
Prognostic Value of Interleukin-32 Expression and Its Correlation with the Infiltration of Natural Killer Cells in Cutaneous Melanoma


Abstract
:1. Introduction
2. Materials and Methods
2.1. IL32 mRNA Expression in Various Tumors
2.2. Analysis of the Correlation between IL32 Expression and Prognostic Value
2.3. Analysis of IL32 Gene Mutations in Skin Cutaneous Melanoma (SKCM)
2.4. Correlation between IL32 Expression and Infiltration of Various Immune Cells
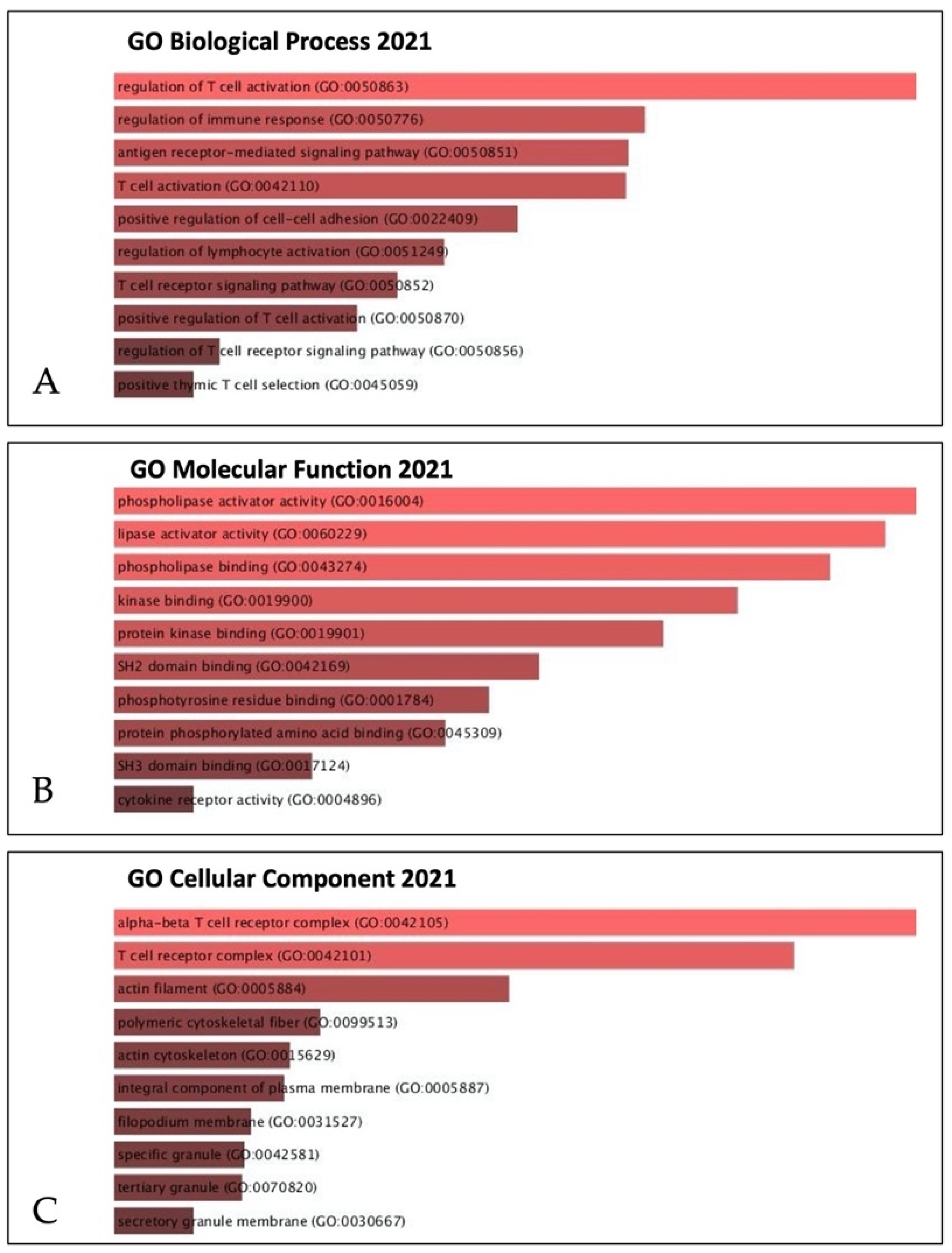
2.5. Correlation between Coexpressed Genes and IL32
2.6. Statistical Analysis
3. Results
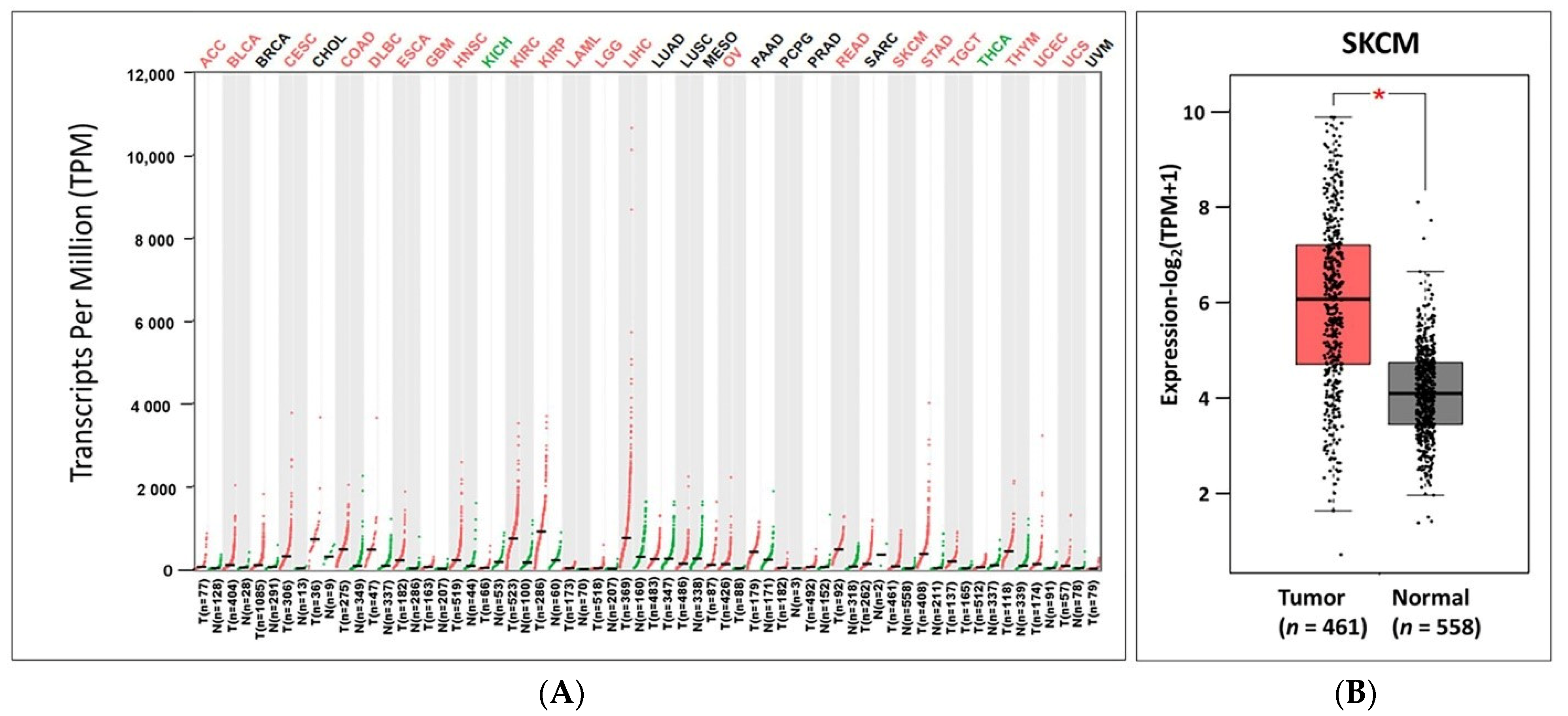
3.1. The Expression Analysis of IL32 in Various Types of Cancers
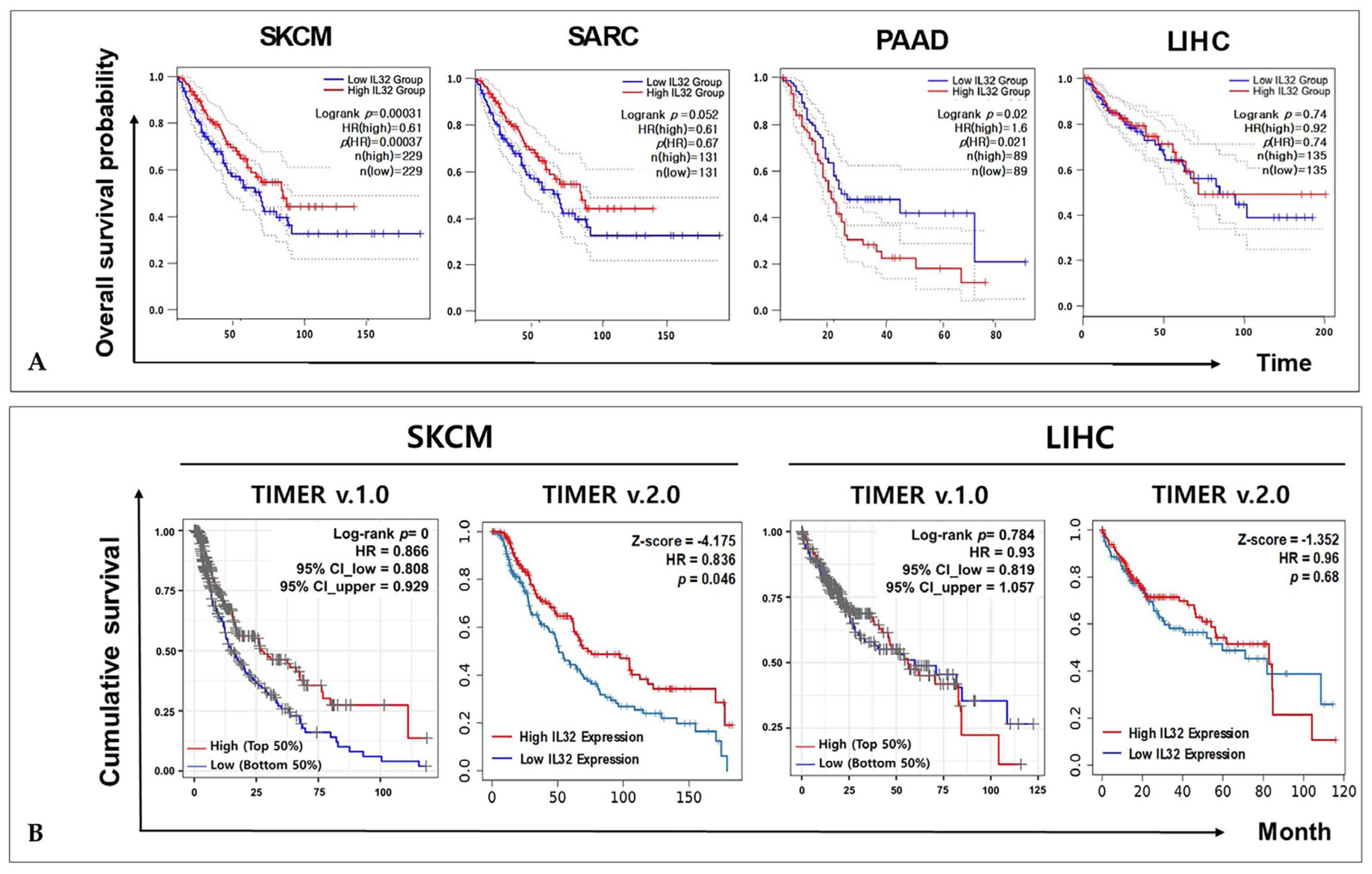
3.2. Correlation between IL32 Expression and Patient Survival in Various Types of Cancers
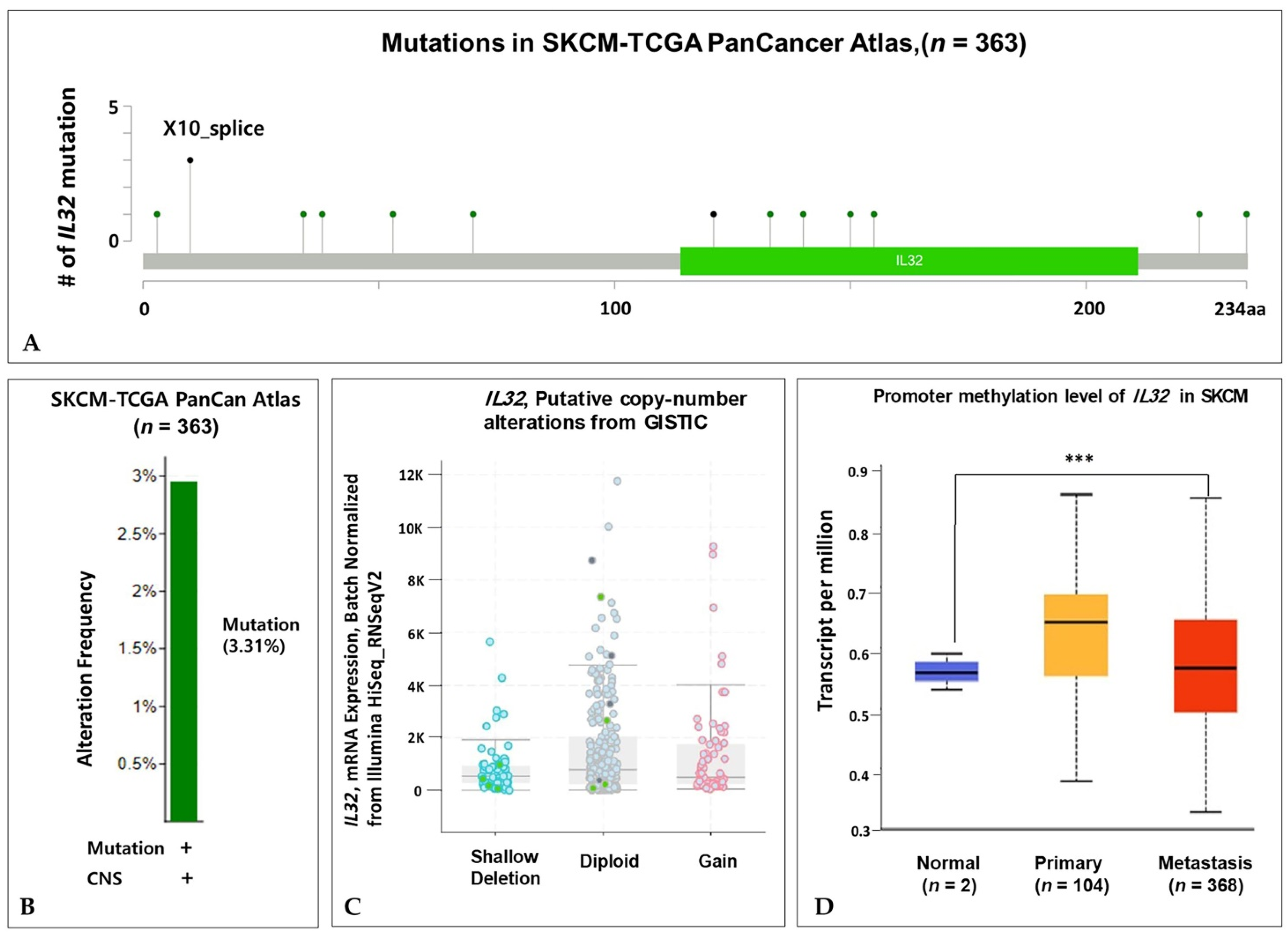
3.3. Genome Change of IL32 Expression in Melanoma
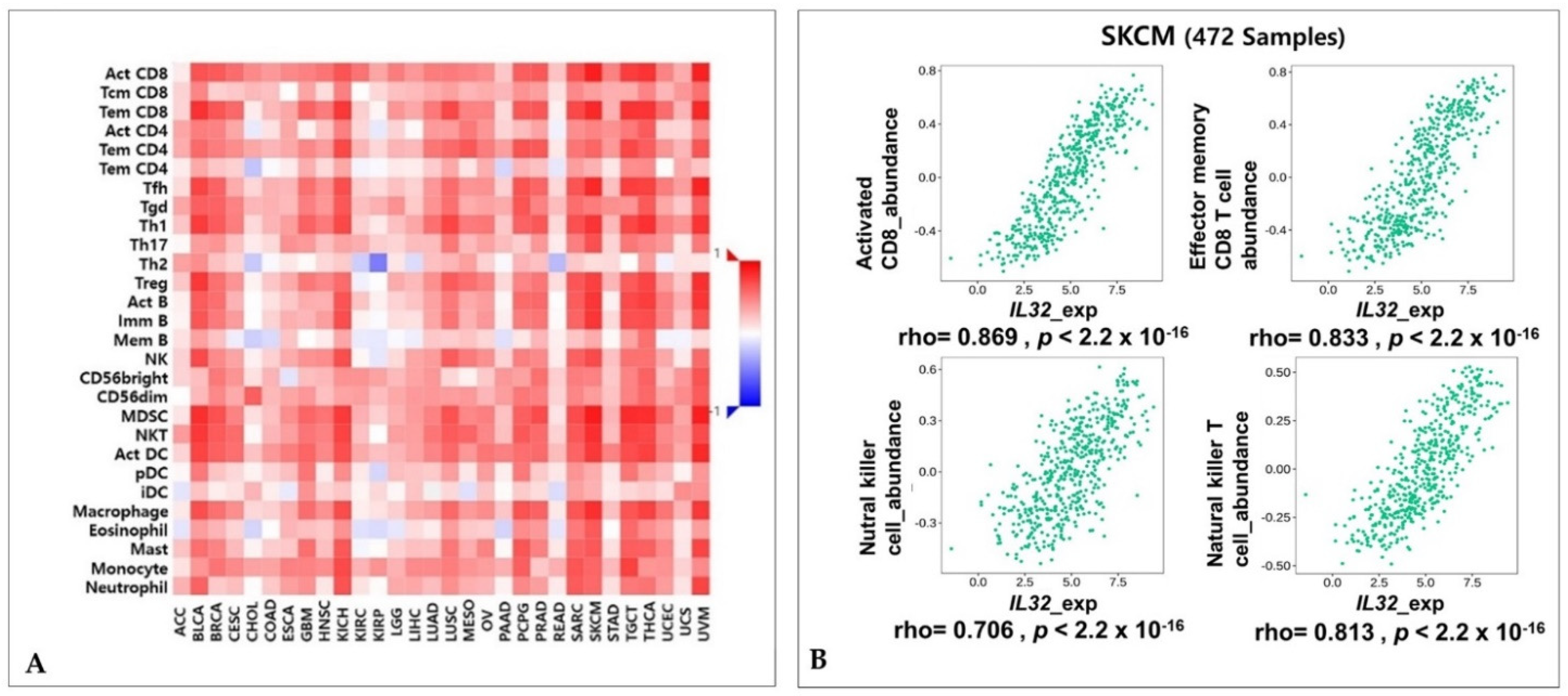
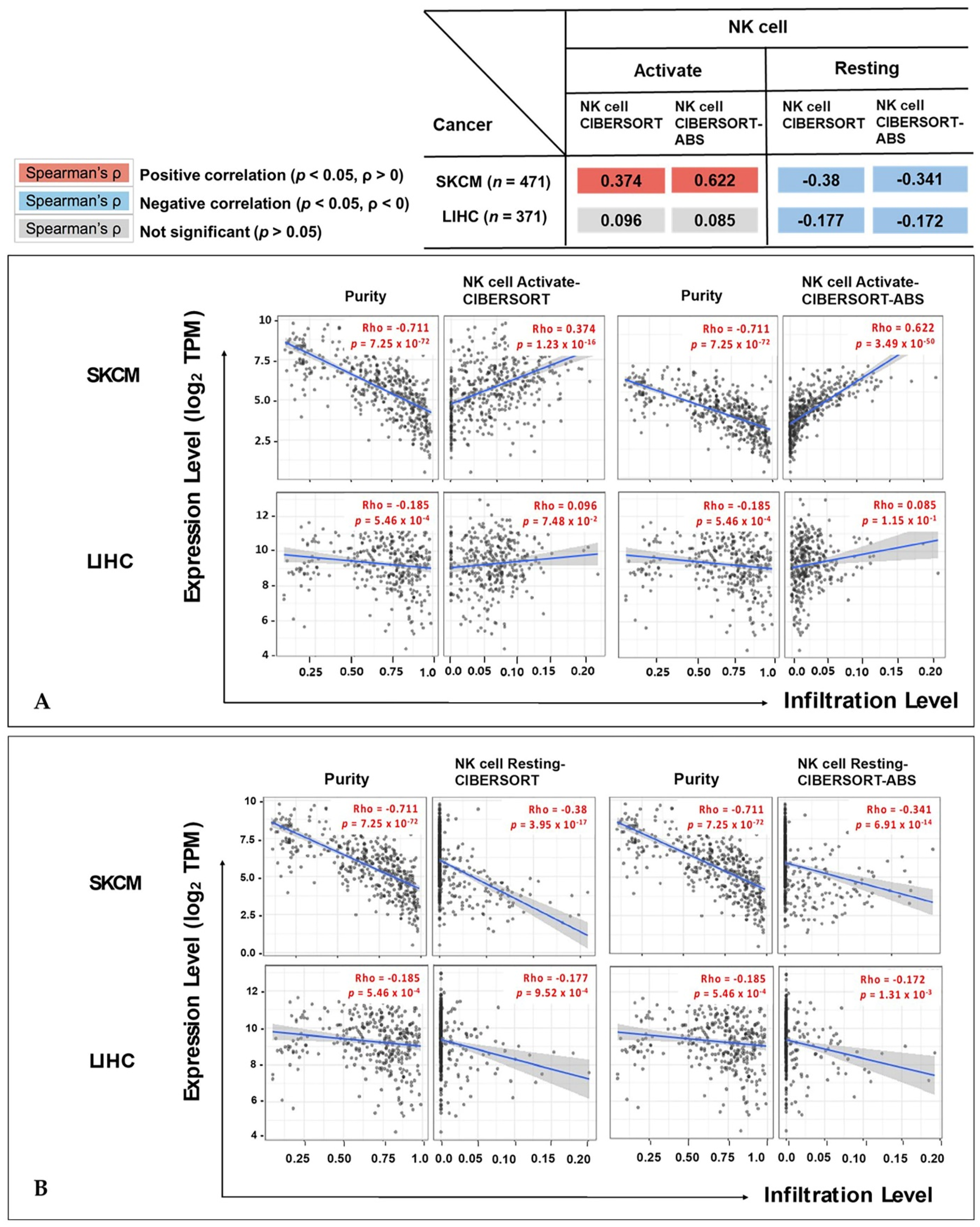
3.4. Correlation of IL32 Expression with Immune Infiltrates
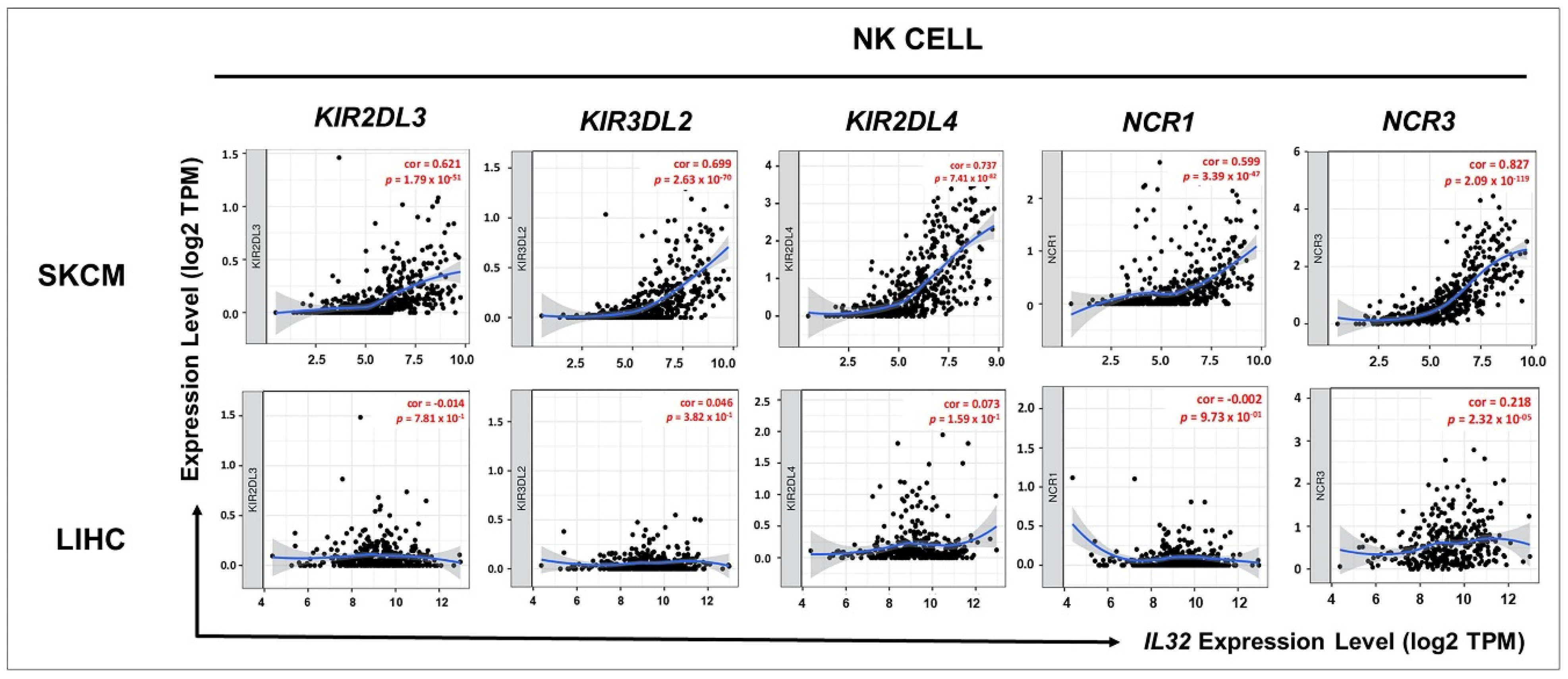
3.5. Correlation between IL32 Expression and Various Subsets of Immune Cells in Melanoma
3.6. Correlation between IL32 Expression and Activation of NK Cells
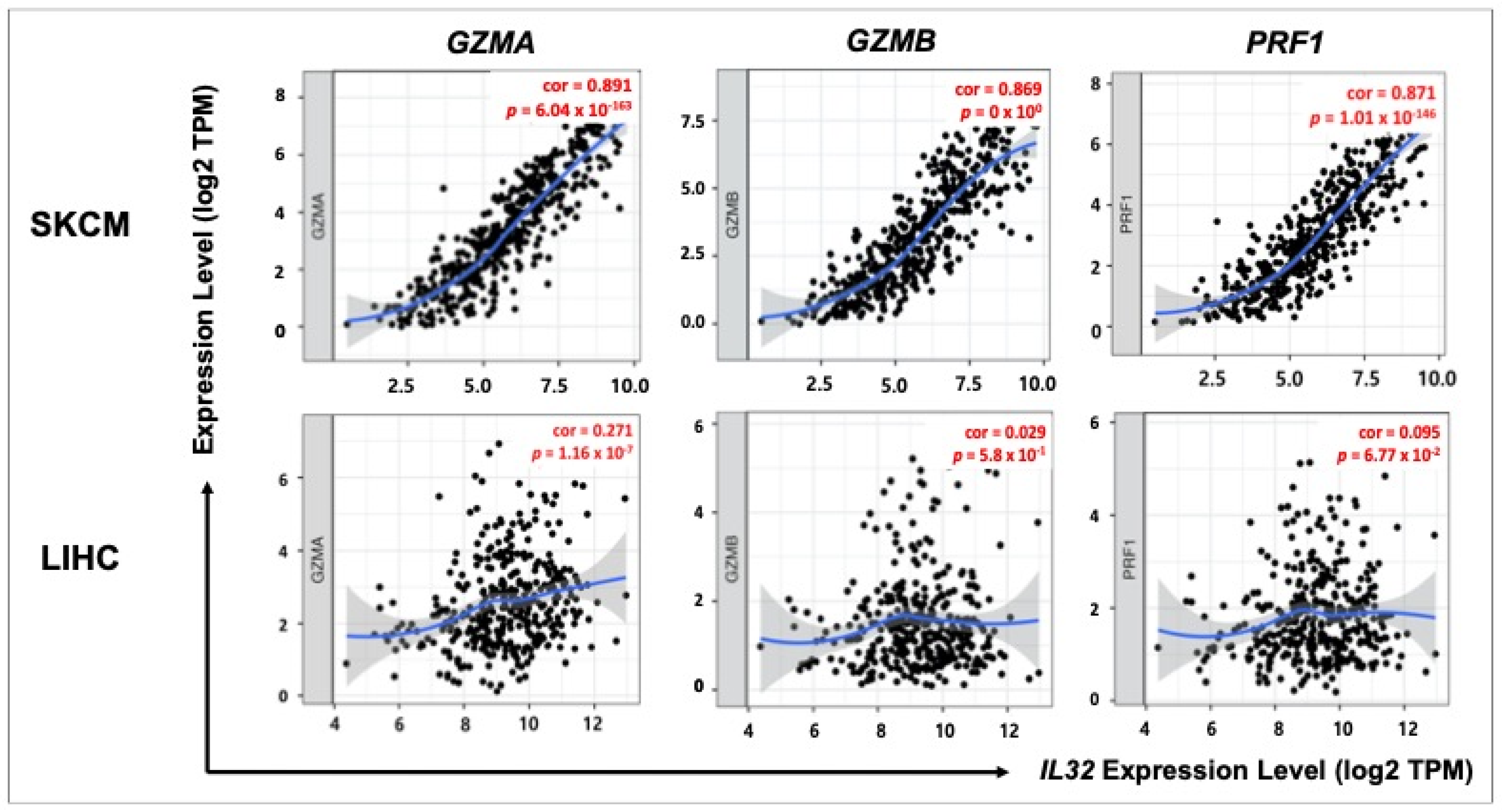
3.7. Correlation between IL32 Expression and Cytolytic Cell of NK Cells
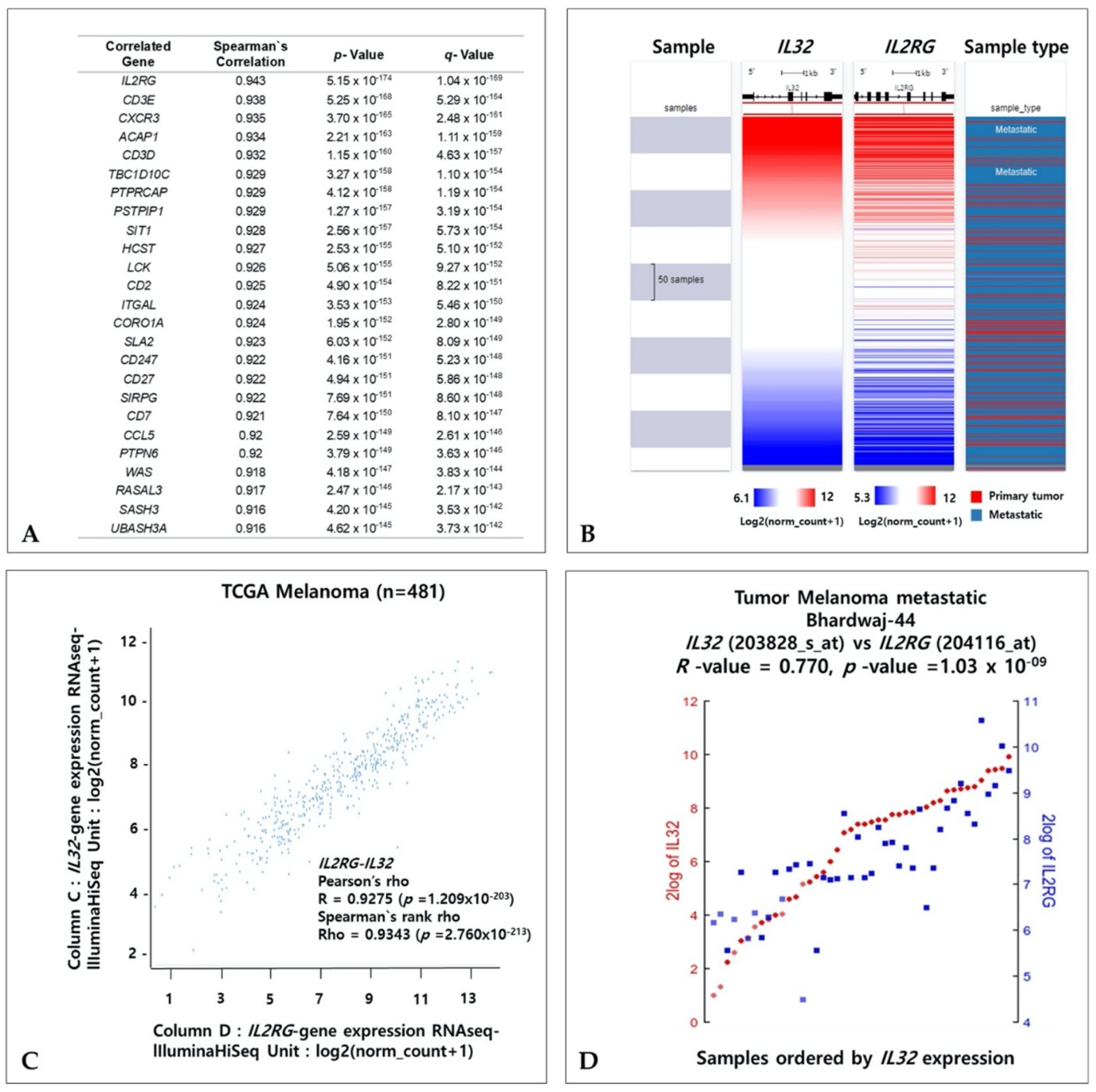
3.8. Coexpressed Genes and IL32 in Melanoma
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Schadendorf, D.; van Akkooi, A.C.J.; Berking, C.; Griewank, K.G.; Gutzmer, R.; Hauschild, A.; Stang, A.; Roesch, A.; Ugurel, S. Melanoma. Lancet 2018, 392, 971–984. [Google Scholar] [CrossRef]
- Li, Y.; Li, M.; Shats, I.; Krahn, J.M.; Flake, G.P.; Umbach, D.M.; Li, X.; Li, L. Glypican 6 is a putative biomarker for metastatic progression of cutaneous melanoma. PLoS ONE 2019, 14, e0218067. [Google Scholar] [CrossRef]
- Olszanski, A.J. Current and Future Roles of Targeted Therapy and Immunotherapy in Advanced Melanoma. J. Manag. Care Pharm. 2014, 20, 346–356. [Google Scholar] [CrossRef]
- Naik, P.P. Cutaneous Malignant Melanoma: A Review of Early Diagnosis and Management. World J. Oncol. 2021, 12, 7–19. [Google Scholar] [CrossRef]
- Fecher, L.A.; Cummings, S.D.; Keefe, M.J.; Alani, R.M. Toward a Molecular Classification of Melanoma. J. Clin. Oncol. 2007, 25, 1606–1620. [Google Scholar] [CrossRef]
- Chin, L.; Garraway, L.A.; Fisher, D.E. Malignant melanoma: Genetics and therapeutics in the genomic era. Genes Dev. 2006, 20, 2149–2182. [Google Scholar] [CrossRef]
- Soengas, M.S.; Lowe, S.W. Apoptosis and melanoma chemoresistance. Oncogene 2003, 22, 3138–3151. [Google Scholar] [CrossRef]
- Topalian, S.L.; Drake, C.G.; Pardoll, D.M. Immune Checkpoint Blockade: A Common Denominator Approach to Cancer Therapy. Cancer Cell 2015, 27, 450–461. [Google Scholar] [CrossRef] [PubMed]
- Ribas, A.; Wolchok, J.D. Cancer immunotherapy using checkpoint blockade. Science 2018, 359, 1350–1355. [Google Scholar] [CrossRef] [PubMed]
- Maibach, F.; Sadozai, H.; Jafari, S.M.S.; Hunger, R.E.; Schenk, M. Tumor-Infiltrating Lymphocytes and Their Prognostic Value in Cutaneous Melanoma. Front. Immunol. 2020, 11, 2105. [Google Scholar] [CrossRef] [PubMed]
- Fridman, W.H.; Zitvogel, L.; Sautes-Fridman, C.; Kroemer, G. The immune contexture in cancer prognosis and treatment. Nat. Rev. Clin. Oncol. 2017, 14, 717–734. [Google Scholar] [CrossRef]
- Park, Y.-J.; Kuen, D.-S.; Chung, Y. Future prospects of immune checkpoint blockade in cancer: From response prediction to overcoming resistance. Exp. Mol. Med. 2018, 50, 1–13. [Google Scholar] [CrossRef]
- Barnes, T.A.; Amir, E. HYPE or HOPE: The prognostic value of infiltrating immune cells in cancer. Br. J. Cancer 2018, 118, e5. [Google Scholar] [CrossRef] [PubMed]
- Curtin, J.A.; Fridlyand, J.; Kageshita, T.; Patel, H.N.; Busam, K.J.; Kutzner, H.; Cho, K.-H.; Aiba, S.; Bröcker, E.-B.; LeBoit, P.E.; et al. Distinct Sets of Genetic Alterations in Melanoma. N. Engl. J. Med. 2005, 353, 2135–2147. [Google Scholar] [CrossRef] [PubMed]
- Passarelli, A.; Mannavola, F.; Stucci, L.S.; Tucci, M.; Silvestris, F. Immune system and melanoma biology: A balance between immunosurveillance and immune escape. Oncotarget 2017, 8, 106132–106142. [Google Scholar] [CrossRef] [PubMed]
- Falletta, P.; del Campo, L.S.; Chauhan, J.; Effern, M.; Kenyon, A.; Kershaw, C.; Siddaway, R.; Lisle, R.J.; Freter, R.; Daniels, M.; et al. Translation reprogramming is an evolutionarily conserved driver of phenotypic plasticity and therapeutic resistance in melanoma. Genes Dev. 2017, 31, 18–33. [Google Scholar] [CrossRef] [PubMed]
- Kim, I.S.; Heilmann, S.; Kansler, E.R.; Zhang, Y.; Zimmer, M.; Ratnakumar, K.; Bowman, R.L.; Simon-Vermot, T.; Fennell, M.; Garippa, R.; et al. Microenvironment-derived factors driving metastatic plasticity in melanoma. Nat. Commun. 2017, 8, 14343. [Google Scholar] [CrossRef]
- Falcone, I.; Conciatori, F.; Bazzichetto, C.; Ferretti, G.; Cognetti, F.; Ciuffreda, L.; Milella, M. Tumor Microenvironment: Implications in Melanoma Resistance to Targeted Therapy and Immunotherapy. Cancers 2020, 12, 2870. [Google Scholar] [CrossRef]
- Letca, A.F.; Ungureanu, L.; Șenilă, S.C.; Grigore, L.E.; Pop, S.; Fechete, O.; Vesa, Ş.C.; Cosgarea, R. Regression and Sentinel Lymph Node Status in Melanoma Progression. Med. Sci. Monit. 2018, 24, 1359–1365. [Google Scholar] [CrossRef]
- Paijens, S.T.; Vledder, A.; de Bruyn, M.; Nijman, H.W. Tumor-infiltrating lymphocytes in the immunotherapy era. Cell. Mol. Immunol. 2021, 18, 842–859. [Google Scholar] [CrossRef]
- Gruosso, T.; Gigoux, M.; Manem, V.S.K.; Bertos, N.; Zuo, D.; Perlitch, I.; Saleh, S.M.I.; Zhao, H.; Souleimanova, M.; Johnson, R.M.; et al. Spatially distinct tumor immune microenvironments stratify triple-negative breast cancers. J. Clin. Investig. 2019, 129, 1785–1800. [Google Scholar] [CrossRef]
- Gata, V.A.; Lisencu, C.I.; Vlad, C.I.; Piciu, D.; Irimie, A.; Achimas-Cadariu, P. Tumor infiltrating lymphocytes as a prognostic factor in malignant melanoma. Review of the literature. J. BUON 2017, 22, 592–598. [Google Scholar]
- Gata, V.A.; Kubelac, P.M.; Buiga, R.; Vlad, I.C.; Valean, D.; Muntean, M.V.; Morariu, D.S.; Bonci, E.A.; Irimie, A.; Dina, C.; et al. The value of tumor infiltrating lymphocytes as prognostic factor for lymph node status and survival amongst patients with cutaneous malignant melanoma. J. BUON 2021, 25, 2700–2707. [Google Scholar]
- Maimela, N.R.; Liu, S.; Zhang, Y. Fates of CD8+ T cells in Tumor Microenvironment. Comput. Struct. Biotechnol. J. 2019, 17, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.-Y.; Fu, T.; Jiang, Y.-Z.; Shao, Z.-M. Natural killer cells in cancer biology and therapy. Mol. Cancer 2020, 19, 120. [Google Scholar] [CrossRef] [PubMed]
- Gajewski, T.F.; Schreiber, H.; Fu, Y.-X. Innate and adaptive immune cells in the tumor microenvironment. Nat. Immunol. 2013, 14, 1014–1022. [Google Scholar] [CrossRef]
- Saddawi-Konefka, R.; Seelige, R.; Gross, E.T.; Levy, E.; Searles, S.C.; Washington, A.; Santosa, E.K.; Liu, B.; O’Sullivan, T.; Harismendy, O.; et al. Nrf2 Induces IL-17D to Mediate Tumor and Virus Surveillance. Cell Rep. 2016, 16, 2348–2358. [Google Scholar] [CrossRef] [PubMed]
- Seelige, R.; Searles, S.; Bui, J.D. Mechanisms regulating immune surveillance of cellular stress in cancer. Cell. Mol. Life Sci. 2017, 75, 225–240. [Google Scholar] [CrossRef] [PubMed]
- Kmiecik, J.; Poli, A.; Brons, N.H.; Waha, A.; Eide, G.E.; Enger, P.Ø.; Zimmer, J.; Chekenya, M. Elevated CD3+ and CD8+ tumor-infiltrating immune cells correlate with prolonged survival in glioblastoma patients despite integrated immunosuppressive mechanisms in the tumor microenvironment and at the systemic level. J. Neuroimmunol. 2013, 264, 71–83. [Google Scholar] [CrossRef] [PubMed]
- Konjević, G.; Martinović, K.M.; Vuletić, A.; Jović, V.; Jurisic, V.; Babović, N.; Spužić, I. Low expression of CD161 and NKG2D activating NK receptor is associated with impaired NK cell cytotoxicity in metastatic melanoma patients. Clin. Exp. Metastasis 2007, 24, 1–11. [Google Scholar] [CrossRef]
- Konjević, G.; Martinović, K.M.; Vuletić, A.; Babović, N. In-vitro IL-2 or IFN-α-induced NKG2D and CD161 NK cell receptor expression indicates novel aspects of NK cell activation in metastatic melanoma patients. Melanoma Res. 2010, 20, 459–467. [Google Scholar] [CrossRef]
- Dahl, C.A.; Schall, R.P.; He, H.L.; Cairns, J.S. Identification of a novel gene expressed in activated natural killer cells and T cells. J. Immunol. 1992, 148, 597–603. [Google Scholar]
- Yan, H.; He, D.; Huang, X.; Zhang, E.; Chen, Q.; Xu, R.; Liu, X.; Zi, F.; Cai, Z. Role of interleukin-32 in cancer biology (Review). Oncol. Lett. 2018, 16, 41–47. [Google Scholar] [CrossRef]
- Paz, H.; Tsoi, J.; Kalbasi, A.; Grasso, C.S.; McBride, W.H.; Schaue, D.; Butterfield, L.H.; Maurer, D.M.; Ribas, A.; Graeber, T.G.; et al. Interleukin 32 expression in human melanoma. J. Transl. Med. 2019, 17, 113. [Google Scholar] [CrossRef]
- Dinarello, C.A.; Kim, S.-H. IL-32, a novel cytokine with a possible role in disease. Ann. Rheum. Dis. 2006, 65, iii61–iii64. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.-D.; Bae, S.-Y.; Hong, J.-W.; Azam, T.; Dinarello, C.A.; Her, E.; Choi, W.-S.; Kim, B.-K.; Lee, C.-K.; Yoon, D.-Y.; et al. Identification of the most active interleukin-32 isoform. Immunology 2009, 126, 535–542. [Google Scholar] [CrossRef] [PubMed]
- Yun, H.-M.; Park, K.-R.; Kim, E.-C.; Han, S.B.; Yoon, D.Y.; Hong, J.T. IL-32α suppresses colorectal cancer development via TNFR1-mediated death signaling. Oncotarget 2015, 6, 9061–9072. [Google Scholar] [CrossRef] [PubMed]
- Sloot, Y.J.E.; Smit, J.W.; Joosten, L.A.B.; Netea-Maier, R.T. Insights into the role of IL-32 in cancer. Semin. Immunol. 2018, 38, 24–32. [Google Scholar] [CrossRef]
- Gruber, T.; Kremenovic, M.; Sadozai, H.; Rombini, N.; Baeriswyl, L.; Maibach, F.; Modlin, R.L.; Gilliet, M.; von Werdt, D.; Hunger, R.E.; et al. IL-32γ potentiates tumor immunity in melanoma. JCI Insight 2020, 5, 5. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Q.; Li, S.; Zhou, Y.; Ou, W.; Cai, X.; Zhang, L.; Huang, W.; Huang, L.; Wang, Q. Interleukin-32 contributes to invasion and metastasis of primary lung adenocarcinoma via NF-kappaB induced matrix metalloproteinases 2 and 9 expression. Cytokine 2014, 65, 24–32. [Google Scholar] [CrossRef] [PubMed]
- Tsai, C.-Y.; Wang, C.-S.; Tsai, M.-M.; Chi, H.-C.; Cheng, W.-L.; Tseng, Y.-H.; Chen, C.-Y.; Lin, C.D.; Wu, J.-I.; Wang, L.-H.; et al. Interleukin-32 Increases Human Gastric Cancer Cell Invasion Associated with Tumor Progression and Metastasis. Clin. Cancer Res. 2014, 20, 2276–2288. [Google Scholar] [CrossRef]
- Wang, S.; Chen, F.; Tang, L. IL-32 promotes breast cancer cell growth and invasiveness. Oncol. Lett. 2015, 9, 305–307. [Google Scholar] [CrossRef]
- Park, J.S.; Choi, S.Y.; Lee, J.-H.; Lee, M.; Nam, E.S.; Jeong, A.L.; Lee, S.; Han, S.; Lee, M.-S.; Lim, J.-S.; et al. Interleukin-32β stimulates migration of MDA-MB-231 and MCF-7cells via the VEGF-STAT3 signaling pathway. Cell. Oncol. 2013, 36, 493–503. [Google Scholar] [CrossRef] [PubMed]
- Tang, Z.; Kang, B.; Li, C.; Chen, T.; Zhang, Z. GEPIA2: An enhanced web server for large-scale expression profiling and interactive analysis. Nucleic Acids Res. 2019, 47, W556–W560. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Fan, J.; Wang, B.; Traugh, N.; Chen, Q.; Liu, J.S.; Li, B.; Liu, X.S. TIMER: A Web Server for Comprehensive Analysis of Tumor-Infiltrating Immune Cells. Cancer Res. 2017, 77, e108–e110. [Google Scholar] [CrossRef] [PubMed]
- Chandrashekar, D.S.; Bashel, B.; Balasubramanya, S.A.H.; Creighton, C.J.; Rodriguez, I.P.; Chakravarthi, B.V.S.K.; Varambally, S. UALCAN: A portal for facilitating tumor subgroup gene expression and survival analyses. Neoplasia 2017, 19, 649–658. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Aksoy, B.A.; Dogrusoz, U.; Dresdner, G.; Gross, B.; Sumer, S.O.; Sun, Y.; Jacobsen, A.; Sinha, R.; Larsson, E.; et al. Integrative Analysis of Complex Cancer Genomics and Clinical Profiles Using the cBioPortal. Sci. Signal. 2013, 6, pl1. [Google Scholar] [CrossRef]
- Cerami, E.; Gao, J.; Dogrusoz, U.; Gross, B.E.; Sumer, S.O.; Aksoy, B.A.; Jacobsen, A.; Byrne, C.J.; Heuer, M.L.; Larsson, E.; et al. The cBio Cancer Genomics Portal: An Open Platform for Exploring Multidimensional Cancer Genomics Data. Cancer Discov. 2012, 2, 401–404. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Fu, J.; Zeng, Z.; Cohen, D.; Li, J.; Chen, Q.; Li, B.; Liu, X.S. TIMER2.0 for analysis of tumor-infiltrating immune cells. Nucleic Acids Res. 2020, 48, W509–W514. [Google Scholar] [CrossRef]
- Patel, H.; Yacoub, N.; Mishra, R.; White, A.; Yuan, L.; Alanazi, S.; Garrett, J.T. Current Advances in the Treatment of BRAF-Mutant Melanoma. Cancers 2020, 12, 482. [Google Scholar] [CrossRef]
- Holčáková, J. Effect of DNA Methylation on the Development of Cancer. Klin. Onkol. Cas. Ceske Slov. Onkol. Spol. 2018, 31, 41–45. [Google Scholar] [CrossRef]
- Gil, M.; Kim, K.E. Systematic Multiomic Analysis of Ly75 Gene Expression and Its Prognostic Value through the Infiltration of Natural Killer (NK) Cells in Skin Cutaneous Melanoma. J. Clin. Med. 2020, 9, 1383. [Google Scholar] [CrossRef] [PubMed]
- Hong, J.T.; Son, D.J.; Lee, C.K.; Yoon, D.-Y.; Lee, D.H.; Park, M.H. Interleukin 32, inflammation and cancer. Pharmacol. Ther. 2017, 174, 127–137. [Google Scholar] [CrossRef] [PubMed]
- Heinhuis, B.; Koenders, M.; Berg, W.B.V.D.; Netea, M.G.; Dinarello, C.A.; Joosten, L.A.B. Interleukin 32 (IL-32) Contains a Typical α-Helix Bundle Structure That Resembles Focal Adhesion Targeting Region of Focal Adhesion Kinase-1. J. Biol. Chem. 2012, 287, 5733–5743. [Google Scholar] [CrossRef]
- Nicholl, M.B.; Chen, X.; Qin, C.; Bai, Q.; Zhu, Z.; Bs, M.R.D.; Fang, Y. IL-32α has differential effects on proliferation and apoptosis of human melanoma cell lines. J. Surg. Oncol. 2016, 113, 364–369. [Google Scholar] [CrossRef]
- Lee, J.; Kim, K.E.; Cheon, S.; Song, J.H.; Houh, Y.; Kim, T.S.; Gil, M.; Lee, K.J.; Kim, S.; Kim, D.; et al. Interleukin-32alpha induces migration of human melanoma cells through downregulation of E-cadherin. Oncotarget 2016, 7, 65825–65836. [Google Scholar] [CrossRef]
- Kang, Y.H.; Park, M.-Y.; Yoon, D.-Y.; Han, S.R.; Lee, C.I.; Ji, N.Y.; Myung, P.-K.; Lee, H.G.; Kim, J.W.; Yeom, Y.I.; et al. Dysregulation of overexpressed IL-32α in hepatocellular carcinoma suppresses cell growth and induces apoptosis through inactivation of NF-κB and Bcl-2. Cancer Lett. 2012, 318, 226–233. [Google Scholar] [CrossRef]
- Yun, H.-M.; Oh, J.H.; Shim, J.-H.; Ban, J.O.; Park, K.-R.; Kim, J.-H.; Lee, D.H.; Kang, J.-W.; Park, Y.H.; Yu, D.; et al. Antitumor activity of IL-32β through the activation of lymphocytes, and the inactivation of NF-κB and STAT3 signals. Cell Death Dis. 2013, 4, e640. [Google Scholar] [CrossRef]
- Jung, M.Y.; Son, M.H.; Kim, S.H.; Cho, D.; Kim, T.S. IL-32γ Induces the Maturation of Dendritic Cells with Th1- and Th17-Polarizing Ability through Enhanced IL-12 and IL-6 Production. J. Immunol. 2011, 186, 6848–6859. [Google Scholar] [CrossRef]
- Daltroy, L.H.; Larson, M.G.; Eaton, H.M.; Partridge, A.J.; Pless, I.B.; Rogers, M.P.; Liang, M.H.; Walker, L.S.; Van Slyke, D.A.; Newbrough, J.R. Psychosocial Adjustment in Juvenile Arthritis. J. Pediatr. Psychol. 1992, 17, 277–289. [Google Scholar] [CrossRef]
- Park, M.H.; Song, M.J.; Cho, M.-C.; Moon, D.C.; Yoon, D.Y.; Han, S.B.; Hong, J.T. Interleukin-32 enhances cytotoxic effect of natural killer cells to cancer cells via activation of death receptor 3. Immunology 2012, 135, 63–72. [Google Scholar] [CrossRef]
- Pagès, F.; Galon, J.; Dieu-Nosjean, M.-C.; Tartour, E.; Sautes-Fridman, C.; Fridman, W.H. Immune infiltration in human tumors: A prognostic factor that should not be ignored. Oncogene 2009, 29, 1093–1102. [Google Scholar] [CrossRef]
- Gil, M.; Kim, K.E. Interleukin-18 Is a Prognostic Biomarker Correlated with CD8+ T Cell and Natural Killer Cell Infiltration in Skin Cutaneous Melanoma. J. Clin. Med. 2019, 8, 1993. [Google Scholar] [CrossRef] [PubMed]
- Han, S.; Yang, Y. Interleukin-32: Frenemy in cancer? BMB Rep. 2019, 52, 165–174. [Google Scholar] [CrossRef]
- Rossi, G.R.; Trindade, E.S.; Souza-Fonseca-Guimaraes, F. Tumor Microenvironment-Associated Extracellular Matrix Components Regulate NK Cell Function. Front. Immunol. 2020, 11, 73. [Google Scholar] [CrossRef] [PubMed]
- Vitale, M.; Cantoni, C.; Pietra, G.; Mingari, M.C.; Moretta, L. Effect of tumor cells and tumor microenvironment on NK-cell function. Eur. J. Immunol. 2014, 44, 1582–1592. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Jiang, P.; Wei, S.; Xu, X.; Wang, J. Regulatory T cells in tumor microenvironment: New mechanisms, potential therapeutic strategies and future prospects. Mol. Cancer 2020, 19, 116. [Google Scholar] [CrossRef]
- Ren, J.; Yu, D.; Fu, R.; An, P.; Sun, R.; Wang, Z.; Guo, R.; Li, H.; Zhang, Y.; Li, Z.; et al. IL2RG -deficient minipigs generated via CRISPR/Cas9 technology support the growth of human melanoma-derived tumours. Cell Prolif. 2020, 53, e12863. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Markers | SKCM | LIHC | |||
|---|---|---|---|---|---|
| R | p | R | p | ||
| NK cell | KIR2DL1 | 0.33 | *** | −0.0046 | 0.93 |
| KIR2DL3 | 0.35 | *** | −0.055 | 0.29 | |
| KIR2DL4 | 0.6 | 0 | 0.056 | 0.28 | |
| KIR3DL1 | 0.37 | 0 | 0.018 | 0.72 | |
| KIR3DL2 | 0.58 | 0 | −0.024 | 0.64 | |
| KIR3DL3 | 0.14 | * | 0.029 | 0.57 | |
| KIR2DS4 | 0.21 | *** | −0.052 | 0.32 | |
| KLRK1 (NKG2D) | 0.69 | 0 | 0.065 | 0.21 | |
| NCR1 (NKp46) | 0.33 | *** | −0.039 | 0.46 | |
| NCR2 (NKp44) | 0.11 | 0.019 | −0.025 | 0.63 | |
| NCR3 (NKp30) | 0.47 | 0 | 0.09 | 0.085 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kang, J.Y.; Kim, K.E. Prognostic Value of Interleukin-32 Expression and Its Correlation with the Infiltration of Natural Killer Cells in Cutaneous Melanoma. J. Clin. Med. 2021, 10, 4691. https://doi.org/10.3390/jcm10204691
Kang JY, Kim KE. Prognostic Value of Interleukin-32 Expression and Its Correlation with the Infiltration of Natural Killer Cells in Cutaneous Melanoma. Journal of Clinical Medicine. 2021; 10(20):4691. https://doi.org/10.3390/jcm10204691
Chicago/Turabian StyleKang, Ji Young, and Kyung Eun Kim. 2021. "Prognostic Value of Interleukin-32 Expression and Its Correlation with the Infiltration of Natural Killer Cells in Cutaneous Melanoma" Journal of Clinical Medicine 10, no. 20: 4691. https://doi.org/10.3390/jcm10204691
APA StyleKang, J. Y., & Kim, K. E. (2021). Prognostic Value of Interleukin-32 Expression and Its Correlation with the Infiltration of Natural Killer Cells in Cutaneous Melanoma. Journal of Clinical Medicine, 10(20), 4691. https://doi.org/10.3390/jcm10204691