
Adenine Nucleotide Metabolites in Uremic Erythrocytes as Metabolic Markers of Chronic Kidney Disease in Children


Abstract
:1. Introduction
2. Materials
2.1. The Study Group
- Age of 3–18 years,
- Diagnosed CKD of varying degrees of progression
- And written consent to participate in the study.
- Group II—15 patients with stage II CKD; including 11 boys, 4 girls;
- Group III—16 patients with stage III CKD; including 10 boys, 6 girls;
- Group IV—8 patients with stage IV CKD; including 4 boys, 4 girls;
- Group of children undergoing renal replacement therapy (RRT)—9 patients undergoing.RRT (hemodialysis, peritoneal dialysis); including 7 boys, 2 girls.
2.2. Ethical Issues
3. Methods
3.1. Collection of Test Material Samples
3.2. Reagents
3.3. Preparation of Samples for the Determination of Nucleotide-Related Metabolites
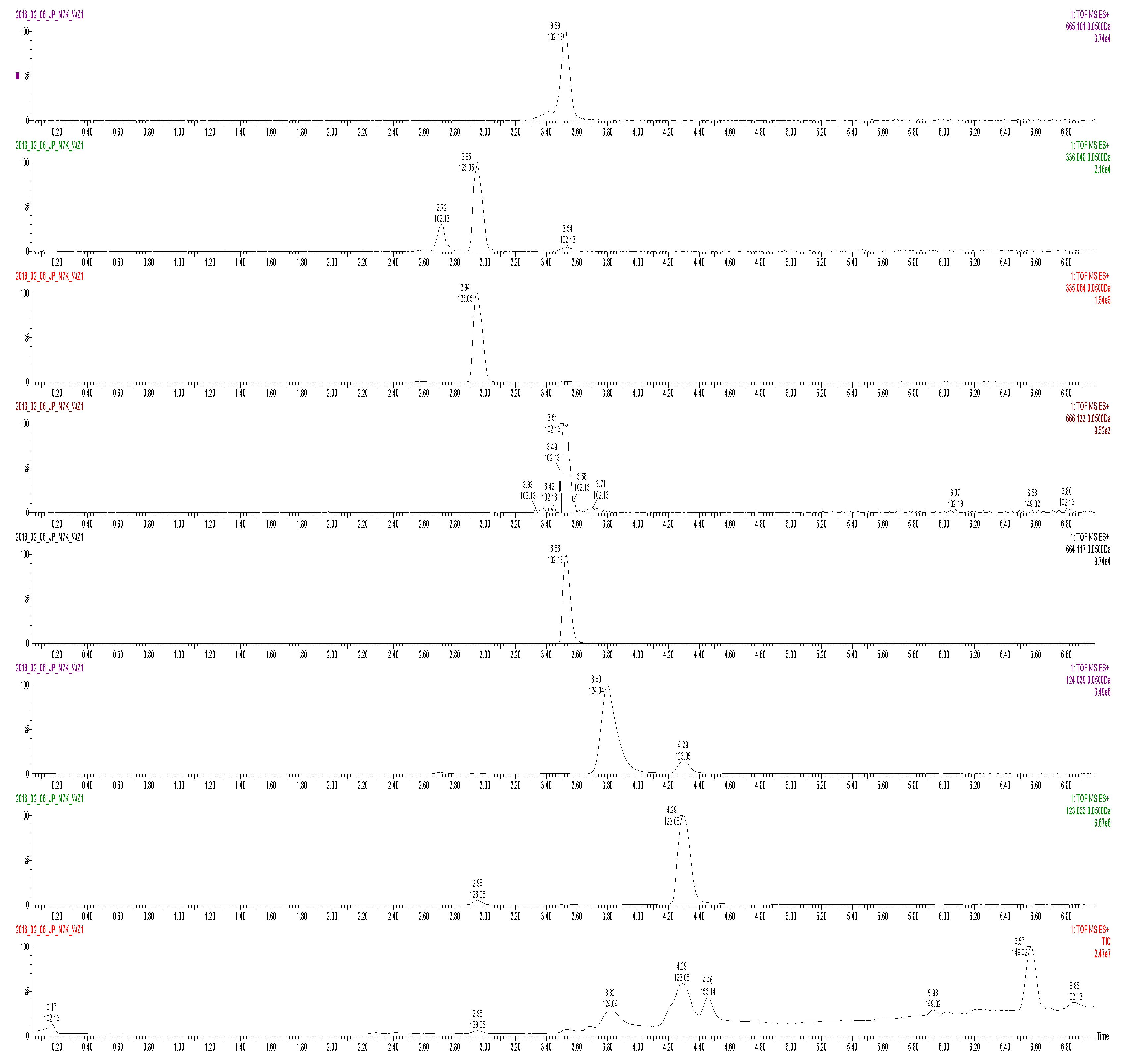
3.4. Method of Analysis for Nucleotide Metabolites
3.5. Statistical Analysis
4. Results
4.1. Comparison of the Results of Determinations between the CKD Children and Control Group
4.2. Comparison of the Content of Nucleotide-Related Metabolites with the CKD Severity
4.3. Assessment of Dependence in the Groups of Children with CKD
- - NAD and NAAD (r = 0.852, p = 0.001),
- and NAMN (r = 0.564, p = 0.001),
- and NMN (r = 0.641, p = 0.001),
- and with NADH (r = 0.850; p = 0.001).
- - NAAD and NAMN (r = 0.677, p = 0.001),
- and NMN (r = 0.742, p = 0.001)
- and with NADH (r = 0.765, p = 0.001).
- - NAMN and NMN (r = 0.874, p = 0.001)
- and with NADH (r = 0.542, p = 0.001)
- - NMN correlated positively with NADH (r = 0.585, p = 0.001)
5. Discussion
6. Conclusions
- CKD children do not have evident abnormalities of RBC metabolism with respect to adenine nucleotide metabolites.
- The significant differences in erythrocyte NAD concentrations between CKD stages may suggest the activation of adaptive defense mechanisms aimed at erythrocyte metabolic stabilization.
- It seems that the implementation of RRT has a positive impact on RBC NAD metabolism, but further research performed on a larger population is needed to confirm it.
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Chen, T.; Knicely, D.H.; Grams, M.E. Chronic Kidney Disease Diagnosis and Management. JAMA 2019, 322, 1294–1304. [Google Scholar] [CrossRef]
- Benito, S.; Sanchez-Ortega, A.; Unceta, N.; Andrade, F.; Aldámiz-Echevarria, L.; Goicolea, M.A.; Barrio, R.J. LC-QTOF-MS-based targeted metabolomics of arginine-creatine metabolic pathway-related compounds in plasma: Application to identify potential biomarkers in pediatric chronic kidney disease. Anal. Bioanal. Chem. 2016, 408, 747–760. [Google Scholar] [CrossRef]
- Benito, S.; Sánchez-Ortega, A.; Unceta, N.; Jansen, J.; Postma, G.; Andrade, F.; Aldámiz-Echevarria, L.; Kupdens, K.L.M.; Goicolea, M.A.; Barrio, R.J. Plasma biomarker discovery for early chronic kidney disease diagnosis based on chemometric approaches using LC-QTOF targeted metabolomics data. J. Pharm Biomed. Anal. 2018, 149, 46–56. [Google Scholar] [CrossRef] [Green Version]
- Hikosaka, K.; Ikutani, M.; Shito, M.; Kazuma, K.; Gulshan, M.; Nagai, J.; Takatsu, K.; Konno, K.; Tobe, K.; Kanno, H.; et al. Deficiency of nicotinamide mononucleotide adenylyltransferase 3 (nmnat3) causes hemolytic anemia by altering the glycolytic flow in mature erythrocytes. J. Biol. Chem. 2014, 289, 14796–14811. [Google Scholar] [CrossRef] [Green Version]
- Slee, A.D. Exploring metabolic dysfunction in chronic kidney disease. Nutr. Metab. 2012, 9, 36. [Google Scholar] [CrossRef] [Green Version]
- Cisek, K.; Krochmal, M.; Klein, J.; Mischak, H. The application of multi-omics and systems biology to identify therapeutic targets in chronic kidney disease. Nephrol. Dial. Transplant. 2016, 31, 2003–2011. [Google Scholar] [CrossRef] [Green Version]
- Mussap, M.; Antonucci, R.; Noto, A.; Fanos, V. The role of metabolomics in neonatal and pediatric laboratory medicine. Clin. Chim. Acta. 2013, 426, 127–138. [Google Scholar] [CrossRef]
- Imai, S.; Guarente, L. NAD+ and sirtuins in aging and disease. Trends Cell Biol. 2014, 25, 464–471. [Google Scholar] [CrossRef]
- Chiarugi, A.; Dölle, C.; Felici, R.; Ziegler, M. The NAD metabolome: A key determinant of cancer cell biology. Nat. Rev. Cancer. 2012, 12, 741–752. [Google Scholar] [CrossRef]
- Houtkooper, R.H.; Canto, C.; Wanders, R.J.; Auwerx, J. The secret life of NAD: An old metabolite controlling new metabolic signaling pathways. Endocr. Rev. 2010, 31, 194–223. [Google Scholar] [CrossRef] [Green Version]
- Picard, F.; Kurtev, M.; Chung, N.; Topark-Ngarm, A.; Senawong, T.; De Oliveira, R.M.; Leid, M.; McBurney, M.W.; Guarente, L. Sirt1 promotes fat mobilization in white adipocytes by repressing PPAR-gamma. Nature 2004, 6993, 771–776. [Google Scholar] [CrossRef]
- Rodgers, J.T.; Lerin, C.; Haas, W.; Gygi, P.; Spiegelman, B.M.; Puigserver, P. Nutrient control of glucose homeostasis through a complex of PGC-1alpha and SIRT1. Nature 2005, 7029, 113–118. [Google Scholar] [CrossRef]
- Liu, J.; Mohandas, N.; An, X. Membrane assembly during erythropoiesis. Curr. Opin. Hematol. 2011, 18, 133–138. [Google Scholar] [CrossRef]
- Liu, R.; Orgel, L.E. Enzymatic synthesis of polymers containing nicotinamide mononucleotide. Nucleic Acids Res. 1995, 23, 3742–3749. [Google Scholar] [CrossRef] [Green Version]
- Yamada, K.; Hara, N.; Shibata, T.; Osago, H.; Tsuchiya, M. The simultaneous measurement of nicotinamide adenine dinucleotide and related compounds by liquid chromatography/electrospray ionization tandem mass spectrometry. Anal. Biochem. 2006, 352, 282–285. [Google Scholar] [CrossRef]
- Schwartz, G.J.; Muñoz, A.; Schneider, M.F.; Mak, R.H.; Kaskel, F.; Furthet, S.L. New equations to estimate GFR in children with CKD. J. Am. Soc. Nephrol. 2009, 20, 629–637. [Google Scholar] [CrossRef] [Green Version]
- Graves, J.W. Diagnosis and management of chronic kidney disease. Mayo Clin. Proc. 2008, 83, 1064–1069. [Google Scholar] [CrossRef] [Green Version]
- Piechowicz, J.; Gajewska-Naryniecka, A.; Kukula, M.; Wiśniewski, J.; Gamian, A. Mass Spectrometry in Clinical Diagnostics. Med. Sci. Technol. 2017, 58, 98–110. [Google Scholar] [CrossRef]
- Cho, J.; Seo, J.; Lim, C. Mitochondrial ATP transporter Ant2 depletion impairs erythropoiesis and B lymphopoiesis. Cell Death Differ. 2015, 22, 1437–1450. [Google Scholar] [CrossRef] [Green Version]
- Rose, I.A. Regulation of human red cell glycolysis: A review. Exp. Eye Res. 1971, 11, 264–272. [Google Scholar] [CrossRef]
- Szołkiewicz, A. Purine Nucleotide Metabolism in Erythrocytes in Children with Cancer. Dissertation, Medical University of Gdańsk, Gdansk, Poland, 2013; pp. 50–63. [Google Scholar]
- Alexandrovich, Y.G.; Kosenko, E.A.; Sinauridze, E.I.; Obydennyi, S.I.; Kiree, I.I.; Ataullakhanov, F.I.; Kaminsky, Y.G. Rapid Elimination of Blood Alcohol Using Erythrocytes: Mathematical Modeling and In Vitro Study. BioMed Res. Int. 2017, 2017, 5849593. [Google Scholar] [CrossRef]
- Xie, N.; Zhang, L.; Gao, W.; Huang, C.; Huber, P.E.; Zhou, X.; Li, H.; Shen, G.; Zou, B. NAD+ metabolism: Pathophysiologic mechanisms and therapeutic potential. Signal Transduct. Target. Ther. 2020, 5, 227. [Google Scholar] [CrossRef]
- Reisz, J.A.; Tzounakas, V.L.; Nemkov, T. Metabolic Linkage and Correlations to Storage Capacity in Erythrocytes from Glucose 6-Phosphate Dehydrogenase-Deficient Donors. Front. Med. 2018, 4, 248. [Google Scholar] [CrossRef] [Green Version]
- Pericom, L.; Benigni, A. The iNADequacy of renal cell metabolism: Modulating NAD+ biosynthetic pathways to forestall kidney diseases. Kidney Int. 2019, 96, 264–267. [Google Scholar] [CrossRef]
- Polak-Jonkisz, D.; Purzyc, L.; Szczepańska, M.; Makulska, I. Erythrocyte caspase-3 levels in children with chronic kidney disease. Clin. Biochem. 2012, 46, 11–14. [Google Scholar] [CrossRef]
- Parikh, S.M. Metabolic Stress Resistance in AKI: Evidence for a PGC1α-NAD+ Pathway. Nephron 2019, 143, 184–187. [Google Scholar] [CrossRef]
- Ralto, K.M.; Rhee, E.P.; Parikh, S.M. NAD+ homeostasis in renal health and disease. Nat. Rev. Nephrol. 2020, 16, 99–111. [Google Scholar] [CrossRef]
- Mehr, A.P.; Tran, M.T.; Ralto, K.M.; Leaf, D.E.; Washco, V.; Messmer, J.; Lerner, A.; Kher, A.; Kim, S.H.; Khoury, C.C.; et al. De novo NAD+ biosynthetic impairment in acute kidney injury in humans. Nat. Med. 2018, 24, 1351–1359. [Google Scholar] [CrossRef]


{kind=link}
| Material | Studied Parameter | Study Group/Patient Group | Control Group | Significance Level p-Value | ||
|---|---|---|---|---|---|---|
| N | Mean ± SD | N | Mean ± SD | |||
| blood | GFR (mL/min/1.73 m2) | 48 | 45.96 ± 26.81 | 33 | 103.15 ± 10.88 | 0.0001 |
| creatinine (mg/dL) | 48 | 2.25 ± 1.99 | 33 | 0.56 ± 0.11 | 0.0001 | |
| inorganic phosphorus (mg/dL) | 44 | 4.94 ± 0.84 | 33 | 5.36 ± 1.17 | 0.194 (NS) | |
| calcium (mg/dL) | 43 | 9.80 ± 0.68 | 32 | 10.04 ± 0.47 | 0.020 | |
| sodium (mg/dL) | 48 | 139.46 ± 3.35 | 33 | 138.94 ± 2.73 | 0.381 (NS) | |
| potassium (mg/dL) | 48 | 4.47 ± 0.46 | 32 | 4.42 ± 0.39 | 0.643 (NS) | |
| urea (mg/dL) | 47 | 60.85 ± 38,91 | 32 | 26.16 ± 15.86 | 0.0001 | |
| Hb (g/dL) | 48 | 11.7 ± 1.51 | 34 | 14.4 ± 1.74 | 0.0001 | |
| Ht (%) | 48 | 34.33 ± 4.15 | 34 | 41.38 ± 4.88 | 0.0001 | |
| RBC (*106/µL) | 48 | 4.14 ± 0.71 | 34 | 4.85 ± 0.55 | 0.0001 | |
| Time [min] | Phase A [%] | Phase B [%] |
|---|---|---|
| 0 | 99 | 1 |
| 2.7 | 0 | 100 |
| 5.7 | 0 | 100 |
| 5.71 | 99 | 1 |
| 13 | 99 | 1 |
| Material | Studied Parameter | Study Group/Patient Group | Control Group | Significance Level p-Value | ||||
|---|---|---|---|---|---|---|---|---|
| N | Mean ± SD | Median (Q25 Q75) | N | Mean ± SD | Median (Q25 Q75) | |||
| erythrocytes | NAD | 48 | 216.98 ± 117.87 N | 222.48 (119.84–317.8) | 33 | 233.30 ± 113.11 | 256.08 (177,60–289.12) | 0.269 (NS) |
| NA | 47 | 8.69 ± 5.08 N | 7.84 (5.44–10.24) | 33 | 9.04 ± 4.66 | 8.00 (6.40–11.52) | 0.756 (NS) | |
| NAM | 46 | 298.56 ± 238.78 | 171.04 (132.16–510.72) | 33 | 242.39 ± 204.04 | 150.64 (139.6–183.52) | 0.183 (NS) | |
| NAAD | 47 | 119.29 ± 73.45 N | 121.92 (49.04–188.00) | 33 | 136.40 ± 69.60 | 156.00 (86.40–172.24) | 0.350 (NS) | |
| NAMN | 47 | 40.00 ± 8.61 | 38.56 (33.28–44.40) | 33 | 41.28 ± 10.30 | 38.00 (33.76–46.48) | 0.809 (NS) | |
| NMN | 47 | 40.90 ± 9.75 | 40.08 (33.52–48.16) | 33 | 43.53 ± 10.68 | 41.44 (34.80–49.84) | 0.350 (NS) | |
| NADH | 47 | 92.38 ± 53.66 N | 106.64 (32.64–129.44) | 33 | 105.61 ± 59.30 | 101.84 (62.72–146.72) | 0.273 (NS) | |
| Variable | NAD | Multiple Comparisons p Values (with a Bonferroni Adjustment) | |||||
|---|---|---|---|---|---|---|---|
| Stages of CKD | N | Median (Q25 Q75) | Control | II | III | IV | RRT |
| Control | 33 | 256.08 (177.6–289.12) | 1.000 | 1.000 | 0.148 | 1.000 | |
| II | 15 | 201.6 (78.64–303.28) | 1.000 | 1.000 | 0.032 | 1.000 | |
| III | 16 | 221.12 (144.84–261.84) | 1.000 | 1.000 | 0.046 | 1.000 | |
| IV | 8 | 340.52 (315.88–353.28) | 0.148 | 0.032 | 0.046 | 0.194 | |
| RRT | 9 | 252.00 (23.44–304.88) | 1.000 | 1.000 | 1.000 | 0.194 | |
| Variables | NA | NAM | NAAD | NAMN | NMN | NADH | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Stages of CKD | N | Median (Q25 Q75) | N | Median (Q25 Q75) | N | Median (Q25 Q75) | N | Median (Q25 Q75) | N | Median (Q25 Q75) | N | Median (Q25 Q75) |
| Control | 33 | 8.00 (6.40–11.52) | 33 | 150,64 (139.6–183.52) | 33 | 156.00 (86.40–172.24) | 33 | 38.00 (33.76–46.48) | 33 | 41.44 (34.8–49.84) | 33 | 101.84 (62.72–146.72) |
| II | 14 | 8.08 (5.44–9.76) | 14 | 202.52 (147.52–566.88) | 14 | 117.60 (44.08–178.16) | 14 | 38.88 (34.56–45.92) | 14 | 40.44 (34.32–49.36) | 14 | 97.36 (27.20–121.52) |
| III | 16 | 7.72 (5.40–10.44) | 15 | 136.56 (128.88–510.72) | 16 | 120.64 (61.72–152.76) | 16 | 37.28 (33.24–42.84) | 16 | 40.24 (33.56–47.52) | 16 | 74.56 (39.16–110.92) |
| IV | 8 | 8.76 (6.44–15.60) | 8 | 177.48 (151.64–362.32) | 8 | 192.88 (66.40–212.00) | 8 | 38.64 (31.64–45.16) | 8 | 40.52 (32.00–50.80) | 8 | 125.88 (116.56–181.04) |
| RRT | 9 | 6.32 (3.36–8.72) | 9 | 159.52 (138.64–184.4) | 9 | 141.76 (16.24–188.00) | 9 | 42.24 (33.92–47.84) | 9 | 38.56 (31.28–48.08) | 9 | 97.92 (26.00–126.56) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Piechowicz, J.; Gamian, A.; Zwolińska, D.; Polak-Jonkisz, D. Adenine Nucleotide Metabolites in Uremic Erythrocytes as Metabolic Markers of Chronic Kidney Disease in Children. J. Clin. Med. 2021, 10, 5208. https://doi.org/10.3390/jcm10215208
Piechowicz J, Gamian A, Zwolińska D, Polak-Jonkisz D. Adenine Nucleotide Metabolites in Uremic Erythrocytes as Metabolic Markers of Chronic Kidney Disease in Children. Journal of Clinical Medicine. 2021; 10(21):5208. https://doi.org/10.3390/jcm10215208
Chicago/Turabian StylePiechowicz, Joanna, Andrzej Gamian, Danuta Zwolińska, and Dorota Polak-Jonkisz. 2021. "Adenine Nucleotide Metabolites in Uremic Erythrocytes as Metabolic Markers of Chronic Kidney Disease in Children" Journal of Clinical Medicine 10, no. 21: 5208. https://doi.org/10.3390/jcm10215208
APA StylePiechowicz, J., Gamian, A., Zwolińska, D., & Polak-Jonkisz, D. (2021). Adenine Nucleotide Metabolites in Uremic Erythrocytes as Metabolic Markers of Chronic Kidney Disease in Children. Journal of Clinical Medicine, 10(21), 5208. https://doi.org/10.3390/jcm10215208