Castellano, J.M.; Garai, K.; Wang, Y.; Jiang, H.; Shah, A.; Bu, G.; Frieden, C.; Holtzman, D.M. ApoE influences amyloid-β (Aβ) clearance despite minimal apoE/Aβ association in physiological conditions. Proc. Natl. Acad. Sci. USA 2013, 110, E1807–E1816. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.-W.A.; Zhou, B.; Nabet, A.M.; Wernig, M.; Südhof, T.C. Differential Signaling Mediated by ApoE2, ApoE3, and ApoE4 in Human Neurons Parallels Alzheimer’s Disease Risk. J. Neurosci. 2019, 39, 7408–7427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simon, R.; Girod, M.; Fonbonne, C.; Salvador, A.; Clément, Y.; Lantéri, P.; Amouyel, P.; Lambert, J.C.; Lemoine, J. Total ApoE and ApoE4 Isoform Assays in an Alzheimer’s Disease Case-control Study by Targeted Mass Spectrometry (n = 669): A Pilot Assay for Methionine-containing Proteotypic Peptides. Mol. Cell. Proteom. 2012, 11, 1389–1403. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, C.; Gerlach, N.; Peter, C.; Gherib, K.; Lange, K.; Friede, T.; Zerr, I. Cerebrospinal fluid Apolipoprotein e concentration and progression of Alzheimer’s disease. J. Alzheimer’s Dis. 2014, 43, 1229–1236. [Google Scholar] [CrossRef] [PubMed]
- Merched, A.; Blain, H.; Visvikis, S.; Herbeth, B.; Jeandel, C.; Siest, G. Cerebrospinal fluid apolipoprotein E level is increased in late-onset Alzheimer’s disease. J. Neurol. Sci. 1997, 145, 33–39. [Google Scholar] [CrossRef]
- Baker-Nigh, A.T.; Mawuenyega, K.G.; Bollinger, J.G.; Ovod, V.; Kasten, T.; Franklin, E.E.; Liao, F.; Jiang, H.; Holtzman, D.; Cairns, N.J.; et al. Human Central Nervous System (CNS) apoE isoforms are increased by age, differentially altered by amyloidosis, and relative amounts reversed in the CNS compared with plasma. J. Biol. Chem. 2016, 291, 27204–27218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hesse, C.; Larsson, H.; Fredman, P.; Minthon, L.; Andreasen, N.; Davidsson, P.; Blennow, K. Measurement of apolipoprotein E (apoE) in cerebrospinal fluid. Neurochem. Res. 2000, 25, 511–517. [Google Scholar] [CrossRef]
- Martínez-Morillo, E.; Hansson, O.; Atagi, Y.; Bu, G.; Minthon, L.; Diamandis, E.P.; Nielsen, H.M. Total apolipoprotein E levels and specific isoform composition in cerebrospinal fluid and plasma from Alzheimer’s disease patients and controls. Acta Neuropathol. 2014, 127, 633–643. [Google Scholar] [CrossRef]
- D’Anneo, A.; Bavisotto, C.C.; Gammazza, A.M.; Paladino, L.; Carlisi, D.; Cappello, F.; de Macario, E.C.; Macario, A.J.L.; Lauricella, M. Lipid chaperones and associated diseases: A group of chaperonopathies defining a new nosological entity with implications for medical research and practice. Cell Stress Chaperones 2020, 25, 805–820. [Google Scholar] [CrossRef] [PubMed]
- Höglund, K.; Kern, S.; Zettergren, A.; Börjesson-Hansson, A.; Zetterberg, H.; Skoog, I.; Blennow, K. Preclinical amyloid pathology biomarker positivity: Effects on tau pathology and neurodegeneration. Transl. Psychiatry 2017, 7, e995. [Google Scholar] [CrossRef]
- Shioda, N.; Yabuki, Y.; Kobayashi, Y.; Onozato, M.; Owada, Y.; Fukunaga, K. FABP3 protein promotes α-synuclein oligomerization associated with 1-Methyl-1,2,3,6-tetrahydropiridine-induced neurotoxicity. J. Biol. Chem. 2014, 289, 18957–18965. [Google Scholar] [CrossRef] [Green Version]
- Owada, Y. Fatty Acid Binding Protein: Localization and Functional Significance in the Brain. Tohoku J. Exp. Med. 2008, 214, 213–220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murphy, E.J.; Owada, Y.; Kitanaka, N.; Kondo, H.; Glatz, J.F.C. Brain arachidonic acid incorporation is decreased in heart fatty acid binding protein gene-ablated mice. Biochemistry 2005, 44, 6350–6360. [Google Scholar] [CrossRef]
- Shioda, N.; Yamamoto, Y.; Watanabe, M.; Binas, B.; Owada, Y.; Fukunaga, K. Heart-Type Fatty Acid Binding Protein Regulates Dopamine D2 Receptor Function in Mouse Brain. J. Neurosci. 2010, 30, 3146–3155. [Google Scholar] [CrossRef] [PubMed]
- Matsumata, M.; Inada, H.; Osumi, N. Fatty acid binding proteins and the nervous system: Their impact on mental conditions. Neurosci. Res. 2016, 102, 47–55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiasserini, D.; Parnetti, L.; Andreasson, U.; Zetterberg, H.; Giannandrea, D.; Calabresi, P.; Blennow, K. CSF levels of heart fatty acid binding protein are altered during early phases of Alzheimer’s disease. J. Alzheimer’s Dis. 2010, 22, 1281–1288. [Google Scholar] [CrossRef] [Green Version]
- Bjerke, M.; Zetterberg, H.; Edman, Å.; Blennow, K.; Wallin, A.; Andreasson, U. Cerebrospinal Fluid Matrix Metalloproteinases and Tissue Inhibitor of Metalloproteinases in Combination with Subcortical and Cortical Biomarkers in Vascular Dementia and Alzheimer’s Disease. J. Alzheimer’s Dis. 2011, 27, 665–676. [Google Scholar] [CrossRef] [PubMed]
- Chiasserini, D.; Biscetti, L.; Eusebi, P.; Salvadori, N.; Frattini, G.; Simoni, S.; De Roeck, N.; Tambasco, N.; Stoops, E.; Vanderstichele, H.; et al. Differential role of CSF fatty acid binding protein 3, α-synuclein, and Alzheimer’s disease core biomarkers in Lewy body disorders and Alzheimer’s dementia. Alzheimer’s Res. Ther. 2017, 9, 52. [Google Scholar] [CrossRef]
- Albert, M.S.; DeKosky, S.T.; Dickson, D.; Dubois, B.; Feldman, H.H.; Fox, N.C.; Gamst, A.; Holtzman, D.M.; Jagust, W.J.; Petersen, R.C.; et al. The diagnosis of mild cognitive impairment due to Alzheimer’s disease: Recommendations from the National Institute on Aging-Alzheimer’s Association workgroups on diagnostic guidelines for Alzheimer’s disease. Alzheimer’s Dement. 2011, 7, 270–279. [Google Scholar] [CrossRef] [Green Version]
- McKhann, G.M.; Knopman, D.S.; Chertkow, H.; Hyman, B.T.; Jack, C.R.; Kawas, C.H.; Klunk, W.E.; Koroshetz, W.J.; Manly, J.J.; Mayeux, R.; et al. The diagnosis of dementia due to Alzheimer’s disease: Recommendations from the National Institute on Aging-Alzheimer’s Association workgroups on diagnostic guidelines for Alzheimer’s disease. Alzheimer’s Dement. 2011, 7, 263–269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lewczuk, P.; Zimmermann, R.; Wiltfang, J.; Kornhuber, J. Neurochemical dementia diagnostics: A simple algorithm for interpretation of the CSF biomarkers. J. Neural Transm. 2009, 116, 1163–1167. [Google Scholar] [CrossRef] [PubMed]
- Wong, M.W.; Braidy, N.; Poljak, A.; Pickford, R.; Thambisetty, M.; Sachdev, P.S. Dysregulation of lipids in Alzheimer’s disease and their role as potential biomarkers. Alzheimer’s Dement. 2017, 13, 810–827. [Google Scholar] [CrossRef]
- Fonteh, A.N.; Cipolla, M.; Chiang, A.J.; Edminster, S.P.; Arakaki, X.; Harrington, M.G. Polyunsaturated Fatty Acid Composition of Cerebrospinal Fluid Fractions Shows Their Contribution to Cognitive Resilience of a Pre-symptomatic Alzheimer’s Disease Cohort. Front. Physiol. 2020, 11, 1–14. [Google Scholar] [CrossRef]
- Guo, L.H.; Alexopoulos, P.; Perneczky, R. Heart-type fatty acid binding protein and vascular endothelial growth factor: Cerebrospinal fluid biomarker candidates for Alzheimer’s disease. Eur. Arch. Psychiatry Clin. Neurosci. 2013, 263, 553–560. [Google Scholar] [CrossRef]
- Guillaume, E.; Zimmermann, C.; Burkhard, P.R.; Hochstrasser, D.F.; Sanchez, J.C. A potential cerebrospinal fluid and plasmatic marker for the diagnosis of Creutzfeldt-Jakob disease. Proteomics 2003, 3, 1495–1499. [Google Scholar] [CrossRef] [PubMed]
- Wada-Isoe, K.; Imamura, K.; Kitamaya, M.; Kowa, H.; Nakashima, K. Serum heart-fatty acid binding protein levels in patients with Lewy body disease. J. Neurol. Sci. 2008, 266, 20–24. [Google Scholar] [CrossRef]
- Teunissen, C.E.; Veerhuis, R.; De Vente, J.; Verhey, F.R.J.; Vreeling, F.; van Boxtel, M.P.J.; Glatz, J.F.C.; Pelsers, M.A.L. Brain-specific fatty acid-binding protein is elevated in serum of patients with dementia-related diseases. Eur. J. Neurol. 2011, 18, 865–871. [Google Scholar] [CrossRef] [PubMed]
- Petersen, M.E.; Zhang, F.; Schupf, N.; Krinsky-McHale, S.J.; Hall, J.; Mapstone, M.; Cheema, A.; Silverman, W.; Lott, I.; Rafii, M.S.; et al. Proteomic profiles for Alzheimer’s disease and mild cognitive impairment among adults with Down syndrome spanning serum and plasma: An Alzheimer’s Biomarker Consortium–Down Syndrome (ABC–DS) study. Alzheimer’s Dement. Diagn. Assess. Dis. Monit. 2020, 12, e12039. [Google Scholar] [CrossRef]
- Yamamoto, Y.; Kida, H.; Kagawa, Y.; Yasumoto, Y.; Miyazaki, H.; Islam, A.; Ogata, M.; Yanagawa, Y.; Mitsushima, D.; Fukunaga, K.; et al. FABP3 in the Anterior Cingulate Cortex Modulates the Methylation Status of the Glutamic Acid Decarboxylase 67 Promoter Region. J. Neurosci. 2018, 38, 10411–10423. [Google Scholar] [CrossRef] [Green Version]
- Sellner, P.A.; Chu, W.; Glatz, J.F.C.; Berman, N.E.J. Developmental role of fatty acid-binding proteins in mouse brain. Dev. Brain Res. 1995, 89, 33–46. [Google Scholar] [CrossRef]
- Kawahata, I.; Sekimori, T.; Wang, H.; Wang, Y.; Sasaoka, T.; Bousset, L.; Melki, R.; Mizobata, T.; Kawata, Y.; Fukunaga, K. Dopamine D2 Long Receptors Are Critical for Caveolae-Mediated α-Synuclein Uptake in Cultured Dopaminergic Neurons. Biomedicines 2021, 9, 49. [Google Scholar] [CrossRef]
- Wang, Y.; Wu, Z.; Bai, Y.-T.; Wu, G.-Y.; Chen, G. Gad67 haploinsufficiency reduces amyloid pathology and rescues olfactory memory deficits in a mouse model of Alzheimer’s disease. Mol. Neurodegener. 2017, 12, 73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruan, Z.; Pathak, D.; Venkatesan Kalavai, S.; Yoshii-Kitahara, A.; Muraoka, S.; Bhatt, N.; Takamatsu-Yukawa, K.; Hu, J.; Wang, Y.; Hersh, S.; et al. Alzheimer’s disease brain-derived extracellular vesicles spread tau pathology in interneurons. Brain 2021, 144, 288–309. [Google Scholar] [CrossRef]
- Panza, F.; Solfrizzi, V.; Colacicco, A.M.; Basile, A.M.; D’Introno, A.; Capurso, C.; Sabba, M.; Capurso, S.; Capurso, A. Apolipoprotein E (APOE) polymorphism influences serum APOE levels in Alzheimer’s disease patients and centenarians. Neuroreport 2003, 14, 605–608. [Google Scholar] [CrossRef]
- Terrisse, L.; Poirier, J.; Bertrand, P.; Merched, A.; Visvikis, S.; Siest, G.; Milne, R.; Rassart, É. Increased Levels of Apolipoprotein D in Cerebrospinal Fluid and Hippocampus of Alzheimer’s Patients. J. Neurochem. 2002, 71, 1643–1650. [Google Scholar] [CrossRef]
- Lefranc, D.; Vermersch, P.; Dallongeville, J.; Daem49s-Monpeurt, C.; Petit, H.; Delacourte, A. Relevance of the quantification of apolipoprotein E in the cerebrospinal fluid in Alzheimer’s disease. Neurosci. Lett. 1996, 212, 91–94. [Google Scholar] [CrossRef]
- Rezeli, M.; Zetterberg, H.; Blennow, K.; Brinkmalm, A.; Laurell, T.; Hansson, O.; Marko-Varga, G. Quantification of total apolipoprotein E and its specific isoforms in cerebrospinal fluid and blood in Alzheimer’s disease and other neurodegenerative diseases. EuPA Open Proteom. 2015, 8, 137–143. [Google Scholar] [CrossRef] [Green Version]
- Castellano, J.M.; Deane, R.; Gottesdiener, A.J.; Verghese, P.B.; Stewart, F.R.; West, T.; Paoletti, A.C.; Kasper, T.R.; DeMattos, R.B.; Zlokovic, B.V.; et al. Low-density lipoprotein receptor overexpression enhances the rate of brain-to-blood Aβ clearance in a mouse model of β-amyloidosis. Proc. Natl. Acad. Sci. USA 2012, 109, 15502–15507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rawat, V.; Wang, S.; Sima, J.; Bar, R.; Liraz, O.; Gundimeda, U.; Parekh, T.; Chan, J.; Johansson, J.O.; Tang, C.; et al. ApoE4 Alters ABCA1 Membrane Trafficking in Astrocytes. J. Neurosci. 2019, 39, 9611–9622. [Google Scholar] [CrossRef]
- Nedergaard, M.; Goldman, S.A. Glymphatic failure as a final common pathway to dementia. Science 2020, 370, 50–56. [Google Scholar] [CrossRef] [PubMed]
- Benveniste, H.; Liu, X.; Koundal, S.; Sanggaard, S.; Lee, H.; Wardlaw, J. The Glymphatic System and Waste Clearance with Brain Aging: A Review. Gerontology 2018, 06519, 1–14. [Google Scholar] [CrossRef]
- Kim, J.; Basak, J.M.; Holtzman, D.M. The Role of Apolipoprotein E in Alzheimer’s Disease. Neuron 2009, 63, 287–303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qiao, F.; Gao, X.-P.; Yuan, L.; Cai, H.-Y.; Qi, J.-S. Apolipoprotein E4 Impairs in vivo Hippocampal Long-Term Synaptic Plasticity by Reducing the Phosphorylation of CaMKIIα and CREB. J. Alzheimer’s Dis. 2014, 41, 1165–1176. [Google Scholar] [CrossRef] [PubMed]
- Safieh, M.; Korczyn, A.D.; Michaelson, D.M. ApoE4: An emerging therapeutic target for Alzheimer’s disease. BMC Med. 2019, 17, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez, G.A.; Burns, M.P.; Weeber, E.J.; Rebeck, G.W. Young APOE4 targeted replacement mice exhibit poor spatial learning and memory, with reduced dendritic spine density in the medial entorhinal cortex. Learn. Mem. 2013, 20, 256–266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, Y.-T.; Seo, J.; Gao, F.; Feldman, H.M.; Wen, H.-L.; Penney, J.; Cam, H.P.; Gjoneska, E.; Raja, W.K.; Cheng, J.; et al. APOE4 Causes Widespread Molecular and Cellular Alterations Associated with Alzheimer’s Disease Phenotypes in Human iPSC-Derived Brain Cell Types. Neuron 2018, 98, 1141–1154.e7. [Google Scholar] [CrossRef] [Green Version]
- Sienski, G.; Narayan, P.; Bonner, J.M.; Kory, N.; Boland, S.; Arczewska, A.A.; Ralvenius, W.T.; Akay, L.; Lockshin, E.; He, L.; et al. APOE4 disrupts intracellular lipid homeostasis in human iPSC-derived glia. Sci. Transl. Med. 2021, 13, eaaz4564. [Google Scholar] [CrossRef]
- Farmer, B.; Kluemper, J.; Johnson, L. Apolipoprotein E4 Alters Astrocyte Fatty Acid Metabolism and Lipid Droplet Formation. Cells 2019, 8, 182. [Google Scholar] [CrossRef] [Green Version]
- Qi, G.; Mi, Y.; Shi, X.; Gu, H.; Brinton, R.D.; Yin, F. ApoE4 Impairs Neuron-Astrocyte Coupling of Fatty Acid Metabolism. Cell Rep. 2021, 34, 108572. [Google Scholar] [CrossRef] [PubMed]
- Olsson, B.; Lautner, R.; Andreasson, U.; Öhrfelt, A.; Portelius, E.; Bjerke, M.; Hölttä, M.; Rosén, C.; Olsson, C.; Strobel, G.; et al. CSF and blood biomarkers for the diagnosis of Alzheimer’s disease: A systematic review and meta-analysis. Lancet Neurol. 2016, 15, 673–684. [Google Scholar] [CrossRef]
- Ko, C.-W.; Qu, J.; Black, D.D.; Tso, P. Regulation of intestinal lipid metabolism: Current concepts and relevance to disease. Nat. Rev. Gastroenterol. Hepatol. 2020, 17, 169–183. [Google Scholar] [CrossRef]
- Liu, J.J.; Green, P.; John Mann, J.; Rapoport, S.I.; Sublette, M.E. Pathways of polyunsaturated fatty acid utilization: Implications for brain function in neuropsychiatric health and disease. Brain Res. 2015, 1597, 220–246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balakrishnan, J.; Kannan, S.; Govindasamy, A. Structured form of DHA prevents neurodegenerative disorders: A better insight into the pathophysiology and the mechanism of DHA transport to the brain. Nutr. Res. 2021, 85, 119–134. [Google Scholar] [CrossRef] [PubMed]
- Montagne, A.; Nation, D.A.; Sagare, A.P.; Barisano, G.; Sweeney, M.D.; Chakhoyan, A.; Pachicano, M.; Joe, E.; Nelson, A.R.; D’Orazio, L.M.; et al. APOE4 leads to blood–brain barrier dysfunction predicting cognitive decline. Nature 2020, 581, 71–76. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Tested Variables | Median (Interquartile Range) | p (Kruskal-Wallis Test) | p (Dwass-Steele-Critchlow-Flinger Test) | ||||
|---|---|---|---|---|---|---|---|
| AD | MCI | Controls | AD vs. CTRL | AD vs. MCI | MCI vs. CTRL | ||
| Aβ42/40 ratio CSF | 0.03 (0.02–0.04) | 0.05 (0.03–0.08) | 0.07 (0.06–0.08) | <0.001 | <0.001 | <0.001 | <0.001 |
| Tau (pg/mL) | 669 (561–943) | 389 (327–495) | 221 (190–256) | <0.001 | <0.001 | <0.001 | <0.001 |
| pTau181 (pg/mL) | 83 (69–111) | 57 (46–68) | 38 (34–41) | <0.001 | 0.001 | <0.001 | 0.002 |
| ApoE4 (ng/mL) CSF | 348,552 (8491–439,189) | 4491 (3911–157,341) | 9021 (6556–10,126) | <0.001 | 0.009 | 0.002 | 0.080 |
| FABP3 (pg/mL) CSF | 3704 (2937–4872) | 2380 (1669–2651) | 1752 (1514–2061) | <0.001 | <0.001 | <0.001 | 0.362 |
| Median (Interquartile Range) | |||
|---|---|---|---|
| AD n = 34 | MCI n = 18 | CTRL n = 18 | |
| Age (years) | 76 (68–81) | 75 (70–78) | 68 (64–75) |
| Gender (Female/Male) | 26/8 | 10/8 | 12/6 |
| MMSE score (range 0–30 p.) | 22 (19–24) | 27 (26–29) | 29 (27–30) |
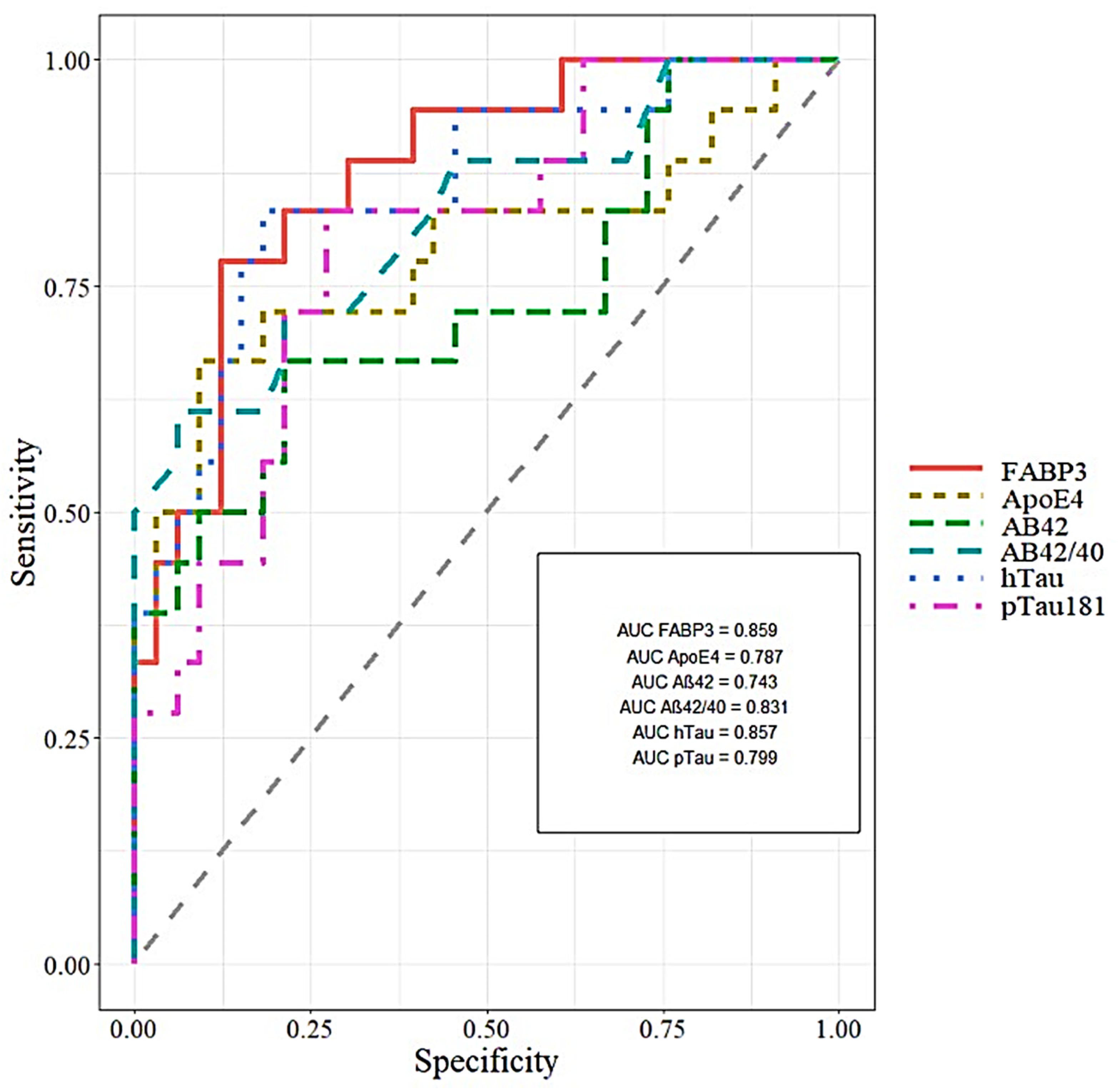
| Tested Parameters | ROC Criteria in AD Compared to CTRL | ROC Criteria in MCI Compared to AD | ||||||
|---|---|---|---|---|---|---|---|---|
| AUC | SE | 95% C.I. (AUC) | p (AUC = 0.5) | AUC | SE | 95% C.I. (AUC) | p (AUC = 0.5) | |
| FABP3 | 0.881 | 0.046 | 0.7646–0.9968 | <0.001 | 0.859 | 0.050 | 0.7569–0.962 | <0.001 |
| ApoE4 | 0.751 | 0.067 | 0.6195–0.8838 | 0.001 | 0.787 | 0.062 | 0.6349–0.9403 | <0.001 |
| Aβ42 | 0.930 | 0.034 | 0.8613–0.9998 | <0.001 | 0.743 | 0.068 | 0.5909–0.896 | <0.001 |
| Aβ42/40 | 1 | 0 | 1 | <0.001 | 0.831 | 0.055 | 0.7083–0.9551 | <0.001 |
| pTau181 | 0.969 | 0.022 | 0.9242–1 | <0.001 | 0.799 | 0.060 | 0.6725–0.9255 | <0.001 |
| Tau | 0.985 | 0.015 | 0.9614–1 | <0.001 | 0.857 | 0.050 | 0.746–0.9696 | <0.001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dulewicz, M.; Kulczyńska-Przybik, A.; Słowik, A.; Borawska, R.; Mroczko, B. Fatty Acid Binding Protein 3 (FABP3) and Apolipoprotein E4 (ApoE4) as Lipid Metabolism-Related Biomarkers of Alzheimer’s Disease. J. Clin. Med. 2021, 10, 3009. https://doi.org/10.3390/jcm10143009
Dulewicz M, Kulczyńska-Przybik A, Słowik A, Borawska R, Mroczko B. Fatty Acid Binding Protein 3 (FABP3) and Apolipoprotein E4 (ApoE4) as Lipid Metabolism-Related Biomarkers of Alzheimer’s Disease. Journal of Clinical Medicine. 2021; 10(14):3009. https://doi.org/10.3390/jcm10143009
Chicago/Turabian StyleDulewicz, Maciej, Agnieszka Kulczyńska-Przybik, Agnieszka Słowik, Renata Borawska, and Barbara Mroczko. 2021. "Fatty Acid Binding Protein 3 (FABP3) and Apolipoprotein E4 (ApoE4) as Lipid Metabolism-Related Biomarkers of Alzheimer’s Disease" Journal of Clinical Medicine 10, no. 14: 3009. https://doi.org/10.3390/jcm10143009
APA StyleDulewicz, M., Kulczyńska-Przybik, A., Słowik, A., Borawska, R., & Mroczko, B. (2021). Fatty Acid Binding Protein 3 (FABP3) and Apolipoprotein E4 (ApoE4) as Lipid Metabolism-Related Biomarkers of Alzheimer’s Disease. Journal of Clinical Medicine, 10(14), 3009. https://doi.org/10.3390/jcm10143009